Darwin's geological research in Argentina Aguirre-Urreta, Beatriz; Griffin, Miguel; Ramos, Victor A. Charles Robert Darwin and Argentina's National Academy of Sciences Depetris, Pedro José
Darwin's observation in South America: what did he find at agua de la zorra, Mendoza province? Poma, Stella; Litvak, Vanesa D.; Koukharsky, Magdalena; Maisonnave, E. Beatriz; Quenardelle, Sonia
Darwin forest at agua de la zorra: the first in situ forest discovered in South America by Darwin in 1835 Brea, Mariana; Artabe, Analía E.; Spalletti, Luis A.
On Darwin's footsteps across the Andes: Tithonian-Neocomian fossil invertebrates from the Piuquenes pass Aguirre-Urreta, Beatriz; Vennari, Verónica
The High Andean Cordillera of central Argentina and Chile along the Piuquenes Pass-Cordon del Portillo transect: Darwin's pioneering observations compared with modern geology Giambiagi, Laura; Tunik, Maisa; Ramos, Victor A.; Godoy, Estanislao
The first geological map of Patagonia Zappettini, Eduardo O.; Mendía, José
The stratigraphy of cretaceous mudstones in the eastern Fuegian Andes: new data from body and trace fossils Olivero, Eduardo B.; Medina, Francisco A.; López C., María I.
Darwin's great patagonian tertiary formation at the mouth of the río Santa Cruz: a reappraisal Parras, Ana; Griffin, Miguel
Sedimentology and paleontology of a miocene marine succession first noticed by Darwin at Puerto Deseado (Port Desire) Casadío, Silvio; Griffin, Miguel
Charles Darwin and the first scientific observations on the patagonian shingle formation (Rodados Patagónicos) Martínez, Oscar A.; Rabassa, Jorge; Coronato, Andrea
Charles Darwin and the oldest glacial events in Patagonia: the erratic blocks of the Río Santa Cruz valley Strelin, Jorge; Malagnino, Eduardo
From Buenos Aires to Santa Fe: Darwin's observations and modern knowledge Iriondo, Martin; Kröhling, Daniela
On the formations of the Pampas in the footsteps of Darwin: south of the Salado Zárate, Marcelo; Folguera, Alicia
Geology of the area of Bahía Blanca, Darwin's view and the present knowledge: a story of 10 million years Quattrocchio, Mirta E.; Deschamps, Cecilia M.; Zavala, Carlos A.; Grill, Silvia C.; Borrome, Ana M.
The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle Fernicola, Juan Carlos; Vizcaíno, Sergio F.; De Iuliis, Gerardo
Young Darwin and the ecology and extinction of pleistocene south american fossil mammals Vizcaíno, Sergio F.; Fariña, Richard A.; Fernicola, Juan Carlos
Darwin at Puente del Inca: observations on the formation of the Inca's bridge and mountain building Ramos, Victor A.
Darwin's geological research in Argentina
Beatriz Aguirre-Urreta, Miguel Griffin, Victor A. Ramos Invited Editors

Portrait with signature gifted by Charles Darwin to the Academia Nacional de Ciencias in 1878 acknowledging his nomination as an Honorary Member. Photograph taken by Julia Margaret Cameron in 1868. Unpublished original copy kept in the Academia (see Depetris this volume)
On the occasion of the 200th. anniversary of Charles Darwin's birth, the Asociación Geológica Argentina decided to prepare a special issue devoted to the geological research undertaken by Darwin in Argentina. As it is well known, during his journeys on board HMS Beagle under the command of Captain Robert FitzRoy, he had the opportunity to survey overland different areas of South America. Darwin spent nearly three years - between August 1832 and April 1835 - visiting and studying different regions of our country, as illustrated in figure 1.

Figure 1: Journeys of HMS Beagle in southern South America (base map modified from Bowly 1990). Dates of Darwin's rides overland are shown in bold while the trips on board HMS Beagle are shown in italics, taken from Keynes (2001).
The aim of this special issue is to analyze his important geological observations and to emphasize the validity of many of his ideas under a 21st Century perspective. In order to accomplish this aim, several key localities that Darwin examined from a geological point of view during his voyage were selected. Such an analysis was carried out by several geologists and paleontologists well acquainted with the diverse problems that Darwin faced along his journeys in Argentina. HMS Beagle's main mission was to survey the coasts of southern South America, which it carried out during several journeys along the coast. The time needed to chart the coast in details gave Darwin several opportunities to spend considerable time on land. During the survey he also had many occasions to land at selected points along the Atlantic coast. The first of these fieldtrips was done between August and October 1832 and gave him the opportunity to visit Punta Alta near the fort of Bahía Blanca (Fig. 1). The second one was between November 1832 and May 1833 to visit the Beagle Channel, the surrounding areas of Tierra del Fuego Island, and the Malvinas Islands. The third and more important fieldtrip was, after landing in Carmen de Patagones in August 1833 (Fig. 1), to ride across the Pampas to Buenos Aires city and later on to Santa Fé and Bajada (today Paraná city) on the Río Paraná. The fourth trip began in Montevideo in December 1833 and allowed Darwin to examine various points along the Patagonian coast such as Puerto Deseado and Puerto San Julián. Afterwards HMS Beagle sailed to Tierra del Fuego where it spent nearly four months until March 1834. Later on it visited again different areas of the Malvinas Islands. In April of 1834 it anchored in the mouth of Río Santa Cruz. It was during this trip that Darwin, FitzRoy and more than 20 crew members sailed upstream the Río Santa Cruz in three small whale boats, almost reaching its headwaters near the foothills of the Patagonian Andes. This fourth trip continued across the Magallanes and strait of along the Pacific coast landing in Valparaíso in July 1834, and spending several months surveying the Chilean coast. Darwin's last visit to Argentina was between March and April 1835 (Fig. 1), when he crossed the Andes between Santiago and Mendoza following two different cordilleran passes. The different contributions of this issue were organized based on stratigraphic record and location (Fig. 2). The sections of the Andes are the first to be analyzed, then Patagonia and Tierra del Fuego, and finally the Cenozoic of the Pampas, ending with the origin of Puente del Inca and Darwin's tectonic ideas on the formation of the Andes.

Figure 2: Approximate location of the different contributions dealing with Darwin's geological research in Argentina indicated by an asterisk (compare with Figure 1).
Pedro J. Depetris describes the documents kept in the Academia Nacional de Ciencias in Córdoba related to the nomination of Darwin, first as corresponding member and then as honorary member of the Academy. This paper also presents Darwin's original letter where he acknowledges the nomination and the front page of the book on the Origin of the Species that Darwin dedicated to the academy. Stella Poma, Vanesa D. Litvak, Magdalena Koukharsky, Beatriz Maisonnave and Sonia Quenardelle describe the rocks in which the Triassic forest at Agua de la Zorra is included as being part of a pyroclastic flow. Their petrographic studies confirm the previous description of Darwin on the presence of fine volcanic material. Based on their observations, they answer Darwin's queries about the process that buried the forest in standing position. It should be noted that these pyroclastic flows were unknown at the time of Darwin's visit. Mariana Brea, Analía E. Artabe and Luis A. Spalletti deal with the first in situ forest discovered in South America by Darwin in his trip across the Andes. This forest was described in detail by Darwin as monotypic and assigned to the Tertiary. Their study reveals that the paleovegetation of Agua de La Zorra forest is composed by conifers and corystosperms reaching up to 20-26 meters high forming a two-storeyed arboreal strata and an understorey of ferns. It was reassigned to the Triassic and reconstructions of the forest landscape are presented. Beatriz Aguirre-Urreta and Verónica Vennari study a small collection of fossil invertebrates from the Piuquenes Pass, a locality discovered by Darwin during his crossing of the High Andes from Santiago to Mendoza. They compared this collection with the fossils that allowed Darwin to propose a Neocomian age for the bearing strata. This assignation is confirmed and the time span stretched to the Tithonian based on key ammonites of that age. Laura Giambiagi, Maisa Tunik, Victor A. Ramos and Estanislao Godoy compare the observations of Darwin on the structure of the High Andes between the Piuquenes and Portillo passes with modern structural knowledge. They remark the awareness of Darwin on the episodic uplift of the Andes with the rise of the Principal Cordillera first and a later uplift of the Frontal Cordillera, both in post- Cretaceous times. Eduardo Zappettini and José Mendía introduce the first geological map of Patagonia ever compiled, publishing the first color hand-drafted copy. It was drawn by Darwin after his trip to South America. These authors outline the main geological units used in the map and validate them with the present knowledge of the geology of the region. Eduardo B. Olivero, Francisco A. Medina and María I. López C. analyze the still unsolved problem of Darwin's "Clay Slate Formation" in the Fueguian Andes. This term coined for the Cretaceous successions, presently includes a series of marine mudstones the stratigraphy of which is based on scarce fossils, mainly inoceramids and ammonites. The present use of trace fossils and distinctive ichnofabrics, together with new ammonites, are contributing to a better understanding of these mudstones, improving stratigraphic correlation among different units. Ana Parras and Miguel Griffin present the sedimentology and fossil contents of a classic Miocene locality, first described by Darwin at the southern bank of Río Santa Cruz, along the Patagonian coast. This is the type locality of most of the fossils found in his "Great Patagonian Tertiary Formation" and later described by Sowerby (1846). The rocks of this section are presently included in the Monte León Formation. The contribution by Silvio Casadio and Miguel Griffin deals with the sedimentology and paleontology of a classic locality at Puerto Deseado (Port Desire). This locality was visited by Darwin in 1833 when he collected fossil mollusks, and remained nearly unexplored by geologists and paleontologists ever since. The authors assess this classic Miocene locality and correlate it with several similar sequences exposed along the Atlantic cliffs. Oscar A. Martínez, Jorge Rabassa and Andrea Coronato discuss one of the most intriguing lithostratigraphic units in the Late Cenozoic of Patagonia named by Darwin the "Patagonian Shingle Formation". They analyze the prevailing paradigm of that time that assigned these rocks to the "Great Universal Dèlluge" and give a modern perspective to the origin of the "Rodados Patagónicos". Jorge Strelin and Eduardo Malagnino analyze the observations of Darwin in his trip upstream the Río Santa Cruz in 1834. Special emphasis is paid to the extension of the big erratic boulders and gravels described by Darwin which are now interpreted as indicators of a maximum glacial expansion in Patagonia. Those blocks together with some new findings are ascribed by these authors to a catastrophic event related to the outburst of a large lake. Martín Iriondo and Daniela Kröhling cover the visit of Darwin to Santa Fé with special emphasis in those localities described by him along the Río Paraná banks. The authors stress the precise descriptions of Darwin and discuss the Late Cenozoic age of these sediments. Marcelo Zárate and Alicia Folguera deal with the geologic characteristics of the Pampas south of Río Salado, a region that impressed Darwin on his ride from Bahía Blanca to Buenos Aires. They discuss the Pampean Formation of Darwin, its present stratigraphic divisions, its age, and the different environments encompassed in this unit, from a modern perspective. Mirta E. Quattrocchio, Cecilia M. Deschamps, Carlos A. Zavala, Silvia C. Grill and Ana M. Borromei give an updated vision of the scenery described by Darwin when he visited Bahía Blanca and surrounding areas. Multidisciplinary results were integrated by these authors in a chronosequence chart. This diagram shows the suggested space-time correlation of the recognized events in the Late Miocene-Holocene history of the southwestern Pampas. Juan Carlos Fernicola, Sergio F. Vizcaíno and Gerardo De Iuliis provide a synthesis of the present knowledge of the large fossil mammals collected by Darwin, mostly from the Pampean region of central Argentina. These authors ponder on the reasons that led Darwin to recognize only megatheres and mastodonts for the large fossil remains and discuss the current taxonomic status of the taxa described or erected by Owen between 1837 and 1845. Sergio F. Vizcaíno, Richard A. Fariña and Juan Carlos Fernicola, in their contribution on the Pleistocene South American mammals, attempt to provide answers to Darwin's questions, such as what has exterminated so many living creatures? Their study on the palaeobiology of these organisms shed light on the understanding of their evolution intermingled with ecology and phylogeny. Victor A. Ramos searches on the origin of Puente del Inca, a natural bridge described by Darwin in the High Andes, during his cross by the Uspallata o La Cumbre Pass. Applying uniformitarism, the author interprets the origin of Puente del Inca as initially formed as an ice bridge, latter on cemented by hot spring waters. Ramos also weighs Darwin's thoughts on mountain uplift based on the Mesozoic successions of the Puente del Inca section. His old ideas on tectonics anticipate in many decades some concepts of the geosynclinal theory and also on the present fold and thrust belt mechanics. We would wish to acknowledge the reviewers of the present issue: S. Bargo (Universidad Nacional de La Plata), M. Bond (Universidad Nacional de La Plata), N. Carmona (CADIC, Ushuaia), C. Costa (Universidad Nacional de San Luis), R. Charrier (Universidad de Chile), J.S. Crampton (Institute of Geological and Nuclear Sciences, New Zealand), A. Crame (British Antarctic Survey, UK), D. Croft (Case Western Reserve University, USA), I.W.D. Dalziel (University of Texas, USA), R. De Barrio (INREMI, La Plata), E. Derbyshire (University of London, UK), C. Deschamps (Universidad Nacional de La Plata), E. Godoy (Santiago de Chile), P. Imbellone (Universidad Nacional de La Plata), C. Janis (Brown University, USA), M. Kaplan (Columbia University, USA), S.M. Kay (Cornell University, USA), H.A. Leanza (MACN, Buenos Aires), E. Llambías (CIG, La Plata), T. Manera (Universidad Nacional del Sur, Bahía Blanca), S. Martinez (Universidad de la República, Uruguay), G. McDonald (National Park Service, USA), A. Mourgues (SERNAGEOMIN, Chile), C.A. Mpodozis (Antofagasta Minerals, Chile), D. Muhs (United States Geological Survey, USA), S. Nielsen (Christian-Albrechts-Universität, Germany), O. Orfeo (Centro de Ecología Aplicada del Litoral, Corrientes), E.G. Ottone (Universidad de Buenos Aires), J. Panza (SEGEMAR, Buenos Aires), R.N. Page (SEGEMAR, Buenos Aires), G. Pastorino (MACN, Buenos Aires), G.J. Retallack (University of Oregon, USA), P. Sruoga (SEGEMAR, Buenos Aires), J. Stilwell (Monash University, Australia), M. Suarez (SERNAGEOMIN, Chile), M. Toledo (Muséum National d'Histoire Naturelle, France) and G. Vucetich (Universidad Nacional de La Plata).
WORKS CITED IN THE TEXT
1. Bowly, J. 1990. Charles Darwin, a new biography. Hutchinson Press, 511 p., London, Sydney, Auckland and Johannesburg.
2. Keynes, R.D. 2001. Charles Darwin's Beagle Diary. Cambridge Univesity Press. 464 p., Cambridge.
3. Sowerby, G.B. II 1846. Descriptions of the Tertiary fossil shells from South America. In Darwin, C. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. Ed., Appendix: 249-264, London.
Charles Robert Darwin and Argentina's National Academy of Sciences
Pedro José Depetris
Academia Nacional de Ciencias, Avenida Vélez Sarsfield 229, 5000 Córdoba.
ABSTRACT: Over 175 years ago Charles Robert Darwin arrived in Argentina to find a bare and boundless plain, the brave centaur called "gaucho", Quaternary fossils everywhere, and a society strikingly strange and aggressive to the British eyes of the young traveller. Although the voyage aboard HMS Beagle was the indispensable way towards increasing his stature as a biologist, Lyell's work awakened an inquisitive geological mind which allowed him to wonder at the splendour of the Andes. Forty-two years after having concluded his voyage on the Beagle, the National Academy of Sciences of Argentina appointed him as an Honorary Member. This must be interpreted as an early gesture of recognition -in the context of those times- to the magnificence of his scientific work.
Keywords: Darwin; Argentina; National Academy of Sciences; Geology; Sarmiento.
RESUMEN: Charles Robert Darwin y la Academia Nacional de Ciencias. Hace más de 175 años, Charles Robert Darwin llegaba a la Argentina para descubrir una llanura desprovista de límites y de árboles, el valeroso centauro que era el gaucho, fósiles cuaternarios por doquier y una sociedad sorprendentemente extraña y agresiva a los británicos ojos del joven viajero. Aunque el viaje a bordo del Beagle fue el camino indispensable para incrementar su estatura como biólogo, la obra de Lyell despertó una mente inquisitivamente geológica que le permitió maravillarse ante la magnificencia de los Andes. Cuarenta y dos años después de haber concluido su viaje en el Beagle, la Academia Nacional de Ciencias de Argentina lo designó Miembro Honorario, en lo que debe interpretarse como un temprano gesto de reconocimiento -en el contexto del momento- por la magnificencia de su obra científica.
Palabras clave: Darwin; Argentina; Academia Nacional de Ciencias; Geología; Sarmiento.
Recibido: 25 de agosto de 2008 Aceptado: 2 de octubre de 2008
INTRODUCTORY COMMENTS
On July 26, 1832, one day short of seven months after leaving Plymouth harbor, HMS. Beagle moored at Montevideo. It was a Cherokee class 10-gun brig-sloop of the Royal Navy named after the Beagle, the famous British dog breed. The most important task entrusted by the British Admiralty to 26-year-old Captain Robert FitzRoy was about to begin - the survey of the coasts south of the Río de la Plata, down to the southern tip of Patagonia and Tierra del Fuego. There were significant gaps in the geographical knowledge that the British Admiralty had gathered through countless reports of sailors who had visited the region for many years. Now that George Canning had signed in 1825 a commercial treaty with the newly independent federation of Argentinean states, trade was flourishing with the ex Spanish colonies and accurate geographical information was urgently needed. Charles Robert Darwin was only 22- years-old when he saw the muddy waters of the Río de la Plata for the first time; he had been recommended to FitzRoy as a companion and naturalist without pay (Darwin's father covered all the expenses involved in the long voyage) by John Stevens Henslow, clergyman, botanist, and mineralogist, whom Darwin had met in Cambridge. Young Darwin was expected to gather information on the natural history and geology of many exotic regions that he was going to visit during a two-year long journey around the world. The trip was initially extended to threeyears and then to five-years and it would be the most extraordinary experience in the life of Charles Darwin; an experience that would change dramatically the view that humankind had of itself and of nature. In his memoirs, he would simply state that "the voyage of the Beagle has been by far the most important event in my life and has determined my whole career". Darwin had left unfinished his medical education at Edinburgh in 1827, and had gone to Cambridge's Christ's College where he obtained his Bachelor of Arts (BA) degree, without honors. He was an enthusiastic entomologist and an avid biological collector and his experience in the Earth Sciences was limited to a field trip to northern Wales with the famous Adam Sedgwick (1785-1873), Woodwardian Professor of Geology at Cambridge. Notwithstanding, Darwin was supposed to become a clergyman and his father -Robert Waring Darwin- initially had strong objections when Charles was offered the opportunity of boarding the HMS Beagle for a journey to South America to "survey the S. extremity". However, after Charles's uncle, Josiah Wedgwood II, wrote an eloquent letter in which he stated that being young Charles a man of enlarged curiosity, "...it affords him such an opportunity of seeing men and things as happens to few". After an absence of five years and two days, on October 2nd, 1836, the Beagle anchored at Falmouth, and on the 4th Darwin returned home to Shrewsbury. As we know today, the voyage of the Beagle made him a scientific celebrity, as he produced several books and numerous articles, and profusely described and distributed many specimens that he brought home with him aboard the ship (Fig. 1). All these events rapidly led him to join the elite world of international science on an equal footing. The revolutionary ideas set into motion by this long voyage swirled through Darwin's later life until, precipitated by Alfred Russel Wallace, they suddenly broke through into Victorian society in 1859, under the form of a book that bore the provocative title of On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life (Fig. 2).
Figure 1: Portrait of young (~31-year-old) Charles Darwin four years after he returned from the voyage of the HMS. Beagle, painted by George Richmond (1840).

Figure 2: Facsimile of the front page of Origin of Species. The volume was sent by the publisher to the National Academy of Sciences, in Córdoba, upon Darwin's request.
The best place to find a detailed and personal account of Darwin's rich experiences is his first book, entitled Journal of Researches into the Geology and Natural History of the Various Countries visited by H.M.S. Beagle, under the Command of Captain FitzRoy, R.N. from 1832 to 1836 (Fig. 3). The book was a success when first published, in 1839, and Darwin's fondness for this particular work was transparent when he wrote at the end of his life that "the success of this my first literary child always tickles my vanity more than that of any of my other books". Originally published in four volumes (Darwin's volume, plus FitzRoy's two and an extended appendix), Darwin describes not only the exciting travels, the exotic ports of call, and the fascinating inland expeditions (in fact, he spent much more time on land than he did at sea), but also his emotions, his intense feelings on first arriving in the tropics, his dangerous overland excursions with the gauchos (which he deeply admired), and the awesome sight of the stars over the Cordillera de los Andes.

Figure 3: Facsimile of the front page of Darwin's Journal of Researches, a.k.a. Voyage of the Beagle.
Moreover, in his first work - later published as Voyage of the Beagle (e.g., Darwin 1989)- it is very clear that Darwin, at least, was never a detached observer and the book shows how he saw a totally foreign society through British eyes and strongly supported the social order and political structure as he understood it from his native country. Along this line, it is particularly interesting to read about his impressions and views on Argentine society in times of severe turmoil, when Juan Manuel de Rosas (whom he met personally) was growing as a political figure in the conflictive scenario that was at the time the Argentine Confederation. Darwin set foot for the first time on Argentina's soil in Patagonia when the Beagle arrived on August the 3rd off the mouth of the Río Negro. He traveled afterwards overland to Bahía Blanca, Buenos Aires, and Santa Fe. On December 6th 1833 the Beagle set sail to Patagonia (Puerto Deseado) and then anchored within the mouth of the Río Santa Cruz, which was explored upstream for several weeks. On June 10th, 1834, the Beagle "bade farewell for ever to Tierra del Fuego". Darwin was not particularly fond of sailing rough seas and one can only imagine the feeling of relief that he experienced when they left Cape Horn for good. Darwin would enter again Argentina's territory when he crossed the Andes and arrived in Mendoza during the 2nd half of March, 1835. The awe-inspiring sight of the Cordillera and the possibility of seeing geological processes at such large scale made a very deep impression on the young scientist.
DARWIN, THE GEOLOGIST
Since this article is published in a geological journal, it seems appropriate to briefly consider the geological facet of Charles Darwin. It is accepted now that geology influenced Darwin and, conversely, he influenced the science (Herbert 2005). Like FitzRoy, he had a special interest in geology and he had read most of the best-known texts in the field, particularly Charles Lyell's famous Principles of Geology (Lyell 1998), which was originally published in 1830. This shared interest in geological sciences was probably a factor that drew FitzRoy and Darwin together during their time on board. In fact, FitzRoy was approached by Lyell before the Beagle sailed off Plymouth in order to ask that specific geological features be recorded, such as erratic boulders of glacial origin. Darwin gathered geological specimens and took detailed notes on geology during the circumnavigation of the globe. Contradicting widespread belief, upon his return to Great Britain, it was his geological findings that first promoted enthusiastic scientific and public opinion. It must be kept in mind that, as a young scientist, Charles Darwin was eager to contribute with a simple theory that would explain most, if not all, of the observed geological phenomena. Only his scheme explaining the structure and distribution of coral reefs has survived to this day (Herbert 2005). The British government sponsored publication of his research and numerous geologists, including Darwin's former teachers like Sedgwick, proved to be an interested audience. Doubtlessly the experience of the voyage of the Beagle had been transformative: the methods and hypothesis of Victorian-era geology profoundly shaped Darwin's mind and his scientific methods as he worked toward a complete comprehension of evolution and natural selection.
DARWIN AND ARGENTINA'S NATIONAL ACADEMY OF SCIENCES
The Academy was founded on September 11th 1869, by a bill passed on to Congress by Argentina's President Domingo Faustino Sarmiento (García Castellanos 1987). The bill authorized President Sarmiento to hire a significant number of European scientists in order to promote scientific research in the natural and exact sciences and foster higher education at the Universidad Nacional de Córdoba. The first to arrive in August 1871 was Prof. M. Siewert, from the Universität Halle (known today as Martin-Luther-Universität Halle-Wittenberg), soon followed by Prof. P.G. Lorentz, a botanist from the Universität Munchen. Others, like the geologist Stelzner, the zoologist H. Weyenbergh, the astronomer K. Schultz-Sellack, arrived the following year (García Castellanos 1987). As academicians and naturalists from prominent European centers of higher learning, all undoubtedly knew about the revolutionary ideas put forward in the Origin of Species and, although not all supported Darwin's views (G. Burmeister, a prominent biologist, organizer of the Academy, for example, opposed evolutionary ideas), there is ground to sustain that most were enthusiastic supporters of Darwin's work. Moreover, during his visit to Argentine territory Charles Darwin made numerous and valuable observations that increased the knowledge on the natural history and geology of the young country. Hence, it should not come as a surprise that Darwin's magnificent work and his contact with Argentina led to his appointment as a Corresponding Member ("Miembro Corresponsal") of the National Academy of Sciences, which still has its seat in the city of Córdoba, Argentina. On August 7th 1878, the President of the Academy, Dr. Hendrik Weyenbergh, wrote a letter to Argentina's Minister of Education Dr. Bonifacio Lastra requesting the appointment of several new members and the change of Charles Robert Darwin's status to Honorary Member ("Miembro Honorario") (Fig. 4), along with D. A. Grisebach. In return, Darwin instructed his publisher to send a copy of the Origin of Species to the Academy (still in the Academy's collection of antique books), sent an autographed picture (see frontispiece of this volume), and wrote a grateful letter to the Academy's President. Darwin's letter dated March 18th 1879 to the Academy's President, Dr. Hendrik Weyenbergh, read: "Dear Sir, I beg leave to acknowledge the receipt of the very handsome Diploma of your Society, and to repeat my thanks for the honour conferred on me. According to your request I enclose my photograph, and I have directed my publisher to send a copy of my Origin of Species to the Society as I suppose that this is the best of my works. I have the honour to remain, Dear Sir, Yours faithfully, Charles Darwin" (Fig. 5).

Figure 4: Facsimile of the letter sent by the National Academy of Science's President, Dr. Hendrik Weyenbergh to Argentina's Minister of Education, Dr. Bonifacio Lastra, requesting the appointment of Charles Darwin as an Honorary Member of the Academy.

Figure 5: Facsimile of the letter sent by Charles Darwin on March 18th 1879 to the Academy's President, Dr. Hendrik Weyenbergh, acknowledging his appointment as Honorary Member of Argentina's National Academy of Sciences.
To this day, the volume presented by Darwin as a gift, as well as the photograph and the letter, are cherished in Argentina's Academy of Sciences as valuable icons of the development of world science. Darwin's approach to explain the evolution of nature and, above all, of humankind rapidly rooted in Argentina's newly born but growing science. Perhaps nothing better than Sarmiento's own words to express the widespread feelings towards these revolutionary biological ideas. In 1882, he expressively said "…Y yo, señores, adhiero a la doctrina de la evolución así generalizada, como procedimiento del espíritu, porque necesito reposar sobre un principio armonioso y bello a la vez, a fin de acallar la duda que es el tormento del alma" (Sarmiento 1951, p. 118). ("And I, gentlemen, adhere to the theory of evolution thus generalized, as a procedure of the spirit, because I need to rest upon a harmonious as well as beautiful principle, in order to hush doubt, that torment of soul"). Darwin died in 1882 after a long illness; it was not realized until after his death that he had suffered from Chagas's disease, which he had contracted while visiting South America. Although he was not the only originator of the evolution hypothesis, he certainly was the first man of science that gained for such theory a wide acceptance among biological experts. By contributing his own ideas on natural selection to the evolutionism outlined by Erasmus Darwin -his grandfather-, Lamarck and other biologists, he raised the evolutionary hypothesis to a provable theory.
ACKNOWLEDGEMENTS
I wish to acknowledge the assistance of my friend and fellow academician, Dr. Alfredo Cocucci, who supplied valuable information for the preparation of this article. Gonzalo Biarnes and librarian Sandra Ledesma, both members of the Academy's staff, kindly supplied the facsimiles herein included. Most important, my wife Elizabeth kindly helped out introducing needed corrections to the original manuscript.
WORKS CITED IN THE TEXT
1. Darwin, C. 1989. Voyage of the Beagle. Penguin Classics, 432 p., London.
2. García Castellanos, T. 1987. Breve historia de la Academia Nacional de Ciencias de Córdoba, República Argentina. Academia Nacional de Ciencias, Miscelánea 75, 39 p.
3. Herbert, S. 2005. Charles Darwin, geologist. Cornell University Press, 512 p., New York.
4. Lyell, C. 1998. Principles of Geology. Penguin Classics, 528 p., London.
5. Sarmiento, D.F. 1951. Obras completas, Discursos Populares. Ed. Luz del Día, 12, 382 p., Buenos Aires.
Darwin's observation in South America: what did he find at agua de la zorra, Mendoza province?
Stella Poma*, Vanesa D. Litvak *, Magdalena Koukharsky*, E. Beatriz Maisonnave and Sonia Quenardelle*
Laboratorio de Petrología, Departamento de Ciencias Geológicas, FCEN (UBA) y *CONICET. Emails: stella@gl.fcen.uba.ar; vane@gl.fcen.uba.ar; mkou@gl.fcen.uba.ar; beatriz@gl.fcen.uba.ar; sonia@gl.fcen.uba.ar
ABSTRACT: Scarcely 23 km from Uspallata, along the track of the old national highway 7, lies the district of Agua de la Zorra, in Mendoza province in western Argentina. Charles Darwin visited the area during his South American journeys in the 19th century and discovered a geological sequence that contained a paleoflora never described before. The flora includes an important number of species, particularly what is considered a small conifer forest with many silicified trunks still in life position. Darwin described and interpreted the sequence as sedimentary; his records show a very detailed level of observation. He also wondered about the processes that would cause the burial of the paleoflora, which he considered had happened in a marine sedimentary environment. In the modern geological framework and after a detailed study of the rocks containing the trunks, it is now interpreted that the conifer forest was buried by pyroclastic flows. Darwin accurately described the fine volcanic materials as an essential part of the deposit, but the key of the enigma about the origin of the deposits and the burial of the forest is the identification of the pyroclastic flow features; these were unknown process at the time of Darwin's observations and interpretation.
Keywords: Darwin Forest; Pyroclastic flow; Triassic; Agua de la Zorra; Mendoza province.
RESUMEN: Las observaciones de Darwin en Sudamérica: ¿Qué encontró en Agua de la Zorra, provincia de Mendoza? A escasos 23 km de Uspallata, a lo largo de la antigua ruta nacional 7, se encuentra la localidad de Agua de la Zorra en la provincia de Mendoza al oeste de Argentina. La región fue explorada por Charles Darwin durante su viaje por Sudamérica en el siglo XIX. Allí descubrió una secuencia geológica conteniendo una paleoflora que no había sido descripta hasta entonces. La flora incluye un importante número de especies y particularmente, un pequeño bosque de coníferas, los que se encuentran silicificados, y en posición de vida. Darwin describió e interpretó la secuencia como sedimentaria; sus notas y descripciones muestran un alto nivel de detalle en las observaciones. Darwin además se cuestionó acerca de los procesos que habrían causado el sepultamiento de la paleoflora, la que de acuerdo a sus deducciones ocurrió en un ambiente sedimentario marino. En el contexto geológico moderno y como consecuencia de un estudio detallado de las rocas que contienen los troncos, se interpreta que el bosque de coníferas fue sepultado por flujos piroclásticos. Darwin fue acertado en describir material volcánico como constituyente esencial del depósito, pero la clave del enigma acerca del origen de la secuencia y el soterramiento del bosque radica en la identificación de las características del depósito. Corresponden a acumulaciones producto del volcanismo fragmentario; que incluye un conjunto de procesos desconocidos al tiempo de las observaciones e interpretación de Darwin.
Palabras clave: Bosque de Darwin; Flujo piroclástico; Triásico; Agua de la Zorra; Provincia de Mendoza.
INTRODUCTION
Charles Darwin travelled around South America between 1832 and 1835. While staying at Valparaiso - one of Chile's most important seaports - during July 1834, HMS Beagle underwent repairs and Captain FitzRoy faced some financial difficulties. This situation provided Dar-win with some spare time and at his expense took a six week land expedition to the east, i.e., the High Andes in Chile and Argentina (Fig. 1). During this journey, Darwin recorded everything he found. In Chapter VII of one of his journals (Darwin 1846) he described the cordilleran geology, particularly the Agua de la Zorra region in Uspallata in Mendoza province. There he found a thick sequence, which he considered sedimentary, that contained some trunks never described before. The objective of this work is to revise Darwin's XIX century observations at Agua de la Zorra area in the modern geological framework.
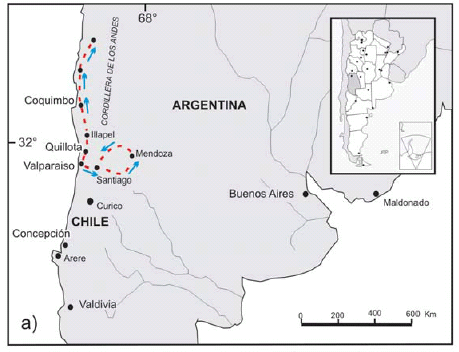
 Figure 1: a) Path followed by Darwin from Valparaiso, towards the north in Chile, and towards the east in Argentina, where he visited the Precordillera in Mendoza. b) Geographical location of Darwin Forest in the Agua de la Zorra area, in Uspallata, Mendoza province.
Figure 1: a) Path followed by Darwin from Valparaiso, towards the north in Chile, and towards the east in Argentina, where he visited the Precordillera in Mendoza. b) Geographical location of Darwin Forest in the Agua de la Zorra area, in Uspallata, Mendoza province.
DARWIN'S OBSERVATIONS
First observations of Darwin in the region of Agua de la Zorra in Uspallata are mentioned in Chapter VII (Darwin 1846), where he describes the area and refers to the paleoflora found as: "… I counted the stumps of fifty-two trees. They projected between two and five feet above the ground, and stood at exactly right angles to the strata, which were here inclined at an angle of about 25° to the west". One of the aspects that amazed him was that the trunks were in life position and well preserved. He not only described his finding but also asked advice from specialists about the type of flora: "…Eleven of these trees were silicified and well preserved: Mr. R. Brown has been so kind as to examine the wood when sliced and polished; he says it is coniferous, partaking of the characters of the Araucarian tribe, with some curious points of affinity with the Yew". His records show a very detailed level of observation: "All the stumps have nearly the same diameter, varying from one foot to eighteen inches; some of them stand within a yard of each other; they are grouped in a clump within a space of about sixty yards across". "They all stand at about the same level. The longest stump stood seven feet out of the ground: the roots, if they are still preserved, are buried and concealed. No one layer of the mudstone appeared much darker than the others, as if it had formerly existed as soil". Paragraphs ahead, he wondered about the processes that may have explained the origin of the deposits and the associated paleoflora: "Certainly the upright trees have been buried under several thousand feet in thickness of matter, accumulated under the sea. As the land, moreover, on which the trees grew, is formed of subaqueous deposits, of nearly if not quite equal thickness with the superincumbent strata, and as these deposits are regularly stratified and fine-grained, not like the matter thrown up on a sea-beach, a previous upward movement, aided no doubt by the great accumulation of lavas and sediment, is also indicated". He interpreted a possible origin for the deposits taking into account the regional geological setting known then: "At first I imagined, that the strata with the trees might have been accumulated in a lake: but this seems highly improbable; for, first, a very deep lake was necessary to receive the matter below the trees, then it must have been drained for their growth, and afterwards re-formed and made profoundly deep, so as to receive a subsequent accumulation of matter several thousand feet in thickness. And all this must have taken place necessarily before the formation of the Uspallata range, and therefore on the margin of the wide level expanse of the Pampas! Hence I conclude that it is infinitely more probable that the strata were accumulated under the sea".
STRATIGRAPHIC SETTING OF DARWIN'S DISCOVERY
Darwin's discovery at Agua de la Zorra is located along the old National Highway 7, which links Caracoles de Villavicencio with the town of Uspallata (Fig. 1). The best exposures occur about 800 m to the east of Agua de la Zorra locality (32° 28'39" S and 69°09'09" W). It was only in 1910 that the deposits where Darwin's paleoflora was found were described again by Stappenbeck (1910) and later by Harrington (1941, 1971). Figure 2 shows a schematic geological map of the area. The sequence containing the trunks, sedimentary in origin as Darwin described, is partially covered by alkaline basalts and intruded by sills of similar composition.
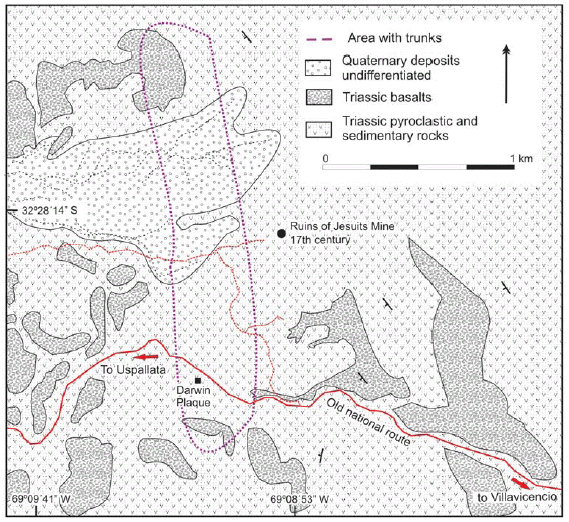 Figure 2: Schematic geologic map of the Agua de la Zorra region in Uspallata, Precordillera of Mendoza, Argentina.
Figure 2: Schematic geologic map of the Agua de la Zorra region in Uspallata, Precordillera of Mendoza, Argentina.
The abundant remains of conifers render it a classic site not only from the paleontological point of view, but also from a geological and historical perspective. Thus, the name of Bosque de Darwin (Darwin's Forest) for the site was formalized in the geological reports of Map 3369- 09, Uspallata (Brodtkorb et al. 1997, Cortés et al. 1997). The geological record of the region was studied by several authors, who also agreed a sedimentary origin for the deposits described by Darwin (Stipanicic 1972, 1979, Rolleri and Criado 1986, Strelkov and Alvarez 1984, Kokogian and Mancilla 1989, Ramos and Kay 1991, Morel 1994, Cortés et al. 1997, Kokogian et al. 1999). Stratigraphicaly, Cortés et al. (1997) identified the deposits described by Darwin as part of the Triassic Paramillo Formation within the Uspallata Group. In a recent analysis of a section of the sequence, Poma et al. (2004) described the volcaniclastic characteristics of the rocks containing the trunks; these are still under study. The area of Agua de la Zorra is part of the Cuyo Triassic extensional basin. According to Kokogian et al. (1999) the Cuyo Basin evolved through three depositional sequences that characterized different stages of rift evolution. The lowermost deposits, Early Triassic in age, were included in the synrift I stage by Kokogian et al. (1999) which is composed of coarse-grained conglomerates and breccias deposited in alluvial fan areas. These coarse-grained rocks are commonly found close to positive areas and basinwards taper into fluvial sandy ephemeral braided deposits and finally pass into muddy playa-lake sequences in the central parts of the basin. Isolated levels of rhyolitic tuffs occur at some places. The second sequence, labelled as the synrift II stage (Kokogian et al. 1999), was very probably deposited during the Middle Triassic and shows a wider areal distribution that the above considered synrift I. During this time prevailed different types of high-sinuosity river deposits composed of cross-bedded sandstones, conglomerates and mudstones. Lacustrine and deltaic sediments also appear in this section and indeed are dominant at the uppermost level of this section. Lake deposits are made up of interbedded sandstones-mudstones sequences with sporadic intercalations of organic-rich shales. A noticeable feature of this interval is the occurrence of abundant pyroclastic levels covered by thick basaltic lava flows and sills included in the uppermost part of the synrift II stage. Finally, during the Late Triassic a sag phase sequence was defined by Ramos and Kay (1991) and Kokogian et al. (1999), which characterized the final stages of subsidence in the basin under limited tectonic and magmatic activity. At this time prevailed sandstones and conglomerates sedimented by anastomosed and braided rivers that were covered by transgressive mudstones, shales and tuffs deposited in lacustrine environments.
GEOLOGY OF THE AGUA DE LA ZORRA REGION
Volcaniclastic deposits
In the Agua de la Zorra region, as Darwin described, there are a series of rocks containing tree trunks typical of coniferous - including Araucarioxylon sp. among them (Brea 1997, 2000) - and buried in life position. As shown in figure 3a, b, and c, Darwin's paleoflora is contained in a sedimentary/volcaniclastic sequence covered by alkaline basalts, both of Triassic age. According to field and microscope observations the material interbedded with the fossils comprises two main deposit types that are repeated with different thickness along the sequence.


 Figure 3: a) General view to the SW of the Triassic pyroclastic deposits covered by Triassic alkaline basalts; b) Closer view of the trunk, paleosoil, roots, and pyroclastic sequence including basal levels; c) Detailed view of the trunk, approximate diameter 40 cm.
Figure 3: a) General view to the SW of the Triassic pyroclastic deposits covered by Triassic alkaline basalts; b) Closer view of the trunk, paleosoil, roots, and pyroclastic sequence including basal levels; c) Detailed view of the trunk, approximate diameter 40 cm.
One of them is a characteristic pyroclastic flow deposit with crystal fragments, glass shards, pumice lapilli, all of them magmatically fragmented juvenile clasts product of their explosive origin. The crystals are small and irregular quartz grains and tabular or irregular fragments of oligoclase-andesine plagioclase. The matrix contains abundant angular glass shards, most of which have cuspate shape (Fig. 4a, and b). Fine ash fills the interstices between the shards. The shards are characteristically undeformed and some of them retain original bubble wall shapes, suggesting that the deposition was at moderate temperature. The lapilli pumices are concentrated at the lower part of the deposits and are smaller than 16 mm (medium lapilli). Occasionally it is possible to find silicified plant sprigs.
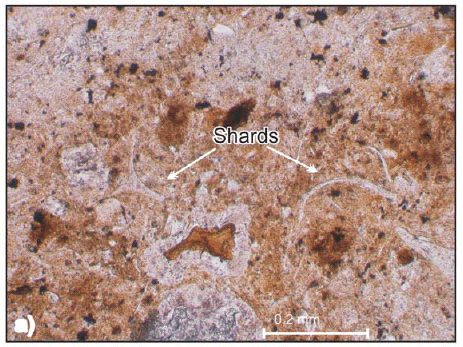
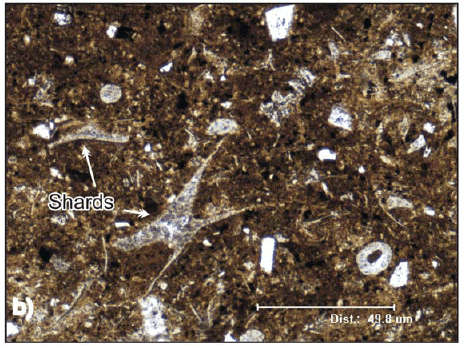 Figure 4: Thin section pictures of Paramillo Formation tuffs. a) Rhyolitic vitric tuff with undeformed shards (Parallel light); b) Rhyolitic vitric tuff with triaxon shards and rounded palagonite fragments of basaltic origin (Cross polarized light).
Figure 4: Thin section pictures of Paramillo Formation tuffs. a) Rhyolitic vitric tuff with undeformed shards (Parallel light); b) Rhyolitic vitric tuff with triaxon shards and rounded palagonite fragments of basaltic origin (Cross polarized light).
The rock is classified as a rhyolitic to dacitic medium lapilli tuff (White and Houghton 2006) with thin (decimetre scale) variably diffuse layering, thickness varying between 50 and 20 centimetres. They are defined by gradational grain size between pumice and lapilli. The set repeats itself several times along the sequence. Typically the diffuse stratification is subparallel with local gradual thinning. The rock commonly grades vertically into massive beds. Associated with the tuff and separated by a clearly noticeable surface there is a thin (centimetre to few decimetre scale) and well sorted deposit. It is poorly lithified and is interpreted as an air fall deposit that accompanied the emplacement of pyroclastic flows. It was preserved because a later ignimbritic flow covered it and protected it from erosion. The conspicuous features in these rocks are pyroclastic cuspate glass shards and pumice fragments. The shards and delicate spines of bubble walls were deposited undeformed, they indicate that the deposit is primary and has not been reworked from primary deposits (Fig. 4a, and b). The glassy shards are now transformed in zeolite and silica material. In the area, some massive lapilli tuff show normal vertical grading patterns defined by more or less gradual changes in grain size. Vertical grading typically occurs across a few decimetres. The record begins (Fig. 5) with a coarse lapilli tuff (White and Houghton 2006) (grain size average between 16 to 64 mm of pumice fragments), crystal fragments and lithics with minor amounts of glassy matrix (<10%), gradually changing to fine lapilli without or with only few pumice fragments. At the base these rocks show channel morphology probably indicative of some erosive behaviour by the flow. In this deposit, generally at lower levels, it is possible to notice small gas escape pipes. These structures are richer in pumice lapilli, lithics and poorer in fine ash than the enclosing tuff. This is characteristic of the removal of fine ash as a result of elutriation by gas escaping through the uncompacted deposit. The pipes were formed simultaneously with the flow accumulation, again illustrating the primary nature of the deposit. The pyroclastic rocks are intercalated with fluvial lenses of fine sandstones to conglomerate, interbedded with shales.
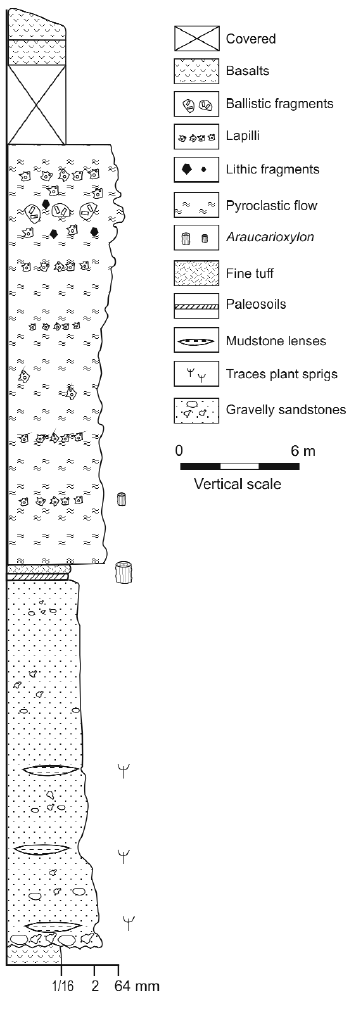 Figure 5: Lithological stratigraphic section of Paramillo Formation of Uspallata Group near Darwin's monument.
Figure 5: Lithological stratigraphic section of Paramillo Formation of Uspallata Group near Darwin's monument.
Geomorphology and patterns of thickness variation of the pyroclastic deposits suggest a possible western provenance even though there is no conclusive evidence.
Tuff-basalts transition
The pyroclastic and sedimentary rocks described are intercalated and covered with Triassic mafic rocks. These are sills and lava flows of basaltic composition (Massabie 1986, Ramos and Kay 1991). In the Agua de la Zorra region the olivine-bearing basalts are easily noticeable because of their dark colours, fine grain and scoriaceous or amygdaloidal textures. The vesicles were partially filled with calcium carbonate and silica and zeolites in subordinate amounts. These materials are conspicuous in altered basalts and sometimes are found in pyroclastic and sedimentary rocks. The lower part of the basalts are resting over a poorly consolidate rhyolitic tuff that contains small spheres of basaltic palagonite lava. The size of the spheres is variable but no larger than 5 cm. They represent the interaction between lava flows and unconsolidated water soaked material. Upwards there are peperites that result from the mingling between basalt, rhyolitic tuff and clastic sediments in variables proportions. These textures suggest that the basalts were produced shortly after or simultaneously with the pyroclastic activity and sedimentation. There were not identified volcanological structures that could relate the rhyolitic pyroclastic deposits with the basaltic lava flows, seeming the latter the result of fissure eruptions.
CHARACTER AND ORIGIN OF THE DEPOSITS
Just as Darwin described, the trunks are arranged vertical to the layers. They stand at approximately the same level, and although Darwin did not identify a normal paleosoil, he understood that these trees had to grow in a dry substrate. He mentioned (Darwin 1846): "As the trees obviously must once have grown on dry land, what an enormous amount of subsidence is thus indicated! Nevertheless, had it not been for the trees, there was no appearance which would have led any one even to have conjectured that these strata had subsided". Obviously, he was amazed at the great thickness of sediments that there are in the sequence, which hinders any simple interpretation on the origin of the set. Finally, Darwin makes up his mind and chooses the simplest interpretation, even though this is not satisfactory. "That it is infinitely more probable that the strata were accumulated under the sea". On the other hand he observes and describes that the fine material plastered around the trunks as a package preserves the mould with the external texture of the plants: "The bark round the trunks must have been circularly furrowed with irregular lines, for the mudstone round them is thus plainly marked". According to the observed and previously described characteristics of the deposits, such as their areal distribution, changing thickness, flow nature, type of structures and textures, we interpret that the deposit that encloses Darwin's Flora in the area of Agua de la Zorra was originated as a consequence of a phreatomagmatic eruption (Poma et al. 2004). The key of the enigma about the origin of the deposits rests in the identification of the pyroclastic features of the rocks containing the trunks, particularly their phreatomagmatic origin; this was an unknown process at the time of Darwin's observations and interpretation (Darwin 1844). Along geologic time and humankind history there are numerous examples of flora, fauna and populations buried by this type of processes. Seldom were human witnesses present while volcanic activity was taking place. In those rare cases, men and women were able to describe the catastrophic episode. Pompeii (79 BC), for example, was buried under four meters of ash and debris. Pliny the Younger, a Roman soldier, witnessed much of the eruption (Francis 1993). Pompeii was covered over with a hot cloud of volcanic ash, steam and some mud material. The pyroclastic cloud flowed down the side of the volcano at a speed of 30 meters per second (Francis 1993). It took less than four minutes for the pyroclastic flow to travel from Vesuvius to Pompeii a few kilometres away. In 1815 more than 100,000 people died when Mount Tambora erupted on the Indonesian island of Sumbawa (Sutawidjaja et al. 2006). Despite the presence of witnesses in Mount Tambora and the discovery of Pompeii, among other eruptions, in Darwin's time advanced knowledge about the mechanism of this volcanic activity was limited. It was only during the last century and after many eruptions that pyroclastic flows were thoroughly studied and documented by hundreds of volcanologists worldwide, such as the 1980 Saint Helens among others. With detailed monitoring and modeling of active volcanism, eruption styles, type of ejecta, flows and clouds were understood and only then past volcanic expressions could be properly interpreted. In the same way it was only recently that science acknowledged the significant impact of these types of phenomena on climatic change at a planetary scale. A phreatomagmatic eruption was also responsible for the burial of Darwin`s forest in Agua de la Zorra. In continental volcanic environments the interaction between hot magma and water frequently produces this kind of volcanism, in which a voluminous explosive eruption of magmatic ejects emulsionated with magmatic and external volatiles. Figure 6 shows an interpretation of the eruption. The characteristics of the pyroclastic sequence of Agua de la Zorra allow us to make another interpretation: The eruption was not destructive (at least not in that sector) as the trees remain in life position; so it is inferred the eruption was not laterally directed and can be supposed to be vertical. However, there is no concluding evidence to disregard a lateral eruption, especially if the deposits are distal or out of the lateral blast influence.
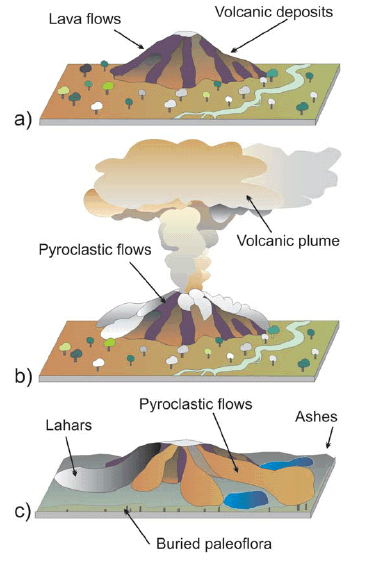 Figure 6: Schematic representation of a three stage evolution for the Darwin's Forest burial. a) Inactive volcanic center surrounded by the forest; b) Pyroclastic eruption with associated volcanic plumes and pyroclastic flows reaching Darwin's Forest; c) Darwin's Forest is buried by the pyroclastic flows and ashes. Lahars recognized by Brodtkorb et al. (1997) are included in the sketch.
Figure 6: Schematic representation of a three stage evolution for the Darwin's Forest burial. a) Inactive volcanic center surrounded by the forest; b) Pyroclastic eruption with associated volcanic plumes and pyroclastic flows reaching Darwin's Forest; c) Darwin's Forest is buried by the pyroclastic flows and ashes. Lahars recognized by Brodtkorb et al. (1997) are included in the sketch.
Volcanic activity was intense enough to eliminate most of the soft tissues of the flora such as leaves, which are more abundant in the sedimentary levels. Burial must have been in a short time, as a consequence of one or more eruptions but close in time, because there are no recorded breaks between the flows that cover the trunks. Only one paleosoil was observed below a tree (Fig. 3b). Its thickness is less than 50 cm. It is dark with root marks. It was covered by a thin and diluted flow with low particle/gas ratio; it possibly was a base surge above which a series of pyroclastic flows accumulated. They are stratified and the sequence ends with an ash fall, possibly related to the co-ignimbritic cloud. There is no clear evidence of an extended ash fall deposit besides the coignimbritic cloud deposits already mentioned. In that regard, the eruptive column is not considered to be too high. As indicated by Sparks and Wilson (1982), a high proportion of steam in phreatomagmatic eruptions results in shorter columns. Changing thickness and varying structures and textures seem to imply an unstable feeding system from the magma chamber to the eruptive column. Instability could have been the consequence of the variations in the magma supply and in any case is responsible for the recorded duality in the massive form or diffusely stratified deposits. This interpretation is consistent with a first deposit in the area made up of initial base surge such as the ones locally observed overlying the paleosoil. Eruptions of these characteristics are violent and also of rapid deposition. This amount of ash had already surprised Darwin who, as we mentioned earlier, had interpreted it as material of sedimentary origin.
CONCLUSIONS
The value of Darwin's work is the accuracy of the descriptions and the detailed degree of his observations, which are still valid nowadays. He was successful in the interpretation that the forest was buried in life. Despite Darwin misinterpreted the origin of the deposits as sedimentary, he was accurate in describing the fine volcanic materials as an essential part of them, although he never considered the eruptive process as the one responsible for the burial. The present interpretation as a phreatomagmatic mechanism is now plausible because of the advances in volcanological knowledge and understanding during the latter years of the Twentieth Century.
ACKNOWLEDGEMENTS
This work was funded by University of Buenos Aires grant UBACYT X207. The authors are particularly grateful to S. Kay, E. Llambías and P. Sruoga for their review of the manuscript and to Miguel Griffin for his valuable suggestions to improve the English version.
WORKS CITED IN THE TEXT
1. Brodtkorb, A., Cortés, J.M., González Bonorino, G. and Koukharsky, M. 1997. Sitios de interés geológico correspondientes a la Hoja 3369-09, Uspallata, provincia de Mendoza, SEGEMAR, unpublished, 40 p. Buenos Aires.
2. Brea, M. 1997. Una nueva especie fósil del género Araucarioxylon Kraus 1870 emend. Maheshwari 1972 del Triásico de Agua de la Zorra, Uspallata, Mendoza, Argentina. Ameghiniana 34: 485-496.
3. Brea, M. 2000. Paleoflora triásica de Agua de la Zorra, Uspallata, provincia de Mendoza, Argentina: Lycophyta y Filicophyta. Ameghiniana 37: 199-204.
4. Cortés, J.M., González Bonorino, G., Koukharsky, M., Pereyra, F. and Brodtkorb, A. 1997. Hoja geológica 3369-09 Uspallata, provincia de Mendoza. SEGEMAR, (unpublished report) 165 p., Buenos Aires.
5. Darwin, C. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 280 p., London.
6. Darwin, C. 1844. Geological observations on the volcanic islands visited during the voyage of H.M.S. Beagle. Smith, Elder and co., 176 p., London.
7. Francis, P. 1993. Volcanoes, a planetary perspective. Clarendon Press. 443 p., New York.
8. Harrington, H.J. 1941. Investigaciones geológicas en las sierras de Villavicencio y Mal País, provincia de Mendoza. Dirección Nacional de Geología y Minería, Boletín 49, 54 p., Buenos Aires.
9. Harrington, H.J. 1971. Descripción geológica de la Hoja 22c, "Ramblón", provincias de Mendoza y San Juan. Dirección Nacional de Geología y Minería, Boletín 114, 81 p., Buenos Aires.
10. Kokogian, D.A. and Mancilla, G.H. 1989. Análisis estratigráfico y secuencial de la Cuenca Cuyana. In Chebli, G. and Spalletti, L. (eds.) Cuencas Sedimentarias Argentinas. Facultad de Ciencias Naturales, Universidad Nacional de Tucumán, Serie Correlación Geológica 6: 169-201.
11. Kokogian, D.A., Spalletti, L., Morel, E., Artabe, A., Martínez, R.N., Alcober, O.A., Milana, J.P., Zavattieri, A.M. and Papú, O.H. 1999. Los depósitos continentales triásicos. In Caminos, R. (ed.) Geología Argentina, Instituto de Geología y Recursos Naturales, Anales 29(15): 377-398, Buenos Aires.
12. Massabie, A. 1986. Filón capa Paramillos de Uspallata, su caracterización geológica y edad, Paramillo de Uspallata, Mendoza. Primeras Jornadas sobre Geología de la Precordillera, Asociación Geológica Argentina, Serie A, 2: 325-330.
13. Morel, E.M. 1994. El Triásico del cerro Cacheuta, Mendoza (Argentina) Parte I: geología, contenido paleoflorístico y cronoestratigrafía. Ameghiniana 31: 161-176.
14. Poma, S., Quenardelle, S. and Koukharsky, M. 2004. Un flujo piroclástico triásico en la Formación Paramillo, provincia de Mendoza. In Brodtkorb, M. K. De, Koukharsky, M., Quenardelle, S. and Montenegro, T. (eds.) Avances en Mineralogía, Metalogenia y Petrología 2004, 385-390, Río Cuarto.
15. Ramos, V.A. and Kay, S. 1991. Triassic rifting and associated basalts in the Cuyo Basin, central Argentina. In Harmon, R.S. and Rapela, C.W. (eds.) Andean Magmatism and its Tectonic Setting, Geological Society of America, Special Paper 265: 79-91.
16. Rolleri, E.O. and Criado, R.P. 1968. La Cuenca Triásica del norte de Mendoza. Terceras Jornadas Geológicas Argentinas (Comodoro Rivadavia), Actas 1: 1-76.
17. Sparks, R.S.J. and Wilson, L. 1982. Explosive volcanic eruptions- V. Observations of plume dynamics during the 1979 Soufrière eruption, St. Vincent. Geophysical Journal Internacional 69: 551-570.
18. Stappenbeck, R. 1910. La Precordillera de San Juan y Mendoza. Ministerio de Agricultura de la Nación, Sección Geología, Mineralogía y Minería, Anales 4(3): 3-187, Buenos Aires.
19. Stipanicic, P.N. 1972. La Cuenca Triásica de Barreal (provincial de San Juan). In Leanza, A.F. (ed.) Geología Regional Argentina, Academia Nacional de Ciencias: 537-566, Córdoba.
20. Stipanicic, P.N. 1979. El Triásico del Valle del Río de Los Patos (provincia de San Juan). In Turner, J.C.M. (ed.) Geología Regional Argentina, Academia Nacional de Ciencias 1: 695-744, Córdoba.
21. Strelkov, E.E. and Alvarez, L.A. 1984. Análisis Estratigráfico y Evolutivo de la Cuenca Triásica Mendocina - Sanjuanina. 9° Congreso Geológico Argentino (Bariloche), Actas 3: 115-130.
22. Sutawidjaja, I.S., Sigurdsson, H. and Abrams, L. 2006. Characterization of volcanic deposits and geoarchaeological studies from the 1815 eruption of Tambora volcano. Jurnal Geologi Indonesia 1(1): 49-57.
23. White, J.D.L. and Houghton, B.F. 2006. Primary volcaniclastic rocks. Geology 34: 677-680.
Darwin forest at agua de la zorra: the first in situ forest discovered in South America by Darwin in 1835
Mariana Brea1,4, Analía E. Artabe2,4 and Luis A. Spalletti3,4
1Laboratorio de Paleobotánica, Centro de Investigaciones Científicas, Diamante. CICYTTP-Diamante, Consejo Nacional de Investigaciones Científicas y Técnicas, Diamante, Entre Ríos, Email: cidmbrea@infoaire.com.ar 2División Paleobotánica, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata. Email: aeartabe@museo.fcnym.unlp.edu.ar 3Centro de Investigaciones Geológicas (CONICET-UNLP), La Plata. Email: spalle@cig.museo.unlp.edu.ar 4Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET).
ABSTRACT
The Agua de la Zorra area (near Uspallata, Mendoza, Argentina) is one of the best renowned fossil localities of the country because of its spectacular in situ fossil forest. This forest was discovered by Charles Darwin in 1835, who described this forest as monotypic and assigned it a Tertiary age. Nowadays, this fossil locality is known as the Darwin Forest. Over a century and a half later it was reinterpreted as a mixed Middle Triassic forest and a new fossil monotypic palaeocommunity of horsetails was discovered. This palaeovegetation is included in the Paramillo Formation (i.e., lower section the Potrerillos Formation) of northwestern Cuyo Basin, Mendoza province (69°12' W and 32°30' S). The sediments were deposited in a sinuous fluvial system, in which channel-filling sand bodies were associated with mud-dominated floodplain deposits. The palaeoforest grew on an andisol soil that developed on volcaniclastic floodplain deposits. It had a density of 427 -759 trees per hectare, and was constituted by conifers and corystosperms distributed in two arboreal strata. The highest reached 20-26 m tall, and was dominated by corystosperms, but it also included the tallest conifers. The second stratum, mainly composed of conifers, ranged between 16-20 m tall. The forest has also emergent corystosperms, which reached 30 m tall. The understorey was composed of ferns. Growth ring anatomy suggests that conifers could have had an evergreen habit. Structure of vegetation, growth ring analyses and sedimentation suggest that the forest developed under dry, subtropical, and strongly seasonal conditions.
Keywords: Palaeovegetation; Palaeoecology; Palaeoenvironments; Middle Triassic; Southwestern Gondwana.
RESUMEN: El Bosque Darwin en Agua de la Zorra: El primer bosque in situ descubierto en América del Sur por Darwin en 1835. El área de Agua de la Zorra (cerca de Uspallata, Mendoza, Argentina) es uno de los sitios fósiles más espectaculares y renombrados del país porque aflora un bosque in situ. Este bosque descripto como monotípico y asignado al Terciario fue descubierto por Charles Darwin en 1835. Un siglo y medio más tarde, se reinterpretó como un bosque mixto del Triásico Medio y se reconoció una nueva paleocomunidad monotípica de esfenófitas. La paleovegetación proviene de la Formación Paramillo (= sección inferior de la Formación Potrerillos) de la región noroeste de la Cuenca Cuyana, provincia de Mendoza (69°12'O y 32°30'S). Los sedimentos fueron depositados por un sistema fluvial de alta sinuosidad. El bosque fósil creció sobre suelos del tipo andisol que se desarrollaron sobre depósitos de planicies volcaniclásticas. El bosque Darwin tiene una densidad de 427-759 árboles por hectárea. Este está constituido por coníferas y corystospermas y tiene dos estratos arbóreos. El más alto se desarrolla entre los 20-26 m, y es dominado por corystospermas, pero también por las coníferas más altas. El segundo estrato arbóreo, principalmente compuesto de coníferas, con un rango entre 16-20 m. El bosque tiene también algunos emergentes de corystospermas las cuales alcanzan hasta 30 m de altura. El sotobosque estaba compuesto de helechos. La anatomía de los anillos de crecimiento sugiere que las coníferas podrían haber tenido hábito siempreverde. La estructura de la vegetación, el análisis de los anillos de crecimiento y la sedimentología sugieren que este bosque se desarrollo bajo condiciones climáticas subtropicales, secas y estacionales.
Palabras clave: Paleovegetación; Paleoecología; Paleoambientes; Triásico Medio; Sudoeste de Gondwana.
INTRODUCTION
The first fossil plants in South America were recorded by European naturalists during the 18th and 19th centuries (Ottone 2005). The British naturalist Charles Robert Darwin (1809- 1882) was the first who provided palaeontological and geological observations of the Agua de la Zorra area, Uspallata, near Villavicencio, Mendoza province in Argentine territory (Darwin 1839a, 1839b, 1845, 1846). Later, this famous fossiliferous locality was visited by other European naturalistexplorers such as Burmeister 1861, Doering 1882, Stelzner 1885, Avé-Lallemant 1891 (see Ottone 2005). The expeditions of Charles Darwin -on board HMS Beagle- during his historic journery around the world between 1831 and 1836, contributed valuable information and observations on the geology, plant and animal fossils, and extant organisms of South America. Darwin also collected a huge number of specimens and samples, many of them new to science. Darwin arrived in Argentina in 1833 and visited the Río de La Plata area, La Bajada (now Paraná city, Entre Ríos), the Santa Cruz River and many other sites in Patagonia and Tierra del Fuego. Then, after crossing the Cordillera de los Andes from Chile, he arrived in Mendoza province and discovered the first in situ forest in South America, at Agua de la Zorra, located about 25 km from the town of Uspallata (Conwentz 1885, Rusconi 1941, Harrington 1971, Brea 1995, Ottone 2005). He recorded the presence of 52 fossil tree trunks measuring 90-152 cm in diameter, buried in sandstones or volcanic sandstones and standing out as columns several meters high (Darwin 1846). This discovery by Darwin is remembered by a monolith (Fig. 1).

 Figure 1: a. Charles Darwin's tribute plaque at Agua de la Zorra, near Uspallata, Mendoza, Argentina. This photo was taken in 1993. b. Panoramic view of Triassic sequences, the arrow shows the plaque.
Figure 1: a. Charles Darwin's tribute plaque at Agua de la Zorra, near Uspallata, Mendoza, Argentina. This photo was taken in 1993. b. Panoramic view of Triassic sequences, the arrow shows the plaque.
On 30 March 1835 he collected the first specimens of fossil wood from Agua de la Zorra. The samples were sent to London and were referred by Robert Brown to the genus Araucarites (Darwin 1846). Recently, one of the authors (M. B.) carried out three research field trips as part of work towards her PhD dissertation. Brea (1995) characterized and studied the Triassic units at Agua de la Zorra from a palaeobotanical and sedimentological viewpoint. She defined two palaeocommunities in the Paramillo Formation, together with their associated palaeoenviroments (Fig. 2). The first paleocommunity, the in situ Darwin Forest (Fig. 2), appears at four fossiliferous sites (Brea 1995), while the second was found at only one locality and included only fossil horsetails (Brea and Artabe 1999, Brea et al. 2008).
 Figure 2: The Darwin Forest at Agua de la Zorra area (near Uspallata, Mendoza). a-b-c-e. Large fossil tree stumps in growth position in Triassic sequences; d-f-h. Petrified standing trees at fossiliferous sites C; g. Large fossil log exposed in Triassic sediments at fossiliferous site B.
Figure 2: The Darwin Forest at Agua de la Zorra area (near Uspallata, Mendoza). a-b-c-e. Large fossil tree stumps in growth position in Triassic sequences; d-f-h. Petrified standing trees at fossiliferous sites C; g. Large fossil log exposed in Triassic sediments at fossiliferous site B.
The Darwin Forest was first referred to the Tertiary by Darwin (1839a, 1839b), but new data now available suggest a late Middle Triassic age (Spalletti et al. 1999, Brea et al. 2008). The most important feature of the Darwin Forest is that the trees are still preserved at the sites on which they grew (Fig. 2); it is the best renowned of its age in southwest Gondwana. Although petrified forests in growth position are very scarce in the geological record, the Triassic fossil record of SW Argentina shows four in situ fossil forests (Paramillo, Cortaderita, Ischigualasto, and Río Blanco Formations) and their occurrences have contributed significantly to our understanding of Triassic Gondwana ecosystems (Zamuner 1992, Artabe et al. 2001, 2007a, Brea et al. 2008). The Darwin Forest was reinterpreted as a subtropical dry seasonal forest (Brea et al. 2005, 2008). It grew on an andisol soil that developed on volcaniclastic floodplain deposits. The volcanic detritus and the rhythmic amalgamation of upper flow-regime tractional deposits overlying the andisol indicate that the forest was buried rapidly by a subaerial, cool and wet pyroclastic base surge flow (Poma et al. 2004, Brea et al. 2008). The continental Triassic succession of southwestern Gondwana occurs in a series of narrow rifts produced as a result of Triassic continental extension. These rift basins are composed of a continental clastic infilling, and record complex interactions between alluvial, fluvial, deltaic and lacustrine depositional systems with intercalations of volcaniclastic sequences in most of these basins. The rich floristic record allowed recognition of several assemblages, biozones and stages characterized by floristic events (Spalletti et al. 1999, 2003, Artabe et al. 2001, 2003, 2007b, Morel et al. 2003). Over the last two decades, most investigations on the Argentinian Triassic have focused on the gross stratigraphy and taxonomy of fossil plants (Stipanicic 2001, Stipanicic and Marsicano 2002, Zamuner et al. 2001, and references therein). However, recently published studies (Spalletti et al. 1999, 2003, Artabe et al. 2001, 2003, 2007b, Morel et al. 2003) provided an increasing understanding of how the Triassic palaeofloras developed and changed over time as a response to variations in depositional systems, tectonism and climate. The most spectacular fossil sites of the Paramillo Formation- in the Agua de la Zorra area -, are located in the northernwestern sector of the Cuyo Basin (between 32º and 36º SL). The Paramillo Formation is composed of a 140 m thick succession of clast-supported conglomerates, pebbly sandstones, tuffaceous sandstones and mudstones (Figs. 3 and 4). These deposits have been previously described by Harrington (1971), Strelkov and Álvarez (1984), Kokogian and Mancilla (1989), Ramos and Kay (1991), Massabie (1985), Massabie et al. (1985), Linares and González (1990), Ramos (1993), Kokogian et al. (1993). These authors all agreed in interpreting the Paramillo Formation as deposited in highly sinuous fluvial systems.
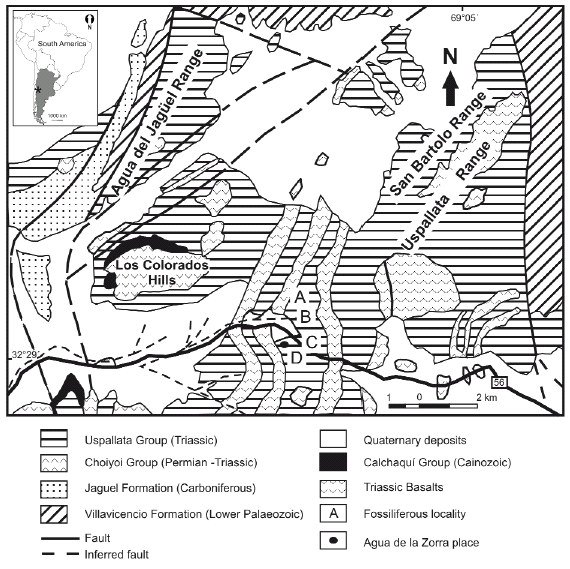 Figure 3: Location map showing Agua de la Zorra fossiliferous locality, Mendoza province, Argentina indicating the four fossiliferous sites: A, B, C, D respectively.
Figure 3: Location map showing Agua de la Zorra fossiliferous locality, Mendoza province, Argentina indicating the four fossiliferous sites: A, B, C, D respectively.
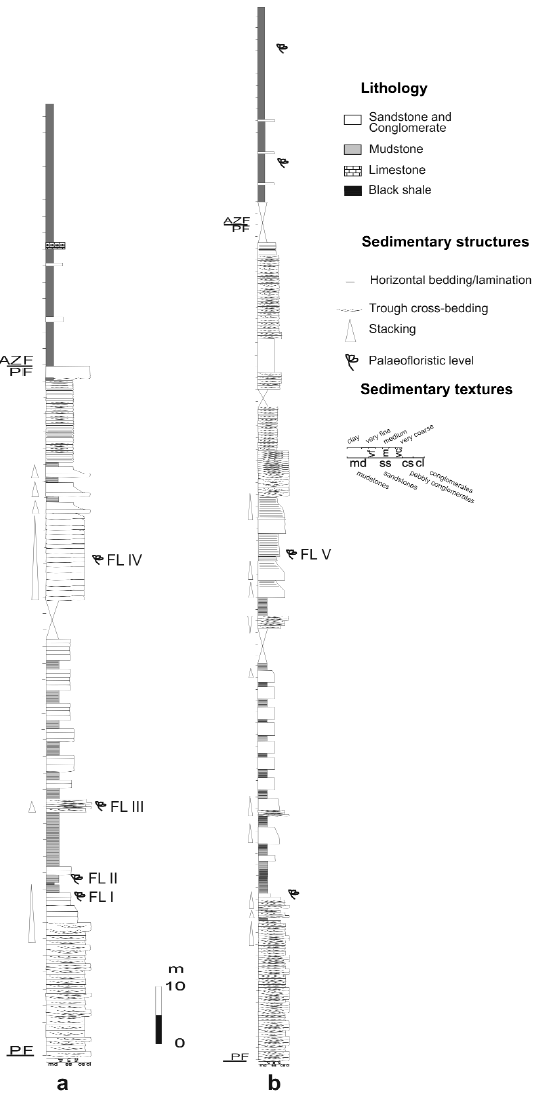 Figure 4: a. Lithostratigraphic section of the Paramillo and Agua de la Zorra Formations at locality A, showing the main lithofacies and the position of the fossil forest level. b. Lithostratigraphic section of the Paramillo and Agua de la Zorra Formations at locality C. PF: Paramillo Formation, AZF: Agua de la Zorra Formation. FL I - V: fossiliferous levels.
Figure 4: a. Lithostratigraphic section of the Paramillo and Agua de la Zorra Formations at locality A, showing the main lithofacies and the position of the fossil forest level. b. Lithostratigraphic section of the Paramillo and Agua de la Zorra Formations at locality C. PF: Paramillo Formation, AZF: Agua de la Zorra Formation. FL I - V: fossiliferous levels.
Previous palaeobotanical studies, comprising mostly lists of fossil plants, were published by Darwin (1846), Conwentz (1885), Stappenbeck (1910), Kurtz (1921), Du Toit (1927), Groeber (1939), Windhausen (1941), Harrington (1971) and Stipanicic et al. (1996), while modern systematic contributions were offered by Brea (1995, 1997, 2000), Brea and Artabe (1999), Artabe and Brea (2003) and Brea et al. (2005, 2008).
THE PARAMILLO FORMATION
The Paramillo Formation is a volcaniclastic unit composed of yellowish lithic sandstones, brownish and yellowish tuffaceous sandstones, dark gray and green shales and mudstones, and pink to reddish ash fall tuffs (Fig. 4). The sedimentary record of the overlying Agua de la Zorra Formation is dominated by bituminous shales and marls with subordinate intercalations of yellowish fine-grained sandstones and mudstones (Fig. 4). Both units are intruded by several sills of olivine diabase dated 235 + 5 Ma (K/Ar whole age) by Ramos and Kay (1991). At that time Mendoza was a part of the vast supercontinent called Gondwana and it was placed approximately at the same geographic latitude as it is nowadays. Brea (1995) studied in more detail the upper part of the Paramillo Formation and the lowermost Agua de la Zorra Formation and defined several lithofacies and facies associations. The Paramillo Formation consists of cross-bedded conglomerates, cross-bedded, plane-bedded and massive pebbly sandstones, cross-bedded, massive, plane bedded and ripple-laminated sandstones and intercalations of laminated mudstones and shales. Brea (1995) interpreted that these sediments were deposited in a highly sinuous (meandering) fluvial system, in which channel-filling sand bodies are associated with mud-dominated floodplain deposits. The cyclic arrangement of well preserved soil horizons and sedimentary deposits lacking evidence of soil formation indicates alternation of periods with strong sediment aggradation produced by non-channelized high-regime flows with periods characterized by very low accommodation rates that favoured the development of immature soil profiles (Brea et al. 2008). The volcanic nature of detrital components suggests that the highly aggrading non-channelized flows that produced the burial of the Darwin Forest could be related to pyroclastic events (Poma et al. 2004). The forest might have died as a result of a diluted, subaerial, cool and wet base surge pyroclastic flow (Poma et al. 2004, Brea et al. 2008). The Paramillo Formation, where the Darwin Forest emerges, has been correlated with the lower section of the Potrerillos Formation (Spalletti et al. 1999, Morel et al. 2003) on the basis of Assemblage Biozone chronology, analysis of main stratigraphic unconformities, and evolution of basin infill.
ECOLOGICAL RECONSTRUCTION OF THE DARWIN FOREST
The Triassic landscape at the Agua de la Zorra area was very different in comparison with the modern-day scenery. Sphenopsids dominated the flooded areas and conifers and corystosperms were the most important components of the arboreal vegetation. Four exposures of the Darwin Forest were found in the Paramillo Formation during fieldwork carried out in 1993-1994 (Figs. 2 and 3). This unit was thoroughly examined at two localities: Darwin and El Sauce (Figs. 3 and 4), where the lithology was logged bed-by-bed in vertical sections at a 1:100 scale (see Brea et al. 2008). Imperfect carbonization processes (Poma et al. 2004) preserved the stumps and in just over 2 km2 one hundred and twenty stumps in growth position and fallen logs were counted (Fig. 2). The fossiliferous levels (FL) with stumps were found at four localities belonging to the same in situ forest. Thus, FL IV (at A locality), FL V (at B and C localities) and FL VI (at D locality) belong to a single stratigraphic level (Figs. 2 and 4). Fossiliferous levels FL I and FL III found at locality A (Fig. 4) have ferns preserved as impressions-compressions (Brea 2000). The sedimentary sequences in which this forest is preserved consist of continental volcaniclastic units, resulting from deposition on a highly sinuous fluvial system associated with river flood-plains (Fig. 4). The palaeoecological reconstruction of the Darwin Forest was based on the quantitative data of the mapped forest (mean separation of trees, basal area per ha, species distribution) integrated with the taxonomic and sedimentological information (Brea et al. 2008). The mixed forest was composed by corystosperms (30-40%) and conifers (60-70%). The corystosperms are a group of extinct plants with mostly fern-like foliage with ovules borne on modified leaves or cupules. This group was the dominant seed plant in Gondwana during the Triassic (Stewart 1983). All conifers are a diverse group of trees and shrubs that underwent a major radiation during the Triassic period, when the first occurrence and radiation of the eight conifer families occurred (Willis and McElwain 2002). The conifers have a pyramidal arborescent growth form, with cone-bearing seed plants; the foliage is either needle-like or scale-like. The plants are mainly small, long, and thin arranged spirally and bearing the male and female reproductive organs in separate cones on different trees, or indifferent parts of the same. The average density of the tree stumps in this forest is 556 trees/ha. The spatial autocorrelation for the species variable indicates that in some places the corystosperms and conifers are intermingled while in others the corystosperms and conifers seem to be aggregated into cohesive social groups (Fig. 5). The Darwin Forest reveals two strata: the highest, developed between 20-24 m, has a preponderance of corystosperms but also the tallest conifers; the second stratum, mainly composed of conifers, ranges between 16-20 m. The forest also has emergent corystosperms which reach 30 m tall (Brea et al. 2008). The corystosperms were assigned by Artabe and Brea (2003) to Cuneumxylon spallettii and constitute the dominant species in the Darwin Foest.
 Figure 5: Reconstruction of the Triassic Darwin Forest landscape in a high sinuosity fluvial system, in which channel-filling sand bodies are associated with mud-dominated floodplain deposits. The canopy is integrated by two arboreal strata and emergent trees with conifers and corystosperms. The understorey is formed by ferns. Cinodonts are characteristic tetrapods during the Triassic of the Cuyo Basin (drawing by Jorge Gonzalez).
Figure 5: Reconstruction of the Triassic Darwin Forest landscape in a high sinuosity fluvial system, in which channel-filling sand bodies are associated with mud-dominated floodplain deposits. The canopy is integrated by two arboreal strata and emergent trees with conifers and corystosperms. The understorey is formed by ferns. Cinodonts are characteristic tetrapods during the Triassic of the Cuyo Basin (drawing by Jorge Gonzalez).
This fossil forest presents values of biomass and stand basal area comparable to those of the current subtropical seasonal forests (Brea et al. 2008). These forests develop under a climatic regime that includes an annual cycle with one season in which water is unavailable to plants because of lack of precipitation. The dry season alternates with another in which there is abundant water. In addition to structural data of the Darwin Forest that match those of an extant monsoon forest, the polyxyly found in Cuneumxylon has been considered an important adaptive wood character to avoid water stress. As suggested by the functional anatomy and distribution of living groups (Fahn 1990), the included phloem associated with great amounts of parenchyma could be a strategy of subtropical plants to fight against water stress in arid regions during drought seasons. During Permian-Triassic biosphere reorganization, aridity of the Earth increased and in the Triassic the Pangea was characterized by strongly seasonal climates in a warm-house period (Parrish et al. 1982, Dubiel et al. 1991, Parrish 1993, Scotese et al. 1999). The spreading of continental climates caused the extension of semiarid belts into middle latitudes and, partly, into high latitudes too (Chumakov and Zharkov 2003). Recently, general circulation models (GCMs) were developed by Sellowood and Valdes (2006) to simulate Mesozoic climatic patterns. In this scheme the Triassic Southwestern Gondwana is modelled as seasonal and winter-wet against the opinion of Robinson (1973) who stated that it was seasonal and summer-wet. Growth-ring analysis of coniferous wood - assigned by Brea (1997) to Araucarioxylon protoaraucana - was used to evaluate climatic conditions (Fritts 1976, Holmes 1985). Conifer growth rings are narrow and subtly demarcated; they are characterized by a relatively wide zone of large, thin-walled early-wood cells terminated by ten to seventeen thick-walled late-wood cells. The presence of narrow growth rings indicates poor conditions for cell division and expansion during growth season (Creber and Chaloner 1984). In addition, a thin late-wood zone within a ring can be the result of water shortage at the end of the growing season, a sharp photoperiod indicating end of growth season or abrupt leaf shedding. Therefore, A. protoaraucana growth rings suggest strongly seasonal conditions. The average ring width in these fossil woods is 1.49 mm (0.80-2.53 mm), the narrowest ring is 0.12 mm and the widest ring is 4.44 mm. Mean sensitivity (MS) values range between 0.14 and 0.38. Average MS values (0.30) indicate that the growing environment was stressed and not uniform. Although available data are limited, the complicated growth trends suggest that competition, disturbance events, or climatic stress influenced the growth of trees in the Darwin Forest. Furthermore, Falcon Lang's method (Falcon-Lang 2000a, 2000b) was used to distinguish between evergreen and deciduous species (see Brea et al. 2008). The quantitative growth ring anatomy analysis method (Falcon-Lang 2000a, 200b) indicates that the conifer fossil woods of the Darwin Forest probably belong to an evergreen gymnosperm. The understorey of the Darwin Forest includes Cladophlebis mesozoica Frenguelli 1947 (Fig. 6, b), Cladophlebis mendozaensis (Geinitz) Frenguelli 1947 (Fig. 6, c), and Cladophlebis kurtzi Frenguelli 1947 (Fig. 6 a) (Brea 2000). Cladophlebis is an extinct genus of ferns characterized by the presence of large sterile bipinnate foliage and it was widely distributed in southern Gondwana during the Mesozoic.
 Figure 6: a. Cladophlebis kurtzii Frenguelli (LPPB 12618); b. Cladophlebis mesozoica Frenguelli (LPPB 12614); c. Cladophlebis mendozaensis (Geinitz) Frenguelli (LPPB 12621); d-e. Neocalamites carrerei (Zeiller) Halle (LPPB 12584 and LPPB 12581); f. aff. Nododendron suberosum Artabe et Zamuner (LPPB 12591); g-h. Neocalamostachys arrondoi Brea et Artabe (LPPB 12593 and 12565). Scale bars, e =2 cm, f, g and h= 1cm.
Figure 6: a. Cladophlebis kurtzii Frenguelli (LPPB 12618); b. Cladophlebis mesozoica Frenguelli (LPPB 12614); c. Cladophlebis mendozaensis (Geinitz) Frenguelli (LPPB 12621); d-e. Neocalamites carrerei (Zeiller) Halle (LPPB 12584 and LPPB 12581); f. aff. Nododendron suberosum Artabe et Zamuner (LPPB 12591); g-h. Neocalamostachys arrondoi Brea et Artabe (LPPB 12593 and 12565). Scale bars, e =2 cm, f, g and h= 1cm.
Fossiliferous level FL II found at locality A (Figs. 3 and 4) records another monotypic palaeocommunity dominated by sphenopsids (Brea and Artabe 1999).The sedimentary sequences in which this community is preserved consist of continental clastic units composed of black shales and mudstones resulting from deposition on the flood plains of a fluvial system. This autochtonous taphocenosis is constituted by stems, nodal diaphragms and reproductive structures assigned to Neocalamites carrerei (Zeiller) Halle 1908 (Fig. 6 d-e), aff. Nododendron suberosum Artabe and Zamuner 1991 (Fig. 6 f), and Neocalamostachys arrondoi Brea and Artabe 1999 (Fig. 6 g-h). The stem of Neocalamites and nodal diaphragm of Nododendron found in close association with reproductive Neocalamostachys were probably based on parts of individuals of a single taxon (Brea and Artabe 1999). Because plants growing in wet habitats are hygrophytes, this monotypic horsetail palaeocommunity conformed a waterdependent, bamboo-like thicket with hygrophytic adaptations associated to the flood plains of a fluvial environment (Fig. 7).
 Figure 7: Reconstruction of Triassic horsetail landscape in the flood-plain of a fluvial environment. In the background are groups of conifers (drawing by Jorge Gonzalez).
Figure 7: Reconstruction of Triassic horsetail landscape in the flood-plain of a fluvial environment. In the background are groups of conifers (drawing by Jorge Gonzalez).
COMPARISON WITH OTHERS TRIASSIC PETRIFIED FORESTS
Three other in situ Triassic petrified forests dominated by corystosperms have been discovered in Argentina, i.e., in the Cortaderita (late Middle Triassic), Ischigualasto (early Late Triassic) and Río Blanco (Late Triassic) formations (Zamuner 1992, Spalletti et al. 1999, Artabe et al. 2001, Artabe et al. 2007b). Presently, the only available structural data are from the Late Triassic corystosperm forest found at La Elcha Mine (Río Blanco Formation, Cuyo Basin). This is an evergreen and monotypic community composed of 150 stumps in life position, and the trees colonized well-drained proximal flood-plain areas, close to channel belts (Artabe et al. 2007b). The permineralized stumps were described and assigned to Elchaxylon zavattieriae (Corystospermaceae) by Artabe and Zamuner (2007). Vegetation analysis of the La Elcha forest shows that it has a clustered distribution pattern, with a forest density of 727-1504 tree /ha. The deduced height of Elchaxylon and the distribution of class diameters suggest that the canopy in the forest community would have had the majority of specimens ranging 13-21 m. Growth ring analysis indicates that the forest community colonized stressed ecosystems (Artabe et al. 2007b). At very high latitudes, in the central Transantarctic Mountains, another in situ corystosperm riparian forest was found and assigned to the Middle Triassic (Cúneo et al. 2003). The permineralized stumps were described as Jeffersonioxylon Del Fueyo et al. (1995) and later assigned to the Corystospermaceae by Cúneo et al. (2003). This monotypic forest is interpreted as deciduous and the tree density and the basal area are around 274 tree/ha and 20.83 m2/ha respectively. The Antarctic plant has been reconstructed as a tree of ~20-30 m with coniferous-like habit (Cúneo et al. 2003). Petrified remains of a Late Triassic forest were preserved in the Paraná Basin, Brazil (Pires et al. 2005). This forest was dominated by Sommerxylon spiralosus, a morphotaxon with taxacean affinities (Pires and Guerra Sommer 2004). Growth ring analysis indicates that the climate was not equable but rather with marked seasonal variations. The fossil wood shows a distinctive seasonal pattern related to growth cycles, with extensive favourable and restricted unfavourable growth periods. The external factors that affected the cycles were mainly related to cyclic restrictions of water supply and irregular changes in environmental conditions, probably linked to occasional droughts during the growing season (Pires et al. 2005). The Petrified Forest National Park in Arizona is a better known Late Triassic fossil forest in the southwestern United States (Ash and Creber 1992). This fossil forest lies within the Chinle Formation and is mainly dominated by Woodworthia arizonica and Schilderia adamanica (Creber and Ash 2004). The fossil woods of this forest do not show annual growth rings but contain irregular growth interruptions similar to those found in trees nowadays growing in humid tropics. These interruptions could be due to endogenous hormonal effects or to occasional local variations in water supply (Ash and Creber 1992).
SUMMARY
Two late Middle Triassic palaeocommunities were preserved in the Paramillo Formation, at Agua de la Zorra, northwestern Cuyo Basin, in southwestern Gondwana, i.e., the Darwin Forest associated with banks of highly sinuous fluvial systems, and second including fossil horsetails that grew on flood plains of fluvial systems. The Darwin Forest was a mixed forest, integrated by corystosperms (Cuneumxylon spallettii) and conifers (Araucarioxylon protoaraucana). Species distribution shows that in some places corystosperms and conifers are intermingled, whereas in others corystosperms and conifers appear to be aggregated into cohesive monotypic groups. The canopy is formed by two arboreal strata with a small number of emergent; the understorey with ferns. The included phloem and associated parenchyma present in Cuneumxylon spallettii may be an important adaptative strategy to avoid water stress. Thus, polyxyly as a functional anatomy characters correlates with a seasonal climatic regime. Morpho-functional analysis, structural parameters, biomass and basal stand area of the Darwin Forest allows classifying it as a mainly dry evergreen subtropical seasonal forest. The anatomy of the growth rings of Araucarioxylon protoaraucana suggests strongly seasonal conditions. Moreover, growth ring analysis indicates that this species was an evergreen gymnosperm. The second palaeocommunity integrated by sphenopsids shows herbaceous-arbustive vegetation with hygrophytic adaptations. A fluvial flood-plain in an open landscape was the environment in which this fossil vegetation developed.
ACKNOWLEDGEMENTS
The authors would like to express their thanks to Guillermo Ottone, Greg Retallack and one anonymous reviewer for their critical and constructive comments on an earlier version of this paper. We would like to thank the financial support provided by the Consejo Nacional de Investigaciones Científicas y Técnicas (Project PIP 5430) and Universidad Nacional de La Plata (11/N535). The charts of the Triassic vegetation reconstruction were made by Jorge Gonzalez.
WORKS CITED IN THE TEXT
1. Artabe, A.E., and Brea, M. 2003. A New Approach to Corystospermales based on petrified stems from the Triassic of Argentina. Alcheringa 27: 209-229.
2. Artabe, A.E., and Zamuner, A.E. 1991. Una nueva equisetal del Triásico de Cacheuta, Argentina, con estructura interna preservada. Ameghiniana 28: 287-294.
3. Artabe, A.E., and Zamuner, A.E. 2007. Elchaxylon, a new corystosperm based on permineralized stems from the late Triassic of Argentina. Alcheringa 31: 85-96.
4. Artabe, A.E., Morel, E.M. and Spalletti, L.A. 2001. Paleoecología de las floras triásicas argentinas. In Artabe, A.E., Morel, E.M. and Zamuner, A.E. (eds.) El Sistema Triásico en la Argentina. Fundación Museo de La Plata Fracisco P. Moreno: 199-225, La Plata.
5. Artabe, A.E., Morel, E.M. and Spalletti, L.A. 2003. Caracterización de las provincias fitogeográficas triásicas del Gondwana extratropical. Ameghiniana 40: 387-405.
6. Artabe, A.E., Morel, E.M and Ganuza, D.G. 2007 a. Las floras trásicas de La Argentina. In Archangelsky, S., Sánchez, T. and Tonni, E.P. (eds.) Asociación Paleontológica Argentina, Publicación Especial 11, Ameghiniana 50° aniversario: 75-86.
7. Artabe, A.E., Spalletti, L.A., Brea, M., Iglesias, A., Morel, E.M. and Ganuza, D.G. 2007 b. Structure of a corystosperm fossil forest from the Late Triassic of Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 243: 451-470.
8. Ash, S.R. and Creber, G.T. 1992. Palaeoclimatic interpretation of the wood structures of the trees in the Chinle Formation (Upper Triassic), petrified forest National Park, Arizona, USA. Palaeeogeography, Palaeoclimatology, Palaeecology 96: 299-317.
9. Avé-Lallemant, H. 1891. Estudios mineros en la provincia de Mendoza. La parte septemtrional de la Sierra de Uspallata. Academia Nacional de Ciencias, Boletín 12: 131-176, Córdoba.
10. Brea, M., 1995. Estudio de la paleoflora de la secuencia triásica de Agua de la Zorra, provincia de Mendoza. Ph. D. Thesis N° 642, Facultad de Ciencias. Naturales y Museo, Universidad Nacional de La Plata, Argentina (unpublished), 202 p.
11. Brea, M. 1997. Una nueva especie fósil del género Araucarioxylon Kraus 1870 emend. Maheshwari 1972 del Triásico de Agua de la Zorra, Uspallata, Mendoza, Argentina. Ameghiniana 34: 485-496.
12. Brea, M. 2000. Paleoflora triásica de Agua de la Zorra, Uspallata, provincia de Mendoza, Argentina: Lycophyta y Filicophyta. Ameghiniana37: 199-204.
13. Brea, M., and Artabe, A.E. 1999. Apocalamitaceae (Sphenophyta) triásicas de la Formación Paramillo, Agua de la Zorra, provincia de Mendoza, Argentina. Ameghiniana 36: 389-400.
14. Brea, M., Artabe, A.E. and Spalletti, L.A. 2005. Paleovegetation studies and growth-ring analysis of a mixed Middle Triassic forest from Argentina. Abstracts Gondwana 12: 77.
15. Brea, M., Artabe, A.E. and Spalletti, L.A. 2008. Ecological reconstruction of a mixed Middle Triassic forest from Argentina. Alcheringa 32: 365-393.
16. Brown, S. 1997. Estimating biomass and biomass change in tropical forests. FAO Forestry Paper 134, Food and Agriculture Organization of the United Nations. 49 p., Rome.
17. Burmeister, H. 1861. Reise durch die La Plata-Staaten, mit besonderer Rücksicht auf die physische Beschaffenheit und den Culturzustand der Argentinischen Republik. Ausgeführt in den Jahren 1857, 1858, 1859 und 1860. Erster Band. H.W. Schimdt, v. 1, 503 p., v. 2, 538 p., Halle.
18. Chumakov, N.M. and Zharkov, M.A. 2003. Climate during the Permian-Triassic biosphere reorganizations. Article 2. Climate of thr Late Permian and Early Triassic: General inferences. Stratigraphy and Geological Correlation 11(4): 361-375.
19. Conwentz, H. 1885. Sobre algunos árboles fósiles del Río Negro. Academia Nacional de Ciencias Boletín , 7: 435-456, Córdoba.
20. Creber, G.T. and Ash, R. 2004. The Late Triassic Schilderia adamanica and Woodworthia arizonica trees of the petrified forest National Park, Arizona, USA. Palaeontology 47: 21-38.
21. Creber, G.T. and Chaloner, W.G. 1984. Influence of environmental factors on the wood structure of living and fossil trees. Botanical Review 50: 357-448.
22. Cúneo, N.R., Taylor, E.L., Taylor, T.N. and Kring, M. 2003. In situ fossil forest from the upper Fremouw Formation (Triassic) of Antarctica: paleoenvironmental setting and paleoclimate analysis. Palaeogeography, Palaeoclimatololgy, Palaeoecology 197: 239-261.
23. Darwin, C. 1839a. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Third Volume: Journal and remarks, 1832-1836, Henry Colburn, 615 p. London.
24. Darwin, C. 1839b. Journal of researches into the geology by HMS Beagle under the command of captain FitzRoy, R.N., from 1832 to 1836, Henry Colburn, 615 p., London.
25. Darwin, C. 1845. Journal of researches into the natural history and geology of countries visited during the voyage of HMS Beagle round the world, under the command of Capt. FitzRoy, R.N. Second Edition, corrected, with additions. John Murray, 519 p., London.
26. Darwin, Ch. 1846. Geological observations on South America. Being the third part of the Geology of the voyage of the Beagle, during the years 1832 to 1836. Smith, Elder and Co. 280 p., London.
27. Del Fueyo, G., Taylor, E.L., Taylor, T.N. and Cúneo, N.R. 1995. Triassic Wood from the Gordon Valley, central transantarctic Mountains, Antarctica. IAWA Journal 16: 111-126.
28. Doering, A. 1882. Informe official de la Comisión Científica agregada al Estado Mayor General de la Expedición al Río negro (Patagonia) realizadas en los meses de Abril, Mayo y Junio de 1879, bajo las órdenes del General D. Julio A. Roca. Tercera Parte. Geología. Ostwald & Martinez, 299-530, Buenos Aires.
29. Dubiel, R.F., Parrish, J.T., Parrish, J.M. and Good, S.C. 1991. The Pagaean megamonsoon - evidence from the Upper Triassic Chinle Formation, Colorado Plateau. Palaios 6: 347-370.
30. Du Toit, A.L. 1927. A geological comparison of South America with South Africa. Carnegie Institution Publication 381, 158 p., Washington.
31. Falcon-Lang, H.J. 2000a. A method to distinguish between woods produced by evergreen and deciduous coniferopsids on the basis of growth ring anatomy: a new palaeoecological tool. Palaeontology 43: 785-793.
32. Falcon-Lang, H.J. 2000b. The relationship between leaf longevity and growth ring markedness in modern conifer woods and its implications for palaeoclimatic studies. Palaeogeogrphy, Palaeoclimatololgy, Palaeoecololy 160: 317- 328.
33. Fahn, A. 1990. Plant anatomy. Pergamon Press. 588 p., Oxford.
34. Frenguelli, J. 1947. El género Cladophlebis y sus representantes en Argentina. Gaea 7: 159-369.
35. Fritts, H.C. 1976. Tree rings and climate. Academic Press. 567 p., London.
36. Groeber, P. 1939. Mapa geológico de Mendoza. Physis 14: 171-220.
37. Halle, T.G. 1908. Zur Kenntnis der Mesozoischen equisetales Schwedens. Kungliga. Svenska vetenskaps akademie Handlingar 43: 3-42.
38. Harrington, H.J. 1971. Descripción geológica de la hoja 22c, "Ramplón" provincias de Mendoza y San Juan. Carta Geológica-Económica de la República Argentina Escala 1:200.000. Ministerio de Industria y Minería, Subsecretaria de Minería, Dirección de Geología y Minería, Buenos Aires, Boletín 114: 87 p.
39. Holmes, R.L. 1985. Computer-assisted quality control in tree-ring dating and measurement. A users manual for program COFECHA. Laboratory of tree-ring Research: 2-23.
40. Kokogian, D., and Mancilla, O. 1989. Análisis estratigráfico y secuencial de la Cuenca Cuyana. In Chebli, G. and Spalletti, L. (eds.) Cuencas Sedimentarias Argentinas. Universidad Nacional de Tucumán, Serie Correlación Geológica 6: 169-201.
41. Kokogián, D., Fernández Seveso, F., and Mosquera, A. 1993. Las secuencias sedimentarias triásicas. In Ramos, V.A. (ed.), Relatorio Geología y Recursos Naturales de Mendoza. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos 1(7): 65-78.
42. Kurtz, F. 1921. Atlas de plantas fósiles de la República Argentina. Academia Nacional de Ciencias, Actas 7 : 129-153, Córdoba.
43. Linares, E., and González, R. 1990. Catálogo de edades radimétricas de la República Argentina 1957-1987. Asociación Geológica Argentina, Serie B (Didáctica y Complementaria) 19: 628 p.
44. Massabie, A.C. 1985. Filón capa Paramillo de Uspallata, su caracterización geológica y edad, Paramillo de Uspallata, Mendoza. Primeras Jornadas sobre Geología de Precordillera, Actas 1: 325-330.
45. Massabie, A.C., Rapalini, A.E. and Soto, J.L. 1985. Estratigrafía del Cerro Los Colorados, Paramillo de Uspallata, Mendoza. Primeras Jornadas sobre Geología de Precordillera, Actas 1: 71-76.
46. Morel, E.M., Artabe, A.E., and Spalletti, L.A. 2003. The Triassic floras of Argentina: Biostratigraphy, Floristic events and comparison with other areas of Gondwana and Laurasia. Alcheringa 27: 231-243.
47. Murphy, P.G. and Lugo, A.E. 1986. Structure and biomass of a subtropical dry forest in Puerto Rico. Biotropica 18: 89-96.
48. Ottone, E.G. 2005. The history of palaeobotany in Argentina during the 19th century. In Bowden, A.J., Burek, C.V. and Wilding, R. (eds.) History of Paleobotany: Selected Essays. Geological Society, Special Publications 241: 281-294.
49. Parrish, J.T. 1993. Climate of the supercontinent Pangea. Journal of Geology 101: 215-233.
50. Parrish, J.T., Ziegler, A.M., and Scotese, C.R. 1982. Rainfall patterns and the distribution of coals and evaporates in the Mesozoic and Cenozoic. Palaeogeography, Palaeoclimatology, Palaeoecology 40: 67-101.
51. Pires, E.T. and Guerra Sommer, M. 2004. Sommerexylon spiralosus from Upper Triassic in southernmost Paraná Basin (Brazil): a new taxon with taxacean affinity. Anais da Academia Brasileira de Ciências 76: 595-609.
52. Pires, E.T., Guerra Sommer, M. and Dos Santos Scherer, C.M. 2005. Late Triassic climate in southernmost Parana Basin (Brazil): evidence from dendrochronological data. Journal of South American Earth Sciences 18: 213-221.
53. Poma, S., Quenardelle, S. and Koukharsky, M. 2004. Un flujo piroclástico del Triásico en la Formación Paramillo, provincia de Mendoza. 7º Congreso Argentino de Mineralogía y Metalogenia: 385-390.
54. Ramos, V.A. 1993. El magmatismo Triásico-Jurásico de intraplaca. In Ramos, V.A. (ed.) Relatorio Geología y Recursos Naturales de Mendoza. 12 Congreso Geológico Argentino y 2 Congreso de Exploración de Hidrocarburos 1(8): 79-86.
55. Ramos, V.A., and Kay, S.M. 1991. Triassic rifting and associated basalts in the Cuyo Basin, central Argentina. In Harmon, R.S. and Rapela, C.W. (eds.) Andean magmatism and its tectonic setting. Geological Society of America, Boulder, Special Paper 265: 79-91.
56. Robinson, P.L. 1973. Palaeoclimatology and continental Drift. Implications of Continental Drift in the Earth Sciences 46: 451-476.
57. Rusconi, C. 1941. "Bosques petrificados" de Mendoza. Anales de la Sociedad Científica Argentina 132: 80-95.
58. Sellwood, B.W., and Valdes, P.J. Mesozoic climates: General circulation models and the rock record. Sedimentary Geology 190: 269-287.
59. Scotese, C.R., Boucot, A.J. and McKerrow, W.S. 1999. Gondwanan palaeogeography and palaeoclimatology. Journal of African Earth Sciences 28: 99-114.
60. Spalletti, L.A., Artabe, A.E. and Morel, E.M. 2003. Geological factors and evolution of Southwestern Gondwana Triassic Plants. Gondwana Research 6: 119-134.
61. Spalletti, L.A., Artabe, A.E., Morel, E.M. and Brea, M. 1999. Biozonación paleoflorística y cronoestratigrafía del Triásico Argentino. Ameghiniana 36: 419 -451.
62. Stappenbeck, R. 1910. La Precordillera de San Juan y Mendoza. Anales del Ministerio de Agricultura, sección Geología, Mineralogía y Minería, 4 (3): 3-183, Buenos Aires.
63. Stelzner, A. 1885. Beiträge zur Geologie der Argentinischen Republik und des Angrenzender, zwischen dem 32. und 33°. S. gelegenen Theiles der chilenischen Cordillere. In Stelzner, A. (ed.) Beiträge zur Geologie und Palaeontologie der Argentinschen Republik, 1. Geologischer Theil. Palaeontograhica, Supplementbände 3, 29: 1-329.
64. Stewart, W.N. 1983. Paleobotany and the evolution of plants. Cambridge University Press. 405 p.
65. Stipanicic, P.N. 2001. Antecedentes Geológicos y Paleontológicos. In Artabe, A.E., Morel, E.M. and Zamuner, A.B. (eds.) El Sistema Triásico en la Argentina. Fundación Museo de La Plata Francisco Pascasio Moreno: 1-21, La Plata.
66. Stipanicic, P.N. and Marsicano, C.A. 2002. Léxico estratigráfico de la Argentina, Volumen VIII: Triásico. Asociación Geológica Argentina, Serie B (Didáctica y Complementaria) 26, 370 p.
67. Stipanicic, P.N., Herbst, R. and Bonetti, M.I.R. 1996. Floras Triásicas. Academia Nacional de Ciencias, Actas 11: 127-184, Córdoba.
68. Strelkov, E.E., and Álvarez, L.A. 1984. Análisis estratigráfico y evolutivo de la Cuenca Triásica Mendocina-Sanjuanina. 9° Congreso Geológico Argentino (Bariloche), Actas 3: 115-130.
69. Willis, K.J. and McElwain, J.C. 2002. The evolution of plants. Oxford University Press, 378 p.
70. Windhausen, H. 1941. El "Rético" del Paramillo de Uspallata (Mendoza), PhD Thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, (unpublished), 28, 90 p., La Plata.
71. Zamuner, A.B. 1992. Estudio de una tafoflora de la localidad tipo, de la Formación Ischigualasto (Neotrias), provincia de San Juan. Ph.D. Thesis, Facultad de Ciencias Naurales y Museo, Universidad Nacional de La Plata, (unpublished), 583, 97 p., La Plata
72. Zamuner, A.B., Zavattieri, A.M., Artabe, A.E. and Morel, E.M. 2001. Paleobotánica. In Artabe, A.E., Morel, E.M. and Zamuner, A.B. (eds.) El Sistema Triásico en la Argentina. Fundación Museo de La Plata Francisco P. Moreno : 143-184. La Plata.
Beatriz Aguirre-Urreta and Verónica Vennari
Laboratorio de Bioestratigrafía de Alta Resolución, Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Ciudad Universitaria, Buenos Aires. Email: aguirre@gl.fcen.uba.ar
ABSTRACT
The aim of this work is to summarize the modern knowledge of the geology of the Piuquenes Pass, in the Main Andes of Argentina and Chile, and to describe a small fauna of Tithonian-Neocomian invertebrates mostly represented by ammonites. The present knowledge of the region is compared with Darwin's, as expressed in his famous book on the Geology of South America.
Keywords: Main Andes; Mesozoic; Ammonites; Piuquenes Pass; Darwin.
RESUMEN: Tras las huellas de Darwin a través de los Andes: invertebrados fósiles del Tithoniano-Neocomiano del paso de Piuquenes. El objetivo de este trabajo es presentar una breve revisión moderna de la geología del Paso de Piuquenes en los Andes Principales de Chile y Argentina y describir una pequeña fauna de invertebrados del Tithoniano-Neocomiano compuesta principalmente por amonites. Se compara también el conocimiento de esta región con la referida por Darwin en su famoso libro sobre la geología de América del Sur.
Palabras clave: Andes Principales; Mesozoico; Amonites; Paso de Piuquenes; Darwin.
INTRODUCTION
During his 5 years' voyage around the world on board the HMS Beagle, Darwin spent more than 3 years on shore. This was most probably due to his interest in exploring new regions, but also to avoid the strong sea-sickness that annoyed him when sailing. During the Beagle years and some time later Darwin was foremost a geologist, though we are used to think of him as a pure biologist. Thus, his enormous collection of fauna and flora came second to his interest as can be seen in the fact that he gave most of it to different specialists after his return to England. In March-April 1835, Darwin undertook a long ride to examine the geology of the Andes. He started the journey in Valparaiso on the Pacific coast, crossed the Andes along the Piuquenes and Portillo passes, reached Mendoza city and went back to Chile by the Cumbre or Uspallata Pass (Fig. 1). This trip was probably one of the most valuable to him from a scientific point of view as can be seen in the long and detailed letters he sent both to Professor Henslow and to his sister Susan. The letter to Henslow begins thus: "I have just returned from Mendoza, having crossed the Cordilleras by two passes. This trip has added much to my knowledge of the geology of the country. Some of the facts, of the truth of which I in my own mind feel fully convinced, will appear to you quite absurd& incredible", followed by several paragraphs devoted to the structure of the cordillera (Burkhardt and Smith 1985).
 Figure 1: Location of the Piuquenes Pass in the Main Andes of Argentina and Chile.
Figure 1: Location of the Piuquenes Pass in the Main Andes of Argentina and Chile.
One hundred and sixty years later, in the summer of 1995 one of us (BAU) followed the steps of Darwin along the Piuquenes Pass, checking the stratigraphy and collecting fossils. Thus, the objective of this work is to summarize the modern knowledge of the geology and palaeontology of the region of the Piuquenes Pass, in the Main Andes of Argentina and Chile, comparing these views with Darwin's (1846), as expressed in his famous book on the Geology of South America.
REGIONAL GEOLOGICAL SETTING
The Piuquenes (Darwin always spelled it as Peuquenes) Pass runs across the boundary between Argentina and Chile in the Main Central Andes (33°38'S, 69°52'W, Fig. 1). It reaches 4,030 m a.s.l., and though it is the international divide, the highest pass is Portillo, located some 24 km to the east, with 4,374 m a.s.l. The geology of this part of the Main Andes was studied in detail by Polanski (1964) as part of the mapping program of the Geological Survey of Argentina. It is characterized by a normal subduction zone, where the oceanic Nazca plate subducts beneath South America at an angle of 30° (Jordan et al. 1983). As a result of that the Mesozoic sequence is heavily deformed in a series of thrust slices (see Giambiagi et al. 2009). These thrusts produced an imbricated sequence, where the strata are repeated several times, creating a complex structure. The basement of the Andes at this latitude is composed of metamorphic rocks of Precambrian age. Carboniferous marine successions devoid of fossils and some rhyolitic tuffs and lavas of Permian and Triassic age are unconformably overlying the basement (Polanski 1964). The Mesozoic sequences, which record a Pacific marine transgression, constitute different stratigraphic cycles described by Groeber (1953), Malumián and Ramos (1984) and Riccardi (1983, 1988). These sedimentary successions are covered by extensive Cenozoic volcanic and pyroclastic rocks, including some Recent volcanic centers, such as San José and Marmolejo volcanoes. Isolated patches of colluvial and alluvial deposits are widely distributed in the region (Fig. 2).
 Figure 2: Geological map of the region of the Piuquenes Pass, Main Andes (from Pángaro et al. 1996).
Figure 2: Geological map of the region of the Piuquenes Pass, Main Andes (from Pángaro et al. 1996).
THE GEOLOGY OF THE PIUQUENES PASS
A geological map of the Piuquenes Pass region is shown in figure 2 (Pángaro et al. 1996), where Darwin's itinerary is marked, and the rock succession according to Darwin (1846) is also shown. He described the following: "The ridge of Peuquenes, which divides the waters flowing into the Pacific and Atlantic oceans, extends in a nearly N.N.W. and S.S.E. line; its strata dip eastward at an angle of between 30° and 45°, but in the higher peaks bending up and becoming almost vertical. Where the road crosses this range, the height is 13,210 feet above the sea-level, and I estimated the neighbouring pinnacles at from 14,000 to 15,000 feet. The lowest stratum visible in this ridge is a red stratified sandstone [P]; on it are superimposed two great masses [Q and S] of black, hard, compact, even having a conchoidal fracture, calcareous, more or less laminated shale, passing into limestone: this rock contains organic remains, presently to be enumerated. The compacter varieties fuse easily in a white glass; and this I may add is a very general character with all the sedimentary beds in the Cordillera: although this rock when broken is generally quite black, it everywhere weathers into an ash-grey tint. Between these two great masses [Q and S], a bed [R] of gypsum is interposed, about 300 feet in thickness, and having the same characters as heretofore described. I estimated the total thickness of these three beds [Q, R, S] at nearly 3000 feet; and to this must be added, as will be immediately seen, a great overlying mass of red sandstone. In descending the eastern slope of this great central range, the strata, which in the upper part dip eastward at about an angle of 40°, become more and more curved, till they are nearly vertical; and a little further onwards there is seen on the further side of a ravine, a thick mass of strata of bright red sandstone [T], with their upper extremities slightly curved, showing that they were once conformably prolonged over the beds [S]: on the southern and opposite side of the road, this red sandstone and the underlying black shaly rocks stand vertical, and in actual juxtaposition" (Darwin 1846, p. 179-180, pl. 1, sketch 1, see Giambiagi et al. 2009). The red sandstones of P correspond to the Río Damas Formation in Chile (Tordillo Formation in Argentina). Q and S of Darwin (1846) are the Lo Valdés Formation in Chile or the Mendoza Group in Argentina, where it is composed by the Vaca Muerta, Chachao and Agrio Formations. R, the gypsum bed, is the Auquilco Formation of Argentina (Río Colina Formation in Chile) of Oxfordian-Kimmeridgian age, and T is again the Tordillo Formation. It is obvious now that Darwin did not quite understand the complex structure of the region, as can be seen in the geological map of figure 2, where a western backthrust puts the Auquilco Formation on top of the Agrio Formation and a second fore-thrust repeats again the Agrio and Chachao Formations (Pángaro et al. 1996, Giambiagi et al. 2009). A section was measured on the Argentinean slope (Fig. 3), along the northern bank of the Arroyo Piuquenes (Aguirre-Urreta 1996; see location in Fig. 2). The top beds of the Tordillo Formation (Kimmeridgian) are sandstones interbedded with siltstones and green shales. The Vaca Muerta Formation begins with a few meters of limestone with algal lamination, followed in its lower part by black papyraceous shales with abundant calcareous nodules with three fossiliferous levels, from bottom to top with: Virgatosphinctes aff. V. denseplicatus rotundus Spath, 1931, Pseudolissoceras zitteli (Burckhardt, 1903) and Aulacosphinctes proximus (Steuer, 1897). These shales grade upwards to dark grey, thinly laminated limestones, reaching more than 110 metres (Fig. 3). The age of the Vaca Muerta Formation in the area is Early to Middle-Late? Tithonian. The Chachao Formation - 40 metres thick - is composed by massive oyster coquinas interbedded with shales bearing Parodontoceras calistoides (Behrendsen, 1891) of Late Tithonian age in its lower part. The Agrio Formation is some 120 metres thick in the measured section but is incomplete, as it is cut by a thrust that puts the Chachao coquinas on top of its upper part. The lower part is mostly composed by intercalations of massive and finely laminated grey and yellowish limestones, bearing - in some levels- Steinmanella sp., Thalassinoides isp. and poorly preserved flattened ammonoids. In the upper part there are non-fossiliferous olive green shales, limestone with small oysters, silty limestones and black shales with Spitidiscus riccardii Leanza and Wiedmann, 1992 (Aguirre-Urreta 2001), Protohemichenopus neuquensis Camacho, 1953, Lucinidea indet., and callianassid crustaceans.
 Figure 3: Stratigraphic section of the Mendoza Group in the Argentine slope of the Piuquenes Pass.
Figure 3: Stratigraphic section of the Mendoza Group in the Argentine slope of the Piuquenes Pass.
The fossils that Darwin collected in Q and S (presently Lo Valdés Formation/ Mendoza Group) were studied by d'Orbigny and cited by Darwin (1846, p. 181) as follows: "The fossils above alluded to in the black calcareous shales are few in number, and are in an imperfect condition; they consist, as named for me by M. d'Orbigny, of
1. Ammonite, indeterminable, near to A. recticostatus, D'Orbig. Pal. Franc. (Neocomian formation). 2. Gryphæa, near to G. Couloni, (Neocomian formations of France and Neufchatel). 3. Natica, indeterminable. 4. Cyprina rostrata, D'Orbig. Pal. Franc. (Neocomian formation). 5. Rostellaria angulosa (?), D'Orbig. Pal. de l'Amer. Mer. 6. Terebratula ?"
However, it seems that d'Orbigny gave Darwin slightly different information (see fig. 4). According to his notes, the specimens collected by Darwin in the "Cordillera Centrale du Chili" comprised: "617/792. Gryphaea - Voisine du Gryphaea Couloni, du terrain Neocomien de France et de Neuchatel 613. Natica (indeterminable) 619. Ammonite indeterminable, voisine de l'A. recticostatus, d'Orb (Paleont. française) Du terrain Neocomien 614. B. Cyprina rostrata, d'Orb. Paleont. franç. Terrain Neocomien (non Lucina) C. Terebratula ? A. Rostellaria angulosa, d'Orb. Paleont. de l'Am. Mer. Pl. 18 fig. 4 ?"
 Figure 4 : List of fossils identified by d'Orbigny for Darwin from his collection of the Central Cordillera of Chile (from www.darwin-online.org.uk).
Figure 4 : List of fossils identified by d'Orbigny for Darwin from his collection of the Central Cordillera of Chile (from www.darwin-online.org.uk).
d'Orbigny also listed : "790. Arca peut être Arca gabrielis, d' Orb. (Paleontologie francaise) du terrain Neocomien" This fossil is not mentioned by Darwin from the Piuquenes Pass, and is almost certainly from Puente del Inca (Darwin 1846, p. 193), as this specimen and the "792 (Gryphaea - Voisine du Gryphaea Couloni)" have a note in the left margin stating Cumbre, probably handwritten by Darwin (Fig. 4). D'Orbigny assigned a bivalve to Cyprina rostrata d'Orbigny, 1853 pointing out that this is not Lucina Lamarck, 1799, which is interesting as here this fossil is referred to Lucinidae indet. (D. Lazo, oral comm.) (Fig. 5, p-q). He also added the figure of Rostellaria angulosa d'Orbigny, 1842 (d'Orbigny 1842, p. 80, pl. 18, fig. 4) which was very helpful in the assignation of our specimens to Protohemichenopus neuquensis Camacho, 1953 (Fig. 5 o). It is worth mentioning here that, besides the ammonites that will be described in detail below, we have also found chelipeds of callianassid crustaceans in both the Vaca Muerta and Agrio Formations. Besides Darwin (1846) the other only published mention of fossils in the Piuquenes range is that of Polanski (1964, p. 48) who listed the following Tithonian fossils: Corongoceras sp., Aulacosphinctes sp., Trigonia carricurensis Leanza, Lucina sp. and Exogyra sp.
 Figure 5: a-h) Pseudolissoceras zitteli (Burckhardt, 1903); a) lateral view CPBA 20552.1; b) lateral and ventral views CPBA 20552.5; c) lateral and ventral views CPBA 20552.4; d) lateral view CPBA 20552.3; e) lateral view CPBA 20552.2; f) lateral view CPBA 20552.6; g) apertural and lateral views CPBA 20552.7; h) lateral and ventral views CPBA 20552.8; i-m) Virgatosphinctes aff. Virgatosphinctes denseplicatus rotundus Spath, 1931; i) lateral view CPBA 20551.3; j) lateral view CPBA 20551.4; k) ventral and lateral views CPBA 20551.2; l) lateral view CPBA 20551.1; m) apertural and lateral views CPBA 20551.5; n) Spitidiscus riccardii Leanza and Wiedmann, 1992, lateral view CPBA 20557; o) Protohemichenopus neuquensis Camacho, 1953, apertural and lateral views CPBA 20556.1; p-q) Lucinidae indet. lateral views CPBA 20555.1-2; r-t) Aulacosphinctes proximus (Steuer, 1897); r) lateral view CPBA 20553.2; s) lateral and ventral views CPBA 20553.1; t) Parodontoceras calistoides (Behrendsen, 1891), lateral view CPBA 20554. All specimens from the Piuquenes Pass, Mendoza. All natural size. Specimens coated with ammonium chloride.
Figure 5: a-h) Pseudolissoceras zitteli (Burckhardt, 1903); a) lateral view CPBA 20552.1; b) lateral and ventral views CPBA 20552.5; c) lateral and ventral views CPBA 20552.4; d) lateral view CPBA 20552.3; e) lateral view CPBA 20552.2; f) lateral view CPBA 20552.6; g) apertural and lateral views CPBA 20552.7; h) lateral and ventral views CPBA 20552.8; i-m) Virgatosphinctes aff. Virgatosphinctes denseplicatus rotundus Spath, 1931; i) lateral view CPBA 20551.3; j) lateral view CPBA 20551.4; k) ventral and lateral views CPBA 20551.2; l) lateral view CPBA 20551.1; m) apertural and lateral views CPBA 20551.5; n) Spitidiscus riccardii Leanza and Wiedmann, 1992, lateral view CPBA 20557; o) Protohemichenopus neuquensis Camacho, 1953, apertural and lateral views CPBA 20556.1; p-q) Lucinidae indet. lateral views CPBA 20555.1-2; r-t) Aulacosphinctes proximus (Steuer, 1897); r) lateral view CPBA 20553.2; s) lateral and ventral views CPBA 20553.1; t) Parodontoceras calistoides (Behrendsen, 1891), lateral view CPBA 20554. All specimens from the Piuquenes Pass, Mendoza. All natural size. Specimens coated with ammonium chloride.
SYSTEMATIC PALAEONTOLOGY
Family Haploceratidae Zittel, 1884 Genus Pseudolissoceras Spath, 1925
Pseudolissoceras zitteli (Burckhardt, 1903) Figs. 5 a-h
? 1897 Oppelia perlaevis Steuer, p. 73, pl. 6, figs. 7-9 1900 Oppelia aff. perlaevis Steuer; Burckhardt, p. 46, pl. 26, figs. 5, 6; pl. 29, fig. 2 1903 Neumayria zitteli Burckhardt, p. 55, pl. 10, figs. 1-8 1907 Neumayria zitteli Burckhardt; Haupt, p. 200, pl. 7, figs. 3a, b, 4a-c ? 1921 Oppelia perlaevis Steuer, p. 102, pl. 6, figs. 7-9 ? 1921 Oppelia perglabra Steuer, p. 104, pl. 7, figs. 13-15 1925 Pseudolissoceras zitteli (Burckhardt); Spath, p. 113 (Gen. nov.) 1926 Pseudolissoceras zitteli Burckhardt; Krantz, p. 436, pl. 17, figs. 4, 5 1928 Pseudolissoceras zitteli Burckhardt; Krantz, p. 18, pl. 1, fig. 6; pl. 4, figs. 9a, b 1931 Pseudolissoceras zitteli Burckhardt; Weaver, p. 401, pl. 43, fig. 291 1942 Pseudolissoceras cf. Pseudolissoceras zitteli (Burckhardt); Imlay, p. 1443, pl. 4, figs. 1-4, 7, 8, 11, 12 1980 Pseudolissoceras zitteli (Burckhardt); Leanza, p. 17, pl. 1, figs. 1a, b, 2a, b 2001 Pseudolissoceras zitteli (Burckhardt); Parent, p. 23, figs. 3a, b, 5a, b, 7a-f
Material: 10 specimens (CPBA 20552.1- 20552.10) from the Piuquenes Pass, Mendoza.
Description: Well preserved, small to medium size specimens (largest specimen is 62.2 mm diameter); body chamber occupies at least half a whorl, usually laterally crushed. Shell discoidal, involute (U c. 21% of diameter) with deep umbilicus, rounded umbilical border and an abrupt umbilical slope on the outermost whorls while on the innermost it becomes gently inclined. Subelliptic transversal section with convex flanks converging on a rounded and slightly acuminated venter. Whorls always higher than wide, with maximum width near the middle of the flanks. Shell surface almost smooth. Only where test is preserved is it possible to distinguish extremely soft, falcoid ribs densely packed, that start at the base of the umbilical slope. No dimorphic sexual structures observed. Suture line not completely preserved.
Dimensions of specimens
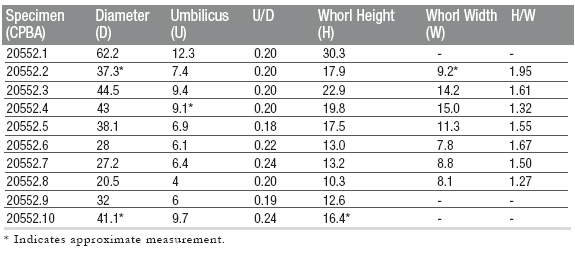
Remarks: Specimens described above can certainly be assigned to Pseudolissoceras zitteli (Burckhardt 1903, p. 55, pl. 10, figs. 1-8), and it is also possible that specimens figured as Oppelia perlaevis Steuer (1897, p. 73, pl. 6, figs. 7-9; 1921, Spanish translation of 1897 edition, p. 102, pl. 6, figs. 7-9) and Oppelia perglabra Steuer (1921, p. 104, pl. 7, figs. 13-15) from the lowermost Middle Tithonian of Mendoza, belong also in the same species. This is due to the fact that there is a strong similarity in general form, ornamentation, and suture line between them and Burckhardt's original form. Nevertheless, we can not confirm this until we have the chance of comparing our specimens with Steuer's holotypes, although we can confidently assert that they do not belong in Oppelia Waagen, 1869 because they show no trace of even a feeble keel preserved on the illustrated material. Specimens described and figured by Haupt (1907, p. 200, pl. 7, fig. 3a, b, 4ac), Krantz (1926, p. 436, pl. 17, figs. 4, 5; 1928, p. 18, pl. 1, fig. 6; pl. 4, fig. 9a, b), Weaver (1931, p. 401, pl. 43, fig. 291) and Leanza (1980, p. 17, pl. 1, figs. 1a, b, 2a, b), were all found at Cerro Lotena, Neuquén, although some specimens have also been recovered from Picún Leufú (Weaver), Rodeo Viejo, Bardas Blancas, Arroyo Cienaguitas and from a locality between Cajón del Burro and Río Choica, in Mendoza (Krantz 1926). Specimens studied by Parent (2001, p. 23, fig. 3a, b, 5a, b, 7a-f) are from Cañadón de los Alazanes, a locality exposed at the Vaca Muerta Range in Neuquén, while Imlay's material (1942, p. 1443, pl. 4, figs. 1-4, 7, 8, 11, 12) is from Cuba. In all cases specimens are highly similar; there is a notable concordance in U/D and H/W ratios with only some minimum differences in transversal section -which can be sometimes more inflated or depressed- or in having a more or less acuminate or rounded venter. In all cases, specimens are found in association with a similar fauna composed mainly of other haploceratid ammonoids, from which they can be generally differentiated by the absence of a marked groove on the flanks, as occurs in Hildoglochiceras Spath, 1924, or its rounded venter and lesser degree of involution than in Parastreblites Donze and Enay, 1961. On the other hand, many authors (Haupt 1907, Krantz 1926, Weaver 1931, Leanza 1980) distinguish Pseudolissoceras pseudoolithicum (Haupt, 1907) recorded in the same beds that contain Pseudolissoceras zitteli. That species never exhibits an umbilical edge, it is only moderately involute, and its flanks are more inflated with whorls as high as wide in transverse section.
Ocurrence: Lowermost Middle Tithonian. Pseudolissoceras zitteli Zone.
Family Perisphinctidae Steinmann, 1890 Subfamily Virgatosphinctinae Spath, 1923 a Genus Virgatosphinctes Uhlig, 1910
Virgatosphinctes aff. Virgatosphinctes denseplicatus rotundus Spath, 1931 Figs. 5 i-m
? 1890 Perisphinctes contiguus Catullo; Toucas, p. 581, pl. 14, fig. 4 ? 1900 Perisphinctes contiguus Catullo; Burckhardt, p. 45, pl. 24, fig. 1 1903 Perisphinctes contiguus Catullo; Burckhardt, p. 38, pl. 4, figs., 7-10 ? 1931 Virgatosphinctes denseplicatus (waagen) var. rotunda Spath, p. 532, pl. 96, figs. 3a-b; pl. 102, fig 4. 1954 Virgatosphinctes cf. denseplicatus (Waagen) var. rotunda Spath; Indans, p. 106, pl. 21, fig. 1 1972 Virgatosphinctes denseplicatus Waagen; Fatmi, p. 346, pl. 8, figs. 5a, b 1980 Virgatosphinctes denseplicatus rotundus Spath; Leanza, p. 31, pl. 2, figs. 2, 3
Material: 6 specimens (CPBA 20551.1-20551.6) from the Piuquenes Pass, Mendoza.
Description: Medium size discoidal phragmacone (largest specimen is 45.9 mm in diameter), body chamber not preserved. Slightly involute (U c. 33% of diameter), with moderately deep umbilicus and umbilical border not very inclined. Venter slightly planar with gently curved flanks. Whorl section somewhat inflated, slightly wider than high, with maximum width at umbilical margin. Fine, sharp ribs, regularly distributed, beginning at the middle of the umbilical slope, with a slightly prorsiradiate tendency and then bifurcate at the external third of the flank, crossing the venter without interruption. A slight weakness occurs in the ribs in specimens that are less than 38 mm in diameter. At least two shallow constrictions are observed in the last whorl preserved, parallel to the rib pattern and bordered posteriorly by a simple asymmetrical rib. Suture line not observed.
Dimensions of specimens

Remarks: Specific assignation of the specimens to Virgatosphinctes denseplicatus rotundus Spath (1931, p. 532, pl. 96, figs. 3a-b; pl. 102, fig. 4) is quite difficult due to the fact that ornamentation suffers a radical change in the last whorl, just as illustrated by Waagen in Virgatosphinctes denseplicatus (1873, p. 201, pl. 46, fig. 3a, b; pl.55, figs. 1a. b), where the innermost fine ribs become stronger over the inner third of the last whorl, and then differentiate in the middle of the flank into a bundle of 4 to 10 secondary ribs that cross the venter without any interruption. It is important to state here that the gradual differentiation from biplicate ribs, into triplicate, virgatotome, and finally fasciculate ribs that constitutes a diagnostic character of Virgatosphinctes Uhlig, 1910 is not evident in Waagen's illustrated specimens from the Lower Tithonian of Kutch. Perisphinctes contiguus Catullo (in Toucas 1890, p. 581, pl. 14, fig. 4, Burckhardt 1900, p. 45, pl. 24, fig. 1, Burckhardt 1903, p. 38, pl. 4, figs. 7-10) closely resembles the specimens described above, with similar rib pattern and alternation of triplicate ribs on one side with biplicate ribs on the other; there is also a slight attenuation of the ornamentation on the venter of some specimens. Toucas' material comes from the Lower Tithonian of the Carpathians, the Alps, the Apennines and from Andalucia, while Burkhardt's specimens are from the Lower Tithonian from Casa Pincheira in Mendoza. Virgatosphinctes cf. denseplicatus (Waagen) var. rotunda Spath (in Indans 1954, p. 106, pl. 21, fig. 1) from Cerro Alto in southern Mendoza also resembles the studied specimens, as well as the specimen from western Pakistan identified as Virgatosphinctes denseplicatus Waagen by Fatmi (1972, p. 346, pl. 8, figs. 5a, b). However, there appears to be a difference in the ornamentation pattern with the Argentinian specimens, as there are simple posterior ribs bordering the constrictions, not joining the immediately anterior biplicate ribs. Finally, there is a strong resemblance between the specimens studied here and Virgatosphinctes denseplicatus rotundus described and figured by Leanza from the Lower Tithonian of Cerro Lotena, Neuquén (1980, p. 31, pl. 2, figs. 2, 3), though his specimens show a more inclined umbilical slope and a more subelliptical whorl section. Virgatosphinctes denseplicatus rotundus is usually found in association with other Virgatosphinctes, like Virgatosphinctes andesensis (Burckhardt), from which it can be differentiated due to its whorl section only slightly wider than high, its softer ornamentation and the lack of intercalatory ribs and primary ribs describing an inflection on the middle of the flanks.
Ocurrence: Basal section of Vaca Muerta Formation, just above the biolaminated carbonate levels. Lower Tithonian, Virgatosphinctes mendozanus Zone.
Family Perisphinctidae Steinmann, 1890 Subfamily Himalayitinae Spath, 1925 Genus Aulacosphinctes Uhlig, 1910
Aulacosphinctes proximus (Steuer, 1897) Figs. 5 r-t
1897 Reineckeia proxima Steuer, p. 34, pl. 8, figs. 7-11 ? 1897 Perisphinctes colubrinus Reinecke; Steuer p. 62, pl. 15, fig. 11 1900 Perisphinctes colubrinus Reinecke; Burckhardt, p. 44, pl. 24, figs. 5, 6; pl. 26, fig. 4 1907 Perisphinctes proximus Steuer; Haupt, p. 192 1921 Reineckeia proxima Steuer, p. 61, pl. 8, figs. 7-11 ? 1921 Perisphinctes colubrinus Reinecke; Steuer, p. 90, pl. 15, fig. 11 1928 Aulacosphinctes wanneri Krantz, p. 42, pl. 2, figs. 6a, b ? 1931 Aulacosphinctes proximus (Steuer); Weaver, p. 411, pl. 44, figs. 298, 299 1931 Aulacosphinctes colubrinus (Reinecke); Weaver, p. 413, pl. 44, figs. 301-303 1980 Aulacosphinctes proximus (Steuer); Leanza, p. 44, pl. 6, figs. 2a, b, 4a, b, 5a, b 1981 Aulacosphinctes proximus (Steuer); Leanza, p. 587, pl. 2, figs. 9, 10
Material: 2 specimens (CPBA 20553.1- 20553.2) from the Piuquenes Pass, Mendoza.
Description: Medium size planulate shell (largest specimen is 52.3 mm in diameter), body chamber partially preserved, of at least 1/4 whorl. Strongly evolute (U c. 45% of diameter), with shallow umbilicus, rounded umbilical border and gradually inclined umbilical slope. Subquadrate transversal section with slightly convex flanks and flat venter, whorls slightly higher than wide. Prominent, fine, sharp and distant ribs start at the base of the umbilical slope in a rursiradiated mode, at the umbilical margin they become prorsiradiated, crossing the flanks in a prorsiradiated to rectiradiated pattern. Number of ribs decreases as diameter increases; in a 52.3 mm diameter specimen there are 25 ribs in the last whorl and 20 in the preceding one. Ribs on the innermost whorls are interrupted over the venter by a narrow and shallow furrow, which tends to disappear in outer whorls although ornamentation there may suffer an important weakening. Simple primary ribs are irregularly intercalated with biplicate ribs that bifurcate at the middle of the flanks without developing any tubercle at the bifurcation point. Some intercalatory ribs are also observed, not extending beyond the middle part of the flanks. Two wide but shallow constrictions occur near the shell end; they are posteriorly bounded by a simple, acute and prominent primary rib. Suture line not visible.
Dimensions of specimens

Remarks: Diagnostic characters observed in the specimens studied, such as evolution degree, rib pattern and the presence of a ventral furrow allow its assignation to Aulacosphinctes proximus (Steuer 1897, p. 34, pl. 8, figs. 7-11 as Reineckeia proxima; 1921 Spanish translation, p. 61, pl. 8, figs. 7-11). However, in the same publication Steuer describes another species, from Arroyo Cienaguitas in Mendoza, and from contiguous beds. This species, (Perisphinctes colubrinus (Reinecke) p. 62, pl. 15, fig. 11), is very similar to Aulacosphinctes proximus (Steuer, 1897), both in general form and ornamentation pattern, but the ventral furrow is not visible on the last whorl preserved, although it is present on previous ones. Nevertheless the specimen illustrated is 83 mm in diameter, so this character may not be noticeable at that size, although it might be quite important in smaller shells as can be clearly seen on the specimens studied by Burckhardt (1900, p. 44, pl. 24, figs. 5, 6; pl. 26, fig. 4) and Weaver (1931, p. 413, pl. 44, figs. 301-303), from Mendoza and Neuquén, respectively. Weaver (1931, p. 411, pl. 44, figs. 298, 299) also described a specimen that was assigned to Aulacosphinctes proximus. However, his description does not mention any rib interruption over the venter at any shell size, an important character in the definition of the genus that casts doubts on its assignation. Aulacosphinctes wanneri Krantz (1928, p. 42, pl. 2, figs. 6a, b) from Arroyo La Manga, Mendoza, closely resembles Aulacosphinctes proximus. The development of a trifurcated rib is the only difference between the two species, an unimportant character that may be considered as intraspecific variation. Finally, the studied specimens are comparable with those studied by Leanza (1980, p. 44, pl. 6, figs. 2a, b, 4a, b, 5a, b; 1981, p. 587, pl. 2, figs., 9, 10) from the Middle Tithonian of Cerro Lotena in Neuquén.
Ocurrence: Middle Tithonian, Aulacosphinctes proximus Zone.
Family Neocomitidae Salfeld, 1921 Subfamily Berriasellinae Spath, 1922 Genus Parodontoceras Spath, 1923 a
Parodontoceras calistoides (Behrendsen, 1891) Fig. 5 t
1891 Hoplites calistoides Behrendsen, p. 402, pl. 23, figs. 1a, b 1897 Odontoceras calistoides Behrendsen; Steuer, p. 41, pl. 17, figs. 13-16 1921 Odontoceras calistoides Behrendsen; Steuer, p. 69, pl. 17, figs. 13-16 1922 Hoplites calistoides Behrendsen, p. 184, pl. 1, figs. 11a, b 1923a Parodontoceras callistoides Behrendsen; Spath, p. 305 (Gen. nov.) 1925 Berriasella calistoides Behrendsen; Gerth, p. 88 1928 Berriasella (Parodontoceras) calistoides (Behrendsen); Krantz, p. 24 1945 Parodontoceras calistoides (Behrendsen); Leanza, p. 41, pl. 5, figs. 5, 6 1988 Paradontoceras calistoides (Behrendsen); Riccardi, pl. 3, figs. 3, 4
Material: 1 specimen (CPBA 20554) from the Piuquenes Pass, Mendoza.
Description: Medium to large phragmacone (97.2 mm diameter), laterally crushed. Shell ellipticone, involute (U c. 22 % of diameter), with poorly developed umbilical border and quite inclined umbilical slope. Flanks are gently convex and venter somewhat flattened. Whorls are higher than wide, with the maximum width probably located at the umbilical border. The whorl height increases considerably with diameter, resulting in an eccentric shaped shell. Ornamentation consists of moderately strong ribs, regularly spaced, that begin at least from the middle part of the umbilical slope in a rursiradiate manner, then bend forwards at the umbilical border describing an inflection, and then another inflection at the bifurcation point in the middle of the flanks. These biplicate ribs are intercalated with simple ribs and some intercalatory ones do not go beyond the external third of the flank. All ribs are interrupted over the venter by a relatively wide and shallow furrow, which becomes less visible as diameter increases until it disappears completely. Suture line is very badly preserved.
Dimensions of specimen
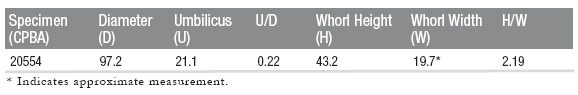
Remarks: Specific assignation of the specimen to Parodontoceras calistoides (Behrendsen, 1891, p. 402, pl. 23, figs. 1a, b) is quite safe despite its rather poor preservation, especially taking into account its eccentric shape, rib pattern and ventral furrow. This species is known from many localities in Neuquén and Mendoza, almost always in association with Substeueroceras koeneni (Steuer, 1897) characterized by a finer and denser rib pattern than Parodontoceras calistoides. Nevertheless, they seem to be very close forms and only a future more detailed study will prove if they are not just morphotypes of the same species.
Ocurrence: Uppermost Tithonian, Substeueroceras koeneni Zone. Family Holcodiscidae Spath, 1923 b Genus Spitidiscus Kilian, 1910
Spitidiscus riccardii Leanza and Wiedmann, 1992 Fig. 5 n
1992 Spitidiscus riccardii Leanza and Wiedmann, p. 33, figs. 4a-b 1992 Spitidiscus aff. S. rotula (J. de C. Sowerby); Leanza and Wiedmann, p. 32, figs. 3a-b 1992 Spitidiscus aff. S. gastaldianus (d'Orbigny); Leanza and Wiedmann, p. 32, figs. 6a-b 1995 Spitidiscus riccardii Leanza and Wiedmann; Aguirre-Urreta, p. 407, pl. 1, figs. 1-23 1999 Spitidiscus riccardii Leanza and Wiedmann; Aguirre-Urreta et al., pl. 1, fig. 4. 2005 Spitidiscus riccardii Leanza and Wiedmann; Aguirre-Urreta et al., figs. 7 e-f
Material: 1 specimen (CPBA 20557) and several fragments and impressions from the Piuquenes Pass, Mendoza.
Description: Shell small, involute (approx. diameter 30 mm), with rounded umbilical slope and slightly rounded flanks converging towards a curved venter. Ornament is composed by ribs and constrictions. The ribs are fine, arise from the umbilical slope and bifurcate in the lower third of the flank where they curve into a falcoid shape. They cross the venter without interruption bending slightly towards the aperture. The constrictions are less curved than the ribs and they dissect them. Suture line not preserved.
Remarks: Although the preservation of the specimens is rather poor, they can be assigned to Spitidiscus riccardii Leanza and Wiedmann, 1992 due to the size, involution of the shell and ribbing pattern. The studied specimens are comparable with those studied by Aguirre-Urreta (1995, p. 407, pl. 1, figs. 1-23) from Agua de la Mula and Agrio del Medio in Neuquén where the species has been studied in detail.
Occurrence: Upper Hauterivian. Spitidiscus riccardii Zone.
CONCLUDING REMARKS
When one compares the geological section of the Piuquenes Pass by Darwin (1846, pl. 1, skecth 1) with the present knowledge of the area (Pángaro et al. 1996, Fig. 2 herein), it can be seen that Darwin was able to clearly identify the succession of Meso-Cenozoic rocks and some structural features. He only partially recognized the highly complex structure of thrusts that affects the sedimentary rocks, repeating parts of the sequence. Regarding the fossil content, it must be understood that Darwin just rode along the trail, suffering the effects of high altitude and that he was being hurried by the "baqueanos" who were afraid of bad weather. He wrote to Henslow: "It was late in the Season, & the situation particularly dangerous for Snow storms. I did not dare to delay, otherwise a grand harvest might have been reaped" (Burkhardt and Smith 1985). He was able to collect the most common fossil, his big oyster Exogyra (Gryphacea) couloni (presently Aetostreon), other bivalves, some gastropods and ammonites. It is difficult to know precisely from where he got his specimens, though it seems he stopped several times along the trail collecting loose material, a fact that would explain why the fossils were poorly preserved. D'Orbigny (manuscript, see Fig. 4, and also Darwin 1846) regarded the assemblage as Neocomian age, based not only in the oysters but also in the presence of an indeterminate ammonite resembling Ammonites recticostatus d'Orbigny, 1840. This species is presently assigned to the genus Macroscaphites Meek, 1876 of Barremian-Early Aptian age, and is completely unknown in the Neuquén Basin, where the youngest ammonites are of Early Barremian age (Aguirre-Urreta et al. 2005). Most probably d'Orbigny confused it with some Tithonian perisphinctids which are common in the area. In spite of the poor preservation of the specimens and the lack of knowledge of faunas of this age in South America, it should be emphasized that Darwin was correct in assigning the whole succession to the Neocomian, following d'Orbigny's advice. This fauna was one of the first to be documented in the High Cordillera of the Main Andes, which now we know that also includes the Tithonian as seen by the ammonites described above.
ACKNOWLEDGEMENTS
The authors wish to acknowledge Dario Lazo for his information on the bivalves and Victor A. Ramos for his assistance in the field and his advice on the structure of the region. The reviewers Estanislao Godoy, Héctor Leanza and Amaro Mourgues improved the original manuscript with their comments.
WORKS CITED IN THE TEXT
1. Aguirre-Urreta, M.B. 1995. Spitidiscus riccardii Leanza y Wiedmann (Ammonoidea) en el Hauteriviano del Neuquén. Ameghiniana 32: 407-410.
2. Aguirre-Urreta, M.B. 1996. El Tithoniano marino de la vertiente argentina del paso de Piuquenes, Mendoza. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos (Buenos Aires), Actas 2: 185.
3. Aguirre-Urreta, M.B. 2001. Marine Upper Jurassic-Lower Cretaceous Stratigraphy and Biostratigraphy of the Aconcagua-Neuquén Basin, Argentina and Chile. Journal of Iberian Geology 27: 71-90.
4. Aguirre-Urreta, M.B., Concheyro, A., Lorenzo, M., Ottone, E.G. and Rawson, P.F. 1999. Advances in the biostratigraphy of the Agrio Formation (Lower Cretaceous) of the Neuquén basin, Argentina: ammonites, palynomorphs, and calcareous nannofossils. Paleogeography, Palaeoclimatology, Palaeoecology 150: 33-47.
5. Aguirre-Urreta, M.B., Rawson, P.F., Concheyro, G.A., Bown, P.R. and Ottone, E.G. 2005. Lower Cretaceous Biostratigraphy of the Neuquén Basin. In Veiga, G., Spalletti, L., Howell, J.A. and Schwarz, E. (eds.) The Neuquén Basin: A case study in sequence stratigraphy and basin dynamics. The Geological Society, Special Publication 252: 57-81, London.
6. Behrendsen, O. 1891-1892. Zur Geologie des Ostabhanges der argentinischen Kordillere. Deutsche Geologische Gesellschaft 43 (1891): 369-420; 44 (1892): 1-42.
7. Behrendsen, O. 1922. Contribución a la geología de la pendiente oriental de la Cordillera Argentina. Academia Nacional de Ciencias, Actas 7: 157-227, Córdoba.
8. Burckhardt, C. 1900. Profils géologiques transversaux de la Cordillère Argentino-Chilienne. Museo de La Plata, Sección Geológica y Mineralógica, Anales 2: 1-136.
9. Burckhardt, C. 1903. Beitrage zur Kenntniss der Jura und Kreide formation der Cordillere. Palaeontographica 50: 1-144.
10. Burkhardt, F. and Smith, S. 1985. The Complete Correspondence of Charles Darwin. Vol. 1. 1821-1836. Cambridge University Press, 752 p., Cambridge.
11. Camacho, H. H. 1953. Algunas consideraciones sobre los "Aporrhaidae" fósiles argentinos. Revista de la Asociación Geológica argentina 8: 183- 194.
12. Darwin, C. 1846. Geological observations on South America, being the third part of the geology of the voyage of the Beagle, under the command of Capt. FirtzRoy, RN, during the years 1832 to 1836. Smith, Elder and co., 268 p. London.
13. d'Orbigny, A. 1840-1841. Paléontologie Francaise. Description des Mollusques et Rayonnés Fossiles. Terrains Crétacés. Masson, 662 p., Paris.
14. d'Orbigny, A. 1842. Voyage dans L'Amérique Méridionale. Tome 3, 4° part: Paleontologie, 188 p., Paris & Strasbourg.
15. Fatmi, A.N. 1972. Stratigraphy of the Jurassic and Lower Cretaceous rocks and Jurassic ammonites from northern areas of West Pakistan. British Museum of Natural History 20: 297-380.
16. Gerth, E. 1925. La fauna Neocomiana de la Cordillera argentina, en la parte meridional de la provincia de Mendoza. Academia Nacional de Ciencias, Actas 9: 57-132, Córdoba.
17. Giambiagi, L., Tunik, M., Ramo, V. A. and Godoy, E. 2009. The High Andean Cordillera of Central Argentina and Chile along the Piuquenes Pass- Cordón del Portillo transect: Darwin's pioueering observations compared with modern geology. Revista de la Asociación Geológica Argentina 64 (1): 44-54.
18. Groeber, P. 1953. Mesozoico. In Geografía de la República Argentina, Sociedad Argentina de Estudios Geográficos GAEA 2(1): 9-541, Buenos Aires
19. Haupt, O. 1907. Beiträge zur Fauna des oberen Malm und der unteren Kreide in der argentinischen Cordillere. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 23: 187-236.
20. Imlay, R.W. 1942. Late Jurassic fossils from Cuba and their economic significance. Geological Society of America, Bulletin 53: 1417-1478.
21. Indans, J. 1954. Eine Ammonitenfauna aus dem Untertithon der argentinischen Kordillere in Süd-Mendoza. Palaeontographica 105 A: 96- 132.
22. Jordan, T., Isacks, B., Ramos, V.A. and Allmendinger, R.W. 1983. Mountain building model: The Central Andes. Episodes 1983(3): 20-26.
23. Kilian, W. 1910. Erste Abteilung: Unterkreide (Palaeocretacicum). Lieferung 2: Das bathyale Palaeocretacicum im südostlichen Frankreich; Valendis-Stufe; Hauterive-Stufe; Barreme-Stufe; Apt-Stufe. In Frech, F., Lethaea Geognostica. II. Das Mesozoicum, Band 3 (Kreide) (1907-1913), Schweizerbart, 169-288, pls. 1-8, Stuttgart.
24. Krantz, F. 1926. Die ammoniten des Mittel-und Obertithons. In Jaworski, E., Krantz, F. and Gerth, E. (eds.) Beiträge zur Paläontologie und Stratigraphie des Lias, Doggers, Tithons und der Unterkreide in der Kordillere im Süden der Provinz Mendoza (Argentinien). Geologische Rundschau 17a: 427-462.
25. Krantz, F. 1928. La fauna del Titono superior y medio en la parte meridional de la provincia de Mendoza. Academia Nacional de Ciencias, Actas 10: 1-57, Córdoba.
26. Leanza, A.F. 1945. Ammonites del Jurásico Superior y del Cretácico Inferior de la Sierra Azul, en la parte meridional de la provincia de Mendoza. Anales del Museo de La Plata NS 1: 1-99.
27. Leanza, H.A. 1980. The Lower and Middle Tithonian Ammonite Fauna from Cerro Lotena, Province of Neuquén, Argentina. Zitteliana 5: 3-49.
28. Leanza, H.A. 1981. Faunas de ammonites del Jurásico superior y del Cretácico inferior de América del Sur, con especial consideración de la Argentina. In Volkheimer, W. and Musacchio, E.A. (eds.) Cuencas sedimentarias del Jurásico y Cretácico de América del Sur. Comité Sudamericano del Jurásico y Cretácico 2: 559-597, Buenos Aires.
29. Leanza, H.A. and Wiedmann, J. 1992. Nuevos Holcodiscidae (Cephalopoda-Ammonoidea) del Barremiano de la Cuenca Neuquina, Argentina y su significado estratigráfico. Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1992, 1: 24-38.
30. Malumián, N. and Ramos, V.A. 1984. Magmatic intervals, transgression regression cycles and oceanic events in the Cretaceous and Tertiary of Southern South America. Earth and Planetary Science Letters 67: 228 237.
31. Pángaro, F., Ramos, V.A. and Godoy, E. 1996. La Faja plegada y corrida de la Cordillera Principal de Argentina y Chile a la latitud del Cerro Palomares (33°35'S). 13° Congreso Geológico Argentino y 3° Congreso Exploración de Hidrocarburos (Buenos Aires), Actas 2: 315-324.
32. Parent, H. 2001. The Middle Tithonian (Upper Jurassic) ammonoid fauna of Cañadón de los Alazanes, southern Neuquén-Mendoza Basin, Argentina. Boletín del Instituto de Fisiografía y Geología 71: 19-38, Rosario.
33. Polanski, J. 1964. Descripción geológica de la Hoja 25a Volcán San José, provincia de Mendoza. Dirección Nacional de Geología y Minería, Boletín 98: 1-94, Buenos Aires.
34. Riccardi, A. 1983. The Jurassic of Argentina and Chile. In Moullade, M. and Nairn, A.E.M. (eds.) The Phanerozoic geology of the world II, The Mesozoic B, Elsevier, 201-263, Amsterdam.
35. Riccardi, A.C. 1988. The Cretaceous System of southern South America. Geological Society of America, Memoir 168: 1-161.
36. Salfeld, H. 1921. Kiel- und Furschenbildung auf der Schalenaussenseite der Ammonoifrrn in ihrer Bedeutung für die Systematik und Festlegung von Biozonen. Zentralblatt für Mineralogie, Geologie und Paläontologie 1921: 343-347.
37. Spath, L.F. 1922. On Cretaceous Ammonoidea from Angola, collected by Professor J.W. Gregory, D. Sc., F.R.S. Transactions of the Royal Society of Edinburgh 53: 91-160.
38. Spath, L.F. 1923 a. On ammonites from New Zealand. In Trechmann, Ch.T. (ed.) The Jurassic Rocks of New Zealand. Geological Society of London, Quarterly Journal 79: 246-312.
39. Spath, L.F. 1923 b. A monograph of the Ammonoidea of the Gault. Part 1. Palaeontographical Society, Monographs 75 (353): 1-72.
40. Spath, L.F. 1925. Ammonites and Aptychi. In Wyllie, B.N.K and Smellie, W.E. (eds.) The collection of fossils and rocks from Somaliland. Hunterian Museum, Glasgow University, Monograph 7: 111-164.
41. Spath, L.F. 1927-1933. Revision of the Jurassic Cephalopod Fauna of Kachh (Cutch). Paleontologia Indica, NS 9, 1 (1927): 1-72, 2 (1928): 73-161, 3 (1928): 163-278, 4 (1931): 279-550, 5 (1931): 551-658, 6 (1933): 659-945.
42. Steinmann, G. 1890. Cephalopoda. In G. Steinmann and L. Döderlein (eds.) Elemente der Paläontologie, 344-475, W. Engelmann, Leipzig.
43. Steuer, A. 1897. Argentinische Jura - Ablagerungen: Eine Beitrage zur Kenntniss der Geologie und Paleontologie der argentinischen Anden. Palaeontologische Abhandlungen NS 7: 127-222, Jena.
44. Steuer, A. 1921. Estratos Jurásicos Argentinos. Contribución al conocimiento de la Geología y Paleontología de los Andes Argentinos entre el Río Grande y el Río Atuel. Academia Nacional de Ciencias, Actas 7: 5-128, Córdoba.
45. Toucas, A. 1890. Étude sur la fauna des couches tihoniques de l'Ardèche. Bulletin de la Société Géologique de France (Serie 3) 18: 560-629.
46. Uhlig, V. 1910. The Fauna of the Spiti Shales. Palaeontologia Indica 15: 133-395.
47. Waagen, W. 1873-1875. Jurassic Fauna of Kutch. Palaeontologia Indica 1 (1873): 1-22, 2 (1875): 23-76, 3 (1875), 77-106, 4 (1875): 107-247.
48. Weaver, C. 1931. Paleontology of the Jurassic and Cretaceous of West Central Argentina. Memoirs of the University of Washington 1: 1-496.
49. Zittel, K.A. 1884. Cephalopoda. In Zittel, K.A. Handbuch der Palaeontologie, R.Oldenbourg, Band 1, Abteilung 2, Lieferung 3, 893 p., Munich and Leipzig..
50. Zittel, K.A. 1895. Grundzüge der Palaeontologie. R. Oldenbourg, viii + 971 p., 2.048 figs, Munich and Leipzig.
Laura Giambiagi1, Maisa Tunik2, Victor A. Ramos3, and Estanislao Godoy4
1 IANIGLA - CONICET, Mendoza. Email: lgiambia@lab.cricyt.edu.ar 2 CONICET-CIMAR - Universidad Nacional del Comahue. 3 Laboratorio de Tectónica Andina (FCEN) Universidad de Buenos Aires - CONICET. 4 Geólogo Consultor, Santiago de Chile.
ABSTRACT: The geological observations made by Darwin in 1835 during his crossing of the Andes from Santiago to Mendoza via the Piuquenes Pass and Cordón del Portillo are compared with the present geological knowledge of the Cordillera Principal and Cordillera Frontal at 33°-34°S. The analysis of the complex stratigraphy of the Cordillera Principal, the imbricated structure of the Aconcagua fold and thrust belt, as well as the stratigraphy and structure of the inter mountain foreland Tunuyán Basin, allows to assess the pioneer observations of Darwin. He recognized the old metamorphic basement and the granitoids and volcanic sequences of late Paleozoic to Triassic age of the Cordillera Frontal, established the Cretaceous age of the marine successions cropping out along the eastern Cordillera Principal and studied the conglomeratic deposits associated with the uplift of the Cordillera in the Alto Tunuyán Basin. Based on the study of clast provenance of the synorogenic deposits of the Alto Tunuyán Basin, Darwin recognized that the Cordillera Frontal was uplifted later than the Cordillera Principal. The present knowledge of this sector of the Andean Cordillera confirms his pioneer observations and show that Darwin was one of the first scientists ever in realizing that in an orogenic system the sequence of uplift and deformation proceeds from hinterland towards foreland, according to a process that is exceptionally well-illustrated along the Piuquenes-Cordón del Portillo transect.
Keywords: Uplift; Provenance; Structure; Metamorphic rocks; Synorogenic deposits.
RESUMEN: La Alta Cordillera de los Andes del centro de Argentina y Chile a lo largo de la transecta del Paso Piuquenes-Cordón del Portillo: Las observaciones pioneras de Darwin comparadas con la geología moderna. Las observaciones geológicas efectuadas por Darwin en 1935 durante su cruce de la Cordillera de Los Andes entre Santiago y Mendoza realizado en 1835 a través de los pasos del Portillo y Piuquenes son examinadas y comparadas con el conocimiento actual existente de las Cordilleras Principal y Frontal entre los 33°-34°S. El análisis de la compleja estratigrafía de la Cordillera Principal, la estructura de las diferentes láminas imbricadas de la faja plegada y corrida del Aconcagua y la cuenca de antepaís intermonatana del Tunuyán, permiten ponderar las pioneras observaciones de Darwin, quien reconoció el basamento metamórfico y los granitoides y secuencias volcánicas neopaleozoicas a triásicas de la Cordillera Frontal, estableció la edad cretácica de las sucesiones marinas del sector oriental de la Cordillera Principal y describió e interpretó los depósitos conglomerádicos asociados al levantamiento de los Andes en el Alto Tunuyán. Sobre la base del estudio de la proveniencia de los clastos de los depósitos sinorogénicos de la cuenca del Alto Tunuyán pudo reconocer que la Cordillera Principal, fue alzada con anterioridad a la Cordillera Frontal, la que a través de varios pulsos adquirió su altura actual en épocas más recientes. Darwin fue uno de los primeros en reconocer que en un sistema orogénico, la secuencia de deformación asociada al levantamiento orogénico comienza en su región interna desde donde avanza progresivamente hacia el antepaís, como lo demuestran sus brillantes observaciones efectuadas ya hace más de 150 años.
Palabras clave: Levantamiento; Procedencia; Estructura; Rocas metamórficas; Depósitos sinorogénicos.
INTRODUCTION
Charles Darwin, a naturalist who thought of himself as a geologist, crossed the Piuquenes and Portillo passes in 1835, during his voyage on board HMS Beagle. The trip, which lasted from March 19th to March 24th, was a milestone in his knowledge of Andean geology. It was in this sector of the Andes that Darwin suspected that the giant mountains were uplifted as late as in the Cenozoic. The idea was a radical one, as well as the interpretation that the different ranges were uplifted one after the other, being the easternmost one the youngest. Later generations of geologists demonstrated that he was essentially right in both concepts. The Piuquenes and Portillo passes are situated in the southern Central Andes, at 33°40'S (Fig. 1). At these latitudes the Andes are formed by the Cordillera Principal and the Cordillera Frontal. The Piuquenes Pass is situated in the Cordillera Principal, at the border between Argentina and Chile, and the Portillo Pass is located in the high summits of the Cordillera Frontal to the east (Fig. 2). At the time Darwin crossed the Andes through the Piuquenes Pass, this sector of the Cordillera Principal was known as the Peuquenes range or western line, and the Cordillera Frontal, as the Portillo range. These two ranges are separated, by a wide valley filled by Neogene sediments (Depresión Intermontana del Alto Tunuyán, Polanski 1957). The Cordillera Principal can be subdivided into a western sector, formed by a thick sequence of Oligocene and Miocene volcanic rocks and intrusives, and an eastern zone corresponding to the strata of Mesozoic sedimentary rocks deformed into the Aconcagua fold and thrust belt (Ramos 1988, Cegarra and Ramos 1996, Ramos et al. 1996a). It was observing these rocks that Darwin became astonished to see "shells which were once crawling on the bottom of the sea, now standing nearly 14,000 feet above its level" (Darwin 1845, p. 320).
 Figure 1: a) Location map of the High Andes with main geological provinces and lithological sequences between Santiago (Chile) and Mendoza (Argentina); b) Block diagram showing the main structures of the Cordillera Principal and Cordillera Frontal with location of the Aconcagua fold and thrust belt.
Figure 1: a) Location map of the High Andes with main geological provinces and lithological sequences between Santiago (Chile) and Mendoza (Argentina); b) Block diagram showing the main structures of the Cordillera Principal and Cordillera Frontal with location of the Aconcagua fold and thrust belt.
 Figure 2: a) Water divide between Chile and Argentina with indication of the Piuquenes Pass. The international landmark (hito) is located on Neocomian limestones; b) Portillo Pass across the Cordillera Frontal. Glacial deposits are overlying late Paleozoic granites; c) Approaching the international boundary along Darwin's trail.
Figure 2: a) Water divide between Chile and Argentina with indication of the Piuquenes Pass. The international landmark (hito) is located on Neocomian limestones; b) Portillo Pass across the Cordillera Frontal. Glacial deposits are overlying late Paleozoic granites; c) Approaching the international boundary along Darwin's trail.
TECTONIC SETTING
The Andes of central Argentina and Chile are a linear orogenic belt formed at the convergent plate margin between the Nazca and South American plates. At 33°-34° S the Nazca plate subducts beneath the South American plate at a relative velocity of nearly 10 cm/yr (Pardo-Casas and Molnar 1987). The distribution of seismicity suggests that the Nazca Plate is characterized by alternating flat and normal segments of subduction (Baranzangi and Isacks 1976, Cahill and Isacks 1992, Pardo et al. 2002). Several features of the Andes orogenic system appear to correlate with these changing subduction geometries, such as volcanism and distribution of morphostructural belts and tectonic style (Isacks et al. 1982, Jordan et al. 1983). North of 33°S, the flat subduction segment lacks active arc magmatism and underlies the Cordillera Principal, Cordillera Frontal, Precordillera and Sierras Pampeanas structural provinces while south of 34°S the "normal" subduction segment of south-central Argentina and Chile shows the active volcanism of the Southern Volcanic zone (see Hildreth and Moorbath 1988, Stern et al. 2007) and the orogenic system is much narrower and formed only by the Cordillera Principal and Cordillera Frontal. The study area (33°-34°S) is located along the transition zone between the flat and normal subduction segments (Fig. 3).
Figure 3: Geologic map of the Cordillera Principal and Cordillera Frontal between Santiago (Chile) and Mendoza (Argentina) based on Giambiagi et al. (2003). The trail followed by Darwin, as well as the different camps, are indicated day by day following Darwin's (1845) description.
STRATIGRAPHY
The stratigraphy, examined in many previous studies such as those of by Polanski (1964), Thiele (1980), Charrier et al. (1997), Ramos et al. (2000), Jordan et al. (2001), Giambiagi et al. (2001, 2002), Tunik (2003), Tunik et al. (2004), Kay et al. (2005), Fock ( 2005) and Farías et al. (2008) includes seven main major tectono- stratigraphic units (Fig. 4): (1) Proterozoic to Lower Triassic metamorphic, volcanic and intrusive rocks; (2) Triassic and Jurassic rift sequences; (3) Titho-Neocomian marine strata; (4) Cretaceous to Paleogene marine and non marine sedimentary rocks; (5) Eocene?-Oligocene to Lower Miocene volcanics; (6) Upper Cenozoic foreland basin deposits; and (7) Upper Cenozoic intrusives and volcanic-arc rocks.
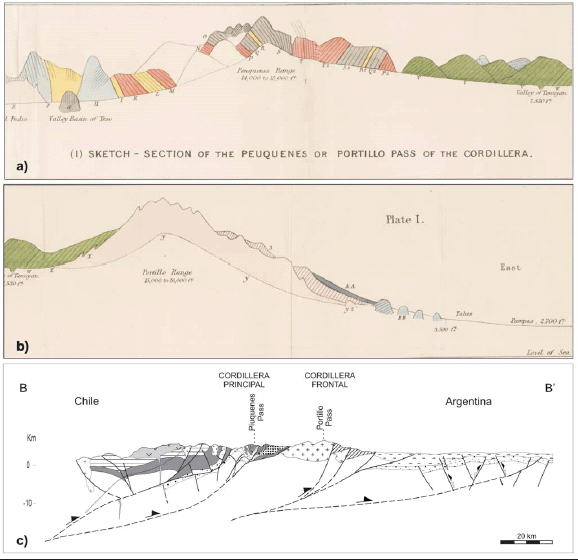 Figure 4: Structural cross sections across the High Andes: a) Section of the Piuquenes pass and b) Portillo pass after Darwin (1846); c) Modern structural cross-section based on Giambiagi and Ramos (2002). Location of the section in figure 3.
Figure 4: Structural cross sections across the High Andes: a) Section of the Piuquenes pass and b) Portillo pass after Darwin (1846); c) Modern structural cross-section based on Giambiagi and Ramos (2002). Location of the section in figure 3.
The oldest rocks correspond to Proterozoic mica-schists cropping out at Cordón del Portillo, in the Cordillera Frontal. These rocks were examined by Darwin at the "base of the Portillo range, ...at a place called Mal Paso. The mica-schist here consists of thick layers of quartz, with intervening folia of finely-scaly mica, often passing into a substance like black glossy clay-slate: in one spot, the layers of quartz having disappeared, the whole mass became converted into glossy clay-slate. Where the folia were best defined, they were inclined at a high angle westward, that is, towards the range... I think it more probable, considering its more perfect metamorphic character and its wellpronounced foliation, that it belongs to an anterior epoch" (Darwin 1846, p. 184). These rocks are unconformably overlain by Upper Paleozoic marine black shales and sandstones and are intruded by Carboniferous to Permian granitoids (Polanski 1964). These granitoids were clearly identified by Darwin who stated that "the red granite, from being divided by parallel joints, has weathered into sharp pinnacles, on some of which, even on some of the loftiest, little caps of mica-schist could be clearly seen: here and there isolated patches of this rock adhered to the mountain-flanks, and these often corresponded in height and position on the opposite sides of the immense valleys. Lower down the schist prevailed more and more, with only a few quite small points of granite projecting through. Looking at the entire eastern face of the Portillo range, the red color far exceeds in area the black; yet it was scarcely possible to doubt, that the granite had once been almost wholly encased by the mica-schist" (Darwin 1846, p. 184). Permian-Triassic intermediate and acid volcanic rocks unconformably overlie the previously deformed rocks. As Darwin pointed out, these volcanics reach a thickness of more than 1,700 m. He described these rocks and assigned them a relative age based on their metamorphism and vein intrusion. Rifting was widespread in central Argentina and Chile during Triassic - Early Jurassic times. This was a consequence of extensional processes linked to the initial stages of the fragmentation of Gondwana and the opening of the South Atlantic (Uliana et al. 1989). A series of NNW trending rift systems was formed at this time along the western margin of Gondwana (Charrier 1979, 1984, Uliana et al. 1989). In the Cordillera Principal, these rift basins, which at this latitude only crop out in the Chilean side of the Cordillera Principal, were filled by Lower to Middle Jurassic black shales (Godoy 1993, Alvarez et al. 1997, 1999). In the Argentinean side, a thick layer of evaporites (Auquilco Formation) unconformably rests upon Jurassic marine strata. This was observed by Darwin who was stunned by its more than 600 m thickness. The evaporites grade into the red sandstones and claystones with subordinated conglomerates of the Kimmeridgian Tordillo Formation. The Jurassic units in the Cordillera Principal are capped by black shales, mudstones, limestones and sandstones, deposited on a stable marine platform during Titho Neocomian thermal times (Biro 1964, Polanski 1964, Aguirre-Ureta 1996). These sedimentary rocks, now grouped into the Mendoza Group, were described in detailed by Darwin who assigned them a Neocomian age based on his collection of invertebrate fossils (see Aguirre-Urreta and Vennari, 2009). The Upper Cretaceous strata, se-parated from the Lower Cretaceous units by an erosional unconformity, consist of conglomerates, sandstones and volcaniclastic rocks. The presence of a continuous interval of Maastrichtian to Da-nian siltstones and limestones deposited during an Atlantic ingression that reached up to the eastern slope of the present day Cordillera Principal (Tunik 2003, Tunik et al. 2004) indicates a subdued relief close to sea level for this region at that time. After at least 10 Ma of magmatic quiescence, volcanic activity considerably increased in the western Cordillera Principal during the Eocene?-early Miocene (Godoy et al. 1996, Charrier et al. 1997, Jordan et al. 2001, Kay et al. 2005, Fock 2005). This volcanic event corresponds to the Abanico and Coya Machalí Formations of Late Eocene to Middle Miocene age (Wyss et al. 1990, Flynn et al. 1995, Gana and Wall 1997, Charrier et al. 2002, Farías et al. 2008). that crop out in two north-trending belts made up of basic lavas, tuffs and intermediate pyroclastic rocks, interbedded with sedimentary rocks totalling at least 2,000 m thick. Eocene ages are reported only on the western belt while, in general, volcanic rocks have a primitive isotopic signature indicating crustal thinning and high paleothermal gradient (Nystrom et al. 1993, Vergara et al. 1988, 1999) during the accumulation of these sequences, probably in a volcano-tectonic extensional /transtensional basin (Charrier et al. 1997, 2002, Godoy et al. 1999, Jordan et al. 2001, Fock 2005). The Abanico and Coya Machalí Formations are covered by 2,000 m of Miocene calc-alkaline andesitic lava and acid pyroclastic flows of the Farellones Formation from which radiometric ages spanning from 20 Ma to 7 Ma have been reported (Rivano et al. 1990). This unit is intruded by several epizonal granitoid plutons and stocks, some of them associated with the giant copper-porphyry copper deposits of Río Blanco and El Teniente with show radiometric ages constrained between 10 to 6 Ma (Cornejo and Mahood 1997, Kurtz et al. 1997, Maksaev et al. 2004, Deckart et al. 2005). During this journey across the Cordillera Principal Darwin observed massive rocks which "rarely contain any quartz" intruded among the volcanic strata that probably correspond to sub volcanic dykes and sills emplaced in the volcanic sequences. Finally, the youngest geological units of the Cordillera Principal at this latitude correspond to large composite Quaternary stratovolcanoes such as the Tupungato, San Juan, Marmolejo and San José, being these the northernmost volcanic centers of the Southern Volcanic Zone (SVZ) of the Andes (Hildreth and Moorbath 1988, Stern et al. 2007). Along the eastern slope of the Cordillera Principal, in Argentina, the oldest Cenozoic volcanic unit is the Contreras Formation, exposed at the base of a thick sequence of Neogene synorogenic deposits, and consisting of basaltic lava flows and breccias. A sample from the middle sector of this unit yielded a whole rock K-Ar age of 18.3 Ma (Giambiagi et al. 2001). Its geochemistry suggests a retroarc setting in a thickening crust during its extrusion (Ramos et al. 1996b). Late Cenozoic foreland clastic deposits occur to the east of the Cordillera Principal. Miocene sedimentary rocks filling the Alto Tunuyán Basin include three units. The oldest Tunuyán Conglomerate consists of up to 1,400 m of conglomerates and sandstones deposited in an alluvialfan setting (Fig. 5b).
 Figure 5: a) Tunuyán and Palomares rivers, looking southward. Cretaceous marine strata unconformably rest over the Paleozoic basement rocks of the Cordón del Portillo; b) Tunuyán Conglomerate in the Palomares river; c) Tuffs and conglomerates of the Palomares Formation unconformably resting over the Tunuyán Conglomerate; d) Butaló Formation.
Figure 5: a) Tunuyán and Palomares rivers, looking southward. Cretaceous marine strata unconformably rest over the Paleozoic basement rocks of the Cordón del Portillo; b) Tunuyán Conglomerate in the Palomares river; c) Tuffs and conglomerates of the Palomares Formation unconformably resting over the Tunuyán Conglomerate; d) Butaló Formation.
The vertical variation in clast composition and paleocurrent data reflect the progressive erosion and unroofing of the Aconcagua fold and thrust belt (Giambiagi 1999). The synchronism of deposition of the upper 400 m of this unit with the emplacement of thrusts within the basin is documented by syntectonic unconformities. Deposition of the Tunuyán Conglomerate started around 18 to 17 Ma, and continued to ~10 Ma. The intermediate Palomares Formation filled a gentle paleorelief in the Tunuyán Conglomerate between 8.5 and 7 Ma and consists of 200 m of volcaniclastic and clastic sediments deposited in an alluvialfan setting (Fig. 5c). The Cordillera Frontal was the main source area of this unit, as indicated by clast composition, paleocurrent measurements and syntectonic geometries (Giambiagi et al. 2001). The Butaló Formation records the final infilling of the sedimentary trough, reaching a thickness of more than 300 m, and is made up of fluvial and lacustrine deposits (Fig. 5d). It is unconformably overlain by andesitic volcanic rocks dated at 5.9 Ma (whole rock K/Ar age, Ramos et al. 2000). Darwin's description of the Conglomerate of Tenuyan (sic) indicated that "the included pebbles are either perfectly or only partially rounded: they consist of purplish sandstones, of various porphyries, of brownish limestone, of black calcareous, compact shale precisely like that in situ in the Peuquenes range, and containing some of the same fossil shells; also very many pebbles of quartz, some of micaceous schist, and numerous, broken, rounded crystals of a reddish orthitic or potash feldspar (as determined by Professor Miller), and these from their size must have been derived from a coarse-grained rock, probably granite. From this feldspar being orthitic, and even from its external appearance, I venture positively to affirm that it has not been derived from the rocks of the western ranges; but on the other hand it may well have come, together with the quartz and metamorphic schists, from the eastern or Portillo line, for this line mainly consists of coarse orthitic granite. The pebbles of the fossiliferous slate and of the purple sandstone, certainly have been derived from the Peuquenes or western ranges." (Darwin 1845, p. 182).
STRUCTURE
Aconcagua Fold and Thrust Belt
During his ride through the Piuquenes Pass Darwin felt admiration for the inclined to overturned Mesozoic layers which had been formed undersea and later uplifted and deformed in an "extraordinary manner", reaching an altitude of more than 4,000 m (Figs. 4 a and b). Presently, the Argentine eastern sector of the Cordillera Principal is interpreted to form part of the Aconcagua fold-andthrust belt. This belt can be divided into two do-mains (Fig. 6b, c).
Figure 6: a) Geological map of the Aconcagua fold-and-thrust belt in the Yeso and Palomares river (modified from Giambiagi et al. 2003) See location in figure 3; b) Structural cross-section A-B (Giambiagi and Ramos 2002); c) The Chacayal thrust in the Chilean slope of the Cordillera Principal, uplifting Late Jurassic red beds (Tordillo Formation) over Early Cretaceous continental and marine sequences (Colimapu Formation and Mendoza Group); d) Southern view of the Cerro Palomares. The Palomares fault system uplifts Cretaceous limestones over the Neogene synorogenic strata of the Tunuyán Conglomerate (a) and the Palomares Formation (b).
The structure of the western domain is dominated by open NNW trending folds and N-S thrust faults. The majority of these thrusts cut the previously developed fold structures and show an eastward vergence, with the exception of a belt of west verging back-thrusts located at the border between Argentina and Chile (Fig. 4c). Although the basement does not crop out, its involvement is deduced from the geometry of the structure and significant thickness variations of the deformed Mesozoic successions (Giambiagi et al. 2003). The eastern domain is broad and narrows towards the south until it disappears south of Volcán Marmolejo. This domain is characterized by a dense array of imbricate low-angle thrusts which exhibits flat and ramp geometries, with flats corresponding to decóllement levels located on the Late Jurassic evaporites and Early Cretaceous shales. In this sector of the belt it is common to observe thrusts cutting previously developed structures, some of them showing younger-over-older relationships. This is the reason why it is interpreted as evolved by a forward propagating thrust sequence with periods of out-of-sequence thrusting
The Alto Tunuyán Foreland Basin
The Alto Tunuyán Basin is a Neogene foreland basin located between the Cordillera Principal and the Cordillera Frontal, from 33°30' to 34°S. The basin was generated by lithospheric flexure as response to thrust belt load of the Aconcagua fold and thrust belt and was filled with more than 1,800 m of continental synorogenic deposits of the Tunuyán Conglomerates and the Palomares and Butaló Formations. These strata were deformed as the Aconcagua fold-andthrust belt deformation migrated eastward and cannibalized the foreland basin. One of Darwin's first geological observation was that the constitutions of the two ranges are totally different and that "one part of the double line of mountains (Cordilleras Principal and Frontal) is of an age long posterior to the other", which is nowadays known as the normal sequence of deformation and uplift of an orogenic system, from the hinterland to the foreland. It is interesting to remark how Darwin, based on a provenance analysis of conglomeradic clasts, was able to infer the order and the relative uplift age of each mountain chain across this section of the Andes. He noted that between the two main ranges "there rest beds of a conglomerate several thousand feet in thickness, which have been upheaved ... and dip at an angle of 45° towards the Peuquenes line. I was astonished to find that this conglomerate was partly composed of pebbles, derived from the rocks, with their fossil shells, of the Peuquenes range; and partly of red potash granite, like that of the Portillo. Hence we must conclude, that both the Peuquenes and Portillo ranges were partially upheaved and exposed to wear and tear, when the conglomerate was forming; but as the beds of the conglomerate have been thrown off at an angle of 45° by the red Portillo granite ..., we may feel sure, that the greater part of the ... upheaval of the already partially formed Portillo line, took place after the accumulation of the conglomerate, and long after the elevation of the Peuquenes ridge. So that the Portillo, the loftiest line in this part of the Cordillera, is not so old as the less lofty line of the Peuquenes. Evidence derived from an inclined stream of lava at the eastern base of the Portillo, might be adduced to show, that it owes part of its great height to elevations of a still later date. ... In most parts, perhaps in all parts, of the Cordillera, it may be concluded that each line has been formed by repeated upheavals" (Darwin 1845, p. 320). These conglomerates are now part of the Alto Tunuyán Basin (Polanski 1957), an intermontane basin studied by Giambiagi (1999), who confirmed the uplift sequence by means of precise provenance studies. The Alto Tunuyán Basin structure comprises thin-skinned fold-thrust sheets above a decóllement developed in Late Cretaceous siltstones. Within this zone, faults related to syntectonic unconformities recorded in the upper part of the Tunuyán Conglomerate indicate sedimentation above an active thrust (Giambiagi et al. 2001). Other faults indicate activity after deposition of the Neogene units. The structure in this domain intrigued Darwin, who observed that the newer coarse conglomerates dipped directly under the much older beds of gypseous and limestone units. He stated that the Mesozoic strata has been broken up by "several, distinct, parallel, unilinal lines of elevation" and estimated a vertical throw of over 600 m. These lines of elevation represent the complex thrust system of the Cerro Palomares (Fig. 6d).
Cordillera Frontal
The Cordillera Frontal consists of several basement blocks uplifted east of the Aconcagua fold-and-thrust belt. During the Late Cenozoic compression - at the latitude of the Portillo Pass- it behaved as a rigid block disrupted by medium to high angle faults showing two regional trends of structures, i.e., NNE-trending faults in the northern region and N to NNW-trending faults in the southern half, related to Late Paleozoic structures. A deep crustal detachment level is required in order to propagate shortening within the Cordillera Frontal. This detachment is inferred to be located between 15 and 20 km deep, probably within a brittle-ductile transition.
GEOMORPHOLOGY
Darwin stated "in the course of ages in this (referring to the Cachapoal) and other valleys, events may have occurred like between, but even on a grander scale than, that described by Molina, when a slip during the earthquake of 1762, banked up for ten days the great river Lontué, which then bursting its barrier inundated the whole country and doubtless transported many great fragments of rocks" (Darwin 1846, p. 66-67). However, "not withstanding this one case of difficulty" he then claimed that "one of the most important conclusions to which my observations on the geology of South America have led me", is his lengthily exposed conviction that the "terrace-like fringes, which are continuously united with the basin-shaped plains at the foot of the Cordillera, have been formed by the arrestment of riverborne detritus at successive levels, in the same manner as we see now taking place at the heads of all those many, deep, winding fiords intersecting the southern coasts." Due to this view Darwin failed to recognize the Meson Alto landslide and the plain in the Yeso valley (Embalse El Yeso in Fig. 6a) and refers to it as "the one exception in which a range was dislocated by a great and abrupt fault" In page 178 this plain is described as a "drained lake", yet once more no relation to a landslide is mentioned. The dam of this paleolake was later described as a moraine and only recently has it been recognized as part of a big landslide derived from the granitic southern slopes of the Yeso valley (Abele 1981).
STRUCTURAL EVOLUTION OF THE ANDES AT 33°40'S
Here we integrate recent structural and sedimentological studies carried out at 33°-34° S (Godoy 1993, Godoy et al. 1996, 1999, Ramos et al. 1996b, 1997, Giambiagi et al. 2001, Giambiagi and Ramos 2002, Giambiagi et al. 2003) as a way to understand what was envisaged by Darwin's during his early observations. During the early Miocene - between 20? and 15 Ma - shortening associated with inversion of the Mesozoic and Early Cenozoic extensional systems started diachronically in the Cordillera Principal. At around 18 Ma deformation and uplift generated a western sediment source for the lower part of the foreland Tunuyán Conglomerate, which at 15 Ma shows an abrupt change in clast composition indicated by the first appearance of Mesozoic sedimentary debris (Giambiagi 1999, Irigoyen et al. 2000). This is considered as evidence for a second phase of deformation, linked to E-W shortening along the eastern Cordillera Principal. Continued migration of deformation toward the foreland is documented by the occurrence of growth structures in the upper section of the Tunuyán Conglomerate. At that time basement thrusts transported eastwards earlier Cordillera Principal thrusts and folds, forming thus a hybrid thin and thick-skinned belt of deformation. Overprinting relationships indicate that the fold and thrust belt evolved by means of a forward propagating thrust sequence including periods of out-of-sequence thrusting. The denudation of the fold and thrust belt provided the material for the uppermost levels of the Tunuyán Conglomerate. The unconformity that separates the Palomares Formation from the Tunuyán Conglomerate, the changes in paleocurrents measured in the former unit, and the occurrence of locally derived conglomerates, provide evidence for the beginning of a third phase of deformation, associated this time with the onset of uplifting in the Cordillera Frontal. This is recorded by the regional influx of thick coarse conglomerates, the generation of a broken foreland basin system, and the accumulation of the wedge of proximal synorogenic deposits of the Palomares and Butaló Formations. The uplift of the Cordillera Frontal generated a sticking point, preventing the propagation of the thrust belt towards the foreland. As a result, a series of out-ofsequence thrusts developed in the eastern Cordillera Principal and the Alto Tunuyán foreland basin was partially cannibalized by the advancing thrust sheets. After the emplacement of granitoids with isotopic signatures indicating magmas sources in a thickened crust (Kurtz et al. 1997, Kay et al. 2005) in the western Cordillera Principal, magmatism shifted eastward to its present position along the active volcanic front of the Tupungato-San José line. Thrusting along the Cordillera Principal ended by ~4 Ma and subsequently the deformation front migrated to the Precordillera - north of 33°Sand to the Cuyo Basin and the eastern Cordillera Frontal foothills south of 33°S. Folding in the Cuyo Basin started after the beginning of deposition of the Mogotes Formation at ~ 3.0 Ma (Yrigoyen 1993, Chiaramonte et al. 2000). The last phase of deformation accommodating ESE-WNW and ENE-WSW shortening concentrated in the Cuyo Basin and in the La Carrera thrust system in the Cordillera Frontal. Earthquake focal mechanisms and neotectonic activity indicate that the eastern border of the Precordillera and the Cuyo Basin are still active (Cortés et al. 1999, Chiaramonte et al. 2000).
CONCLUDING REMARKS
Darwin was the first geologist to describe the stratigraphy of the Cordillera Principal and the Cordillera Frontal of central Chile and Argentina recognizing the presence of an old metamorphic basement - much older than its "red granites" - presently considered to be late Paleozoic intrusives in the Cordillera Frontal. He correctly assigned a Cretaceous age to the marine sediments cropping out along the eastern Cordillera Principal and also showed that the uplift of the Andean range was much younger than the marine sedimentation of the Cretaceous deposits. By studying the composition of clasts included in Tertiary sediments of the Alto Tunuyán Basin he was able to establish the relative uplifting times for the Cordillera Principal and Cordillera Frontal conluding that the Piuquenes range (Cordillera Principal) was uplifted before the Portillo range (Cordillera Frontal). To explain the drainage pattern of the Tunuyán-Cordón del Portillo region, he proposed a gradual elevation of the Cordón del Portillo after the uplift of the Piuquenes range, which allowed the waters of the Tunuyán Depression to cut a deep canyon through the latter as the range was been uplifted. As a tribute to his exceptional observational skills we should emphasize that the progressive advance of an orogenic deformation front from hinterland towards foreland, now considered a common process in most mountain chains, was early envisaged by Darwin more than 150 years ago during is historic 1835 journey across the Andes.
ACKNOWLEDGEMENTS
This work was found by the University of Buenos Aires (UBACYT x182) and CONICET (PIP 4162, 5965). The authors acknwoledge with special thanks the reviews of Reynaldo Charrier, Miguel Griffin, and Constantino Mpodozis.
WORKS CITED IN THE TEXT
1. Abele, G. 1981. Trockene Massenbewegungen, Schlammströme und rasche Abflüsse. Mainzer Geographische Studien 23, 102 p., Mainz.
2. Aguirre-Urreta, M.B. 1996. El Tithoniano marino de la vertiente argentina del paso de Piuquenes, Mendoza. 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos (Buenos Aires), Actas 5: 185.
3. Aguirre-Urreta, B. and Vennari, V. 2009. On Darwin's footsteps across the Andes: Tithonian-Neocomian fossil invertebrates from the Piuquenes pass. Revista de la Asociación Geológica Argentina 64(1): 32-43.
4. Alvarez, P.P., Aguirre-Urreta, M.B., Godoy, E. and Ramos V.A. 1997. Estratigrafía del Jurásico de la Cordillera Principal de Argentina y Chile (33°45' - 34°00' LS). 8° Congreso Geológico Chileno, Actas 1: 425-429, Santiago.
5. Alvarez, P.P., Godoy, E. and Giambiagi, L. 1999. Estratigrafía de la Alta Cordillera de Chile Central a la latitud del paso Piuquenes (33°35' LS). 14° Congreso Geológico Argentino (Salta), Actas 1: 55.
6. Barazangi, B.A. and Isacks, B.L. 1976. Spatial distribution of earthquakes and subduction of the Nazca Plate beneath South America. Geology 4: 686-692.
7. Biro, L. 1964. Límite Titónico-Neocomiano en Lo Valdés. Memoria de título, Departamento de Geología, Universidad de Chile, unpublished, 118 p., Santiago.
8. Cahill, T. and Isacks, B.L. 1992. Seismicity and shape of the subducted Nazca Plate. Journal Geophysical Research 97: 17503-17529.
9. Cegarra, M. and Ramos, V.A. 1996. La faja plegada y corrida del Aconcagua. In Ramos, V.A. et al. Geología de la región del Aconcagua, Provincias de San Juan y Mendoza. Subsecretaría de Minería de la Nación, Dirección Nacional del Servicio Geológico, Anales 24: 387-422, Buenos Aires.
10. Charrier, R. 1979. El Triásico de Chile y regiones adyacentes de Argentina: una reconstrucción paleogeográfica y paleoclimática. Comunicaciones 26: 1-37.
11. Charrier, R. 1984. Areas subsidentes en el borde occidental de la cuenca tras-arco Jurásico-Cretácica, Cordillera Principal Chilena entre 34° y 34°30' S. 9° Congreso Geológico Argentino, Actas 2: 107-124.
12. Charrier, R., Baeza, O., Elgueta, S., Flynn, J.J., Gans, P., Kay, S.M., Muñoz, N., Wyss, A.R. and Zurita, E. 2002. Evidence for Cenozoic extensional basin development and tectonic inversión south of the flat-slab segment, southern Central Andes, Chile (33°-36°S.L.). Journal of South American Earth Sciences 15: 117-139.
13. Charrier, R., Flynn, J.J., Wyss, A.R., Zapatta, F. and Swisher, C.C. 1997. Antecedentes bio y cronoestratigráficos de la Formación Coya Machalí - Abanico, entre los ríos Maipo y Teno (33°55' y 35°10'S), Cordillera Principal, Chile Central. 8° Congreso Geológico Chileno (Antofagasta), Actas 1: 465-469.
14. Chiaramonte, L., Ramos, V.A. and Araujo, M. 2000. Estructura y sismotectónica del anticlinal Barrancas, Cuenca Cuyana, provincia de Mendoza. Revista Asociación Geológica Argentina 55: 309-336.
15. Cornejo, P. and Mahood, G. 1997. Seeing past the effects of re-equilibration to reconstruct magmatic gradients in plutons: La Gloria pluton, central Chile. Contributions to Mineralogy and Petrology 127: 159-175.
16. Cortés, J.M., Vinciguerra, P., Yamín, M. and Pasini, M.M. 1999. Tectónica cuaternaria de la region andina del Nuevo Cuyo (28°-38°LS). In Caminos, R. (ed.) Geología Argentina, Subsecretaría de Minería de la Nación, Servicio Geológico Minero Argentino, Anales 29: 760-778.
17. Darwin, C.R. 1845. Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. Beagle round the world, under the Command of Capt. Fitz Roy, R.N. John Murray 2d. edition, 528 p., London.
18. Darwin, C.R. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co., 280 p., London.
19. Deckart, K., Clark, A., Aguilar, C., Vargas R., Bertens, A., Mortensen, J. and Fanning, M. 2005. Magmatic and hydrothermal chronology of the giant Río Blanco porphyry copper deposit, Central Chile: Implications of an integrated U-Pb and 40Ar/39Ar database. Economic Geology 100: 905-934.
20. Farías, M., Charrier, R., Carretier, S., Martinod, J., Fock, A., Campbell, D., Cáceres, J. and Comte, D. 2008. Late Miocene high and rapid surface uplift and its erosional response in the Andes of central Chile (33°-35°S). Tectonics 27, TC1005 doi:10.1029/2006TC002046.
21. Flynn, J.J., Wyss, A.R., Charrier, R. and Swisher, C.C. 1995. An early anthropoid skull from the Chilean Andes. Nature 373: 603-607.
22. Fock, A. 2005. Cronología y tectónica de la exhumación en el Neógeno de los Andes de Chile central entre los 33° y los 34° S. Memoria para optar al título de Geólogo, Departamento de Geología, 179 p., Universidad de Chile, Santiago.
23. Gana, P. and Wall, R. 1997. Geocronología Ar/ Ar y K/Ar del límite Cretácico-Terciario en el borde oriental de la cuenca de Santiago, Chile Central (33°-33°30'). 7° Congreso Geológico Chileno (Antofagasta), Actas 2: 1280-1284.
24. Giambiagi, L. 1999. Interpretación tectónica de los depósitos neógenos de la cuenca de antepaís del Alto Tunuyán, en la región del río Palomares, Cordillera Principal de Mendoza. Revista Asociación Geológica Argentina 54: 361-374.
25. Giambiagi, L. and Ramos, V.A. 2002. Structural evolution of the Andes between 33º30' and 33º45'S, above the transition zone between the flat and normal subduction segment, Argentina and Chile. Journal of South American Earth Sciences 15: 99-114.
26. Giambiagi, L., Tunik, M. and Ghiglione, M. 2001. Cenozoic tectonic evolution of the Alto Tunuyán foreland basin above the transition zone between the flat and normal subduction segment (33°30'-34°S), western Argentina. Journal of South American Earth Sciences 14: 707-724.
27. Giambiagi, L.B., Ramos, V.A., Godoy, E. and Alvarez, P.P. 2002. Deformational history of the Andes, between 33° and 34° South Latitude, Chile and Argentina. 5° International Symposium on Andean Geodynamics: 247-250.
28. Giambiagi, L.B., Alvarez, P.P., Godoy, E. and Ramos, V.A. 2003. The control of pre-existing extensional structures in the evolution of the southern sector of the Aconcagua fold and thrust belt. Tectonophysics 369: 1-19.
29. Godoy, E., 1993. El Caloviano del Estero Yeguas Muertas, Río Yeso del Maipo, Chile: implicancias tectónicas y paleogeográficas. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza), Actas 2: 104-107.
30. Godoy, E., Yañez, G. and Vera, E. 1996. Late Miocene volcanic arc thickening in the Chilean Central Andes: Seismic and gravity constraints. 3° International Symposium on Andean Geodynamics, Actas 1: 43-46.
31. Godoy, E., Yañez, G. and Vera, E. 1999. Inversion of an Oligocene volcano-tectonic basin and uplift of its superimposed Miocene magmatic arc, Chilean Central Andes: first seismic and gravity evidence. Tectonophysics 306: 217-326.
32. Hildreth, W. and Moorbath, S. 1988. Crustal contributions to arc magmatism in the Andes of Central Chile. Contributions to Mineralogy and Petrology 98: 455-489.
33. Irigoyen, M.V., Buchan, K.L. and Brown, R.L. 2000. Magnetostratigraphy of Neogene Andean foreland-basin strata, lat 33°S, Mendoza Province, Argentina. Geological Society of America Bulletin 112: 803-816.
34. Isacks, B.L., Jordan, T.E., Allmendinger, R.W. and Ramos, V.A. 1982. La segmentación tectónica de los Andes Centrales y su relación con la geometría de la Placa de Nazca subductada. 5° Congreso Latinoamericano de Geología (Buenos Aires), Actas 3: 587-606.
35. Jordan, T.E., Isacks, B.L., Allmendinger, R.W., Brewer, J.A., Ramos, V.A. and Ando, C.J. 1983. Andean tectonics related to geometry of subducted Nazca plate. Geological Society of America Bulletin 94: 341-361.
36. Jordan, T.E., Burns, W.M., Veiga, R., Pangaro, F., Copeland, P., Kelley, S. and Mpodozis, C. 2001. Extension and basin formation in the southern Andes caused by increases convergence rate: a Mid-Cenozoic trigger for the Andes. Tectonics 20: 308-324
37. Kay, S.M., Godoy, E. and Kurtz, A. 2005. Episodic arc migration, crustal thickening, subduction erosion, and magmatism in the southcentral- Andes. Geological Society of America Bulletin 117: 67-88.
38. Kurtz, A., Kay, S.M., Charrier, R. and Farrar, E. 1997. Geochronology of Miocene plutons and Andean uplift history in the El Teniente region, Central Chile (34°-35°S). Revista Geológica de Chile 24: 75-90.
39. Maksaev, V., Munizaga, F., McWilliams, M., Fanning, M., Mathur, R., Ruiz, J. and Zentilli, M. 2004. New chronology for El Teniente, Chilean Andes, from U/Pb, 40Ar/39Ar, Re/Os and fission-track dating: Implications for the evolution of a supergiant porphyry Cu-Mo deposit. In Sillitoe, R.H., Perlló, J. and Vidal, C.E. (eds.) Andean Metallogeny: New discoveries, Concepts, Update. Society of Economic Geologists, Special Publication 11: 15-54.
40. Nystrom, J.O., Parada, M.A. and Vergara, M. 1993.Sr-Nd isotope composition of Cretaceous to Miocene volcanic rocks in Central Chile: a trend towards a MORB signature and a reversal with time. 2° International Symposium on Andean Geodynamics, Actas: 411-414, Paris.
41. Pardo, M., Comte, D. and Monfret, T. 2002. Seismotectonic and stress distribution in the Central Chile subduction zone. Journal of South American Earth Sciences 15: 11-22, doi: 10.1016/S0895-9811(02)00003-2.
42. Pardo Casas, F. and Molnar, P. 1987. Relative motion of the Nazca (Farallon) and South American Plates since Late Cretaceous time. Tectonics 6: 233-248.
43. Polanski, J. 1957. Prolegómeno a la estratigrafía y tectónica del Terciario de la Depresión Intermontánea del Alto Tunuyán. Universidad de Buenos Aires, Facultad de Ciencias Físicas y Naturales, Contribuciones científicas, Serie Geología 1(2): 95-139, Buenos Aires.
44. Polanski, J. 1964. Descripción geológica de la Hoja 25 a-b - Volcán de San José, provincia de Mendoza, Dirección Nacional de Geología y Minería, Boletín 98, 92 p., Buenos Aires.
45. Ramos, V.A. 1988. The tectonics of the Central Andes (30°-33°S latitude). In Clark, S., Burchfiel, D. and Suppe, D. (eds.) Proceses in continental lithospheric deformation. Geological Society of America, Special Paper 218: 31-54.
46. Ramos, V.A., Cegarra, M.I. and Cristallini, E.1996a. Cenozoic Tectonic of the High Andes of West-Central Argentina (30 - 36 S Latitude). Tectonophysics 259: 185-200.
47. Ramos, V.A., Godoy, E., Godoy, V. and Pángaro, F. 1996b. Evolución tectónica de la Cordillera Principal argentino-chilena a la latitud del Paso de Piuquenes (33°30'S). 13° Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos (Buenos Aires), Actas 2: 337-352.
48. Ramos, V.A., Alvarez, P.P., Aguirre Urreta, M.B. and Godoy, E. 1997. La Cordillera Principal a la latitud del paso Nieves Negras (33°50'S), Chile-Argentina. 8° Congreso Geológico Chileno, (Antofagasta) Actas 3: 1704-1708.
49. Ramos, V.A., Aguirre Urreta, M.B., Alvarez, P.P., Coluccia, A., Giambiagi, L., Pérez, D., Tunik, M. and Vujovich, G. 2000. Descripción de la Hoja Geológica Cerro Tupungato (1: 250.000), Subsecretaría de Minería de la Nación, Dirección Nacional del Servicio Geológico, (unpublished), 144 p., Buenos Aires
50. Rivano, S., Godoy, E., Vergara, M. and Villaroel, R. 1990. Redefinición de la Formación Farellones en la Cordillera de los Andes de Chile Central (32°-34°S). Revista Geológica de Chile 17: 205-214.
51. Stern, Ch., Moreno, H., López-Escobar L, Clavero, J., Lara, L., Naranjo, J.A., Parada, M.A. and Skewes, M. 2007. Chilean volcanoes. In Moreno, T. and Gibbons, W. (eds.) The Geology of Chile. The Geological Society, 148-179, London.
52. Thiele, R. 1980. Hoja Santiago, Instituto de Investigaciones Geológicas, Carta Geológica de Chile 39, 51 p., Santiago.
53. Tunik, M.A. 2003. Interpretación paleoambiental de los depósitos de la Formación Saldeño (Cretácico superior), en la alta Cordillera de Mendoza. Revista de la Asociación Geológica Argentina 58: 417-433.
54. Tunik, M.A, Concheyro, A., Ottone, G and Aguirre-Urreta, M.B. 2004. Paleontología de la Formación Saldeño (Maastrichtiano), Alta Cordillera de Mendoza, Argentina. Ameghiniana 41: 143-160.
55. Uliana, M.A., Biddle, K. and Cerdán, J. 1989. Mesozoic extension and the formation of Argentine sedimentary basins. In Tankard, A. and Balkwill, H.R. (eds.) Extensional tectonics and stratigraphy of the North Atlantic Margins. American Association of Petroleum Geologists, Memoir 46: 599-614.
56. Vergara, M., Charrier, R., Munizaga, F., Rivano, S., Sepúlveda, P., Thiele, R. and Drake, R.E. 1988. Miocene volcanism in the central Chilean Andes (31°30'S34°35'S). Journal of South American Earth Sciences 1: 199-209.
57. Vergara, M., Morata, D., Villarroel, R., Nyström, J. and Aguirre, L. 1999. Ar/Ar ages, very lowgrade metamorphism and geochemistry of the volcanic rocks from "Cerro El Abanico", Santiago Andean Cordillera (33°30'S-70°30'- 70°25'W). 4° International Symposium on Andean Geodynamics, (Göt-tingen), Actas: 785-788.
58. Wyss, A.R., Norell, M.A., Flynn, J.J., Novacek, M. J., Charrier, R., McKenna, M.C., Frassinetti, D., Salinas, P. and Meng, J. 1990. A new early Tertiary mamal fauna from central Chile: implications for stratigraphy and tectonics. Journal of Vertebrate Paleontology 10: 518-522.
59. Yrigoyen, M.R. 1993. Los depósitos sinorogénicos terciarios. In Ramos, V.A. (ed.) Geología y recursos naturales de la Provincia de Mendoza, 12° Congreso Geológico Argentino y 2° Congreso Exploración de Hidrocarburos, Relatorio: 123-148, Mendoza.
The first geological map of Patagonia
Eduardo O. Zappettini and José Mendía
Servicio Geológico Minero Argentino (SEGEMAR) - Av. Julio A. Roca 651 (1322) Buenos Aires Emails: ezappe@mecon.gov.ar, jmendi@mecon.gov.ar
ABSTRACT This contribution analyses the first geological map of Patagonia drawn by Darwin around 1840, and colour-painted by Darwin himself. It had remained unpublished and only a small version in black and white had been printed before. The different units mapped by Darwin are analysed from a modern perspective, and his ability to show a synthesis of the complex geological structure of Patagonia is stressed.
Keywords: Geological map; Patagonia; Patagonian Shingle; Darwin geologist.
RESUMEN: El primer mapa geológico de la Patagonia. La presente contribución analiza el primer mapa geológico de la Patagonia realizado por Darwin cerca de 1840, pintado en colores por el mismo Darwin, que ha permanecido inédito y del que sólo se conocía una versión de tamaño reducido en blanco y negro. Se analizan las diferentes unidades mapeadas por Darwin desde una perspectiva actual, destacándose su habilidad para mostrar en esa síntesis la compleja estructura de la Patagonia.
Palabras clave: Mapa geológico; Patagonia; Rodados Patagónicos; Darwin geólogo.
DARWIN AND THE VOYAGE OF HMS BEAGLE
At the time Charles Darwin set sail on board HMS Beagle on a journey that was to last two years and ended up lasting five, he was not more than an amateur naturalist that had quitted his medical courses and after that abandoned his intention of applying for a position in the Church of England, just to embrace the study of natural history. In those years Charles Lyell (1830-1833) published his theory of gradualism (the present is the key of the past) as a counterpart to catastrophism then prevailing in geological thought. During the five years (1831-1835) that his journey on board HMS Beagle lasted - of which three years and one month were spent ashore - Darwin made countless observations on geology and biology and even on the communities that he visited (Fig. 1). These altogether paved the way towards his famous theory on natural selection and the origin of species and the influence that the environment - including its geological constitution and evolution - has on the success and adaptability of species.
 Figure 1: Itinerary of HMS Beagle and landings and journeys of Charles Darwin in the interior of South America.
Figure 1: Itinerary of HMS Beagle and landings and journeys of Charles Darwin in the interior of South America.
DARWIN'S CONTRIBUTIONS TO GEOLOGICAL KNOWLEDGE
Darwin's contributions to the geological knowledge of South America were recorded in four main papers, i.e., On the connection of certain volcanic phenomena in South America (1838), On the distribution of erratic boulders and on the contemporaneous stratified deposits of South America (1841), An account of the fine dust which often falls on vessels in the Atlantic Ocean (1846a,[Read 4 June 1845), On the Geology of the Falkland Islands (1846b), and finally the book Geological Observations on South America (1846c) reprinted in 1876 as part of his Geological observations on the volcanic islands and parts of the South America visited during the voyage of H.M.S. Beagle. On the other hand, a first unpublished essay of his geological observations was written during his voyage on board H.M.S. Beagle ca. 1834 which is titled Reflection on reading my geological notes. In this work Darwin put forward a theory on the geological formation of South America and included his interpretations on the history of life in the continent which, albeit not evolutionist is nevertheless sequential (cf. S. Herbert 1995). In these works there are numerous and detailed descriptions of geological units. However, figures representing his observations are few and of limited interest. Despite of this, Darwin at some stage of his life as a scientist tried to synthesize in a map the main units that were subject of his work, although such a map was never published. The Library at Cambridge University keeps an original unpublished map painted with water-colours by Darwin himself. This is the earliest known Regional Geological Map of the Patagonian region of Chile and Argentina (Fig. 2). A black and white version was published by Rhodes (1991, Fig. 3) who stated "the magnificent structure of this map" in contrast with the other Darwin's unpublished map of Patagonia which is rather simple (Herbert 1991 and 2005, plate 6). The geological observations were represented on the topography produced by (1826-1830) and published in London by John Murray. The watercolour is from the time of Darwin's voyage on the Beagle or maybe just a few years later, circa 1840. On the back of the map there is an annotation reading "p 94", which suggests that it was drawn for inclusion as an illustration to one of the volumes of the Geology of the voyage of H.M.S. Beagle (1846c), but it was never printed. The only previous regional maps are those published by d'Orbigny (1842) together with the geological sketch of the provinces of Misiones, Corrientes, Entre Ríos, Santa Fe, and Buenos Aires, including northern Patagonia down to Valdés Peninsula.
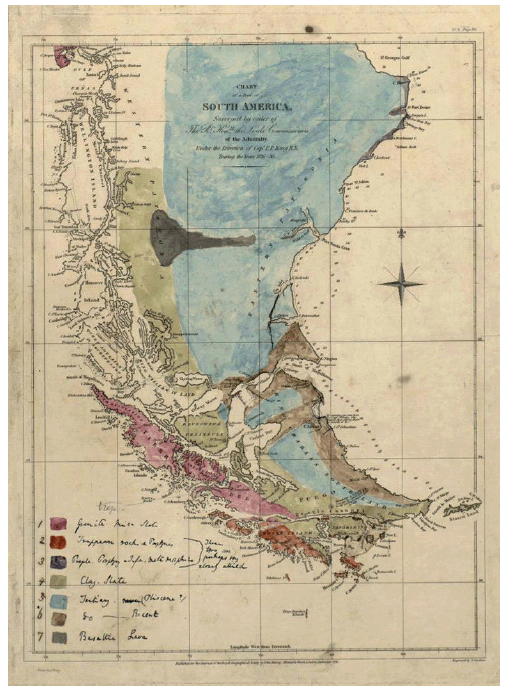 Figure 2: Geologic map of Patagonia (Darwin, circa 1840, unpublished).
Figure 2: Geologic map of Patagonia (Darwin, circa 1840, unpublished).
GEOLOGICAL MAP OF PATAGONIA
The observations by Darwin recorded on the map are mainly referred to coastal areas surveyed by HMS. Beagle (Fig. 2) and involve seven geological units:
1) Granite, Mica slate, 2) Trappean rock and porphyries, 3) Purple porphyries and infra metamorphics, 4) Clay slate, 5) Tertiary [newer - crossed out -] (Pliocene?), 6) Recent, 7) Basaltic lava.
About units 2 and 3 he states "These two are perhaps very closely allied".
UNIT 1 (Granite, Mica slate). It mainly involves the Cretaceous so-called Coast Batolith that Darwin stated as exposed in Chilean Patagonia south of the Strait of Magellan. It includes outcrops at the notheastern corner of Hoste Island and at the northwestern sector of Navarino Island. In this unit are also included the Mica slates of the southernmost end of the Chonos Archipelago in the Taitao and Tres Montes peninsulas. In his description of this unit Darwin recorded different lithologies, among which he distinguished - west of the Beagle Channel bifurcation - a complex of gneiss and granitic slates that comprise the Cordillera presently bearing his name (Eastern Metamorphic Complex of Aysen and Magallanes in Chile and Lapataia Formation in Argentina). Towards the northeast he pointed out the predominance of gneiss and granites (Coast Batolith in Chile). Between Cabo Tres Montes and the north of the Chonos Islands the predominant rock is a Mica slate with organic matter (presently Canal King Formation).
UNIT 2 (Trappean rock and porphyries). It includes intrusive and extrusive rocks exposed in Hoste and Londonderry Islands (Chile) and also Mesozoic ophiolitic rocks exposed in Wollaston Island. It comprises Paleogene monzodiorites, diorites and gabros and Mesozoic basaltic lavas of the Hardy Formation, and the Sarmiento and Tortugas ophiolitic complexes in Chile. In Wollaston Island Darwin described "greenstones" with pyrite and epidote with evidence of metamorphism, and basaltic submarine volcanic rocks. In Hardy Peninsula he recorded the pyrite-rich "trappean and basaltic rocks", trachytes with columnar jointing, and diorites rocks. Darwin's precise drawing of the contact between this unit and Unit 1 is especially remarkable when compared to modern maps.
UNIT 3 (Purple porphyries and infra metamorphics). This unit was recognized by Darwin on the coast of Santa Cruz. It comprises the volcanic rocks of the Chon Aike Formation, which in this sector represents the easternmost exposures of the Deseado Massif. Darwin described these volcanics as trachytic porphyries. The unit includes the tectonic silicified breccias assigned to the Bahía Laura Group, and exposed at Cabo Blanco. These were erroneously interpreted by Darwin as siliceous sedimentary rocks and compared them to those of the Malvinas Islands. Off-limits he describes purple to pink, and sometimes laminated, porphyric rocks, which are slightly vesicular with feldspar and quartz crystals. He stated these as the dominant rocks in between Port Santa Elena, Camarones and Malaspina up to the "Paps of Pineda" (sic). At Puerto Deseado he records the presence of porphyries up to 40 km west of the coast, covered by Tertiary deposits. He indicates that the composition is similar to that of those described further north and points out the presence of local chert beds.
UNIT 4 (Clay slate). According to Darwin's description, it comprises siltstones, feldspar volcanics, graywackes, scarce black limestones with Lower Cretaceous fossil remains, and metamorphic schists. These are the core of the mountain ranges of southern Tierra del Fuego and Navarino Island. He describes slates exposed along the Beagle Channel between its bifurcation and Lemaire Strait, constituting Navarino Island to the south and the eastern sector of Hoste Island and Hardy Peninsula. North of the Beagle Channel this unit runs in a northwestern direction towards Brunswick Peninsula on both coasts of Admiralty Strait, continuing north along the eastern flank of the Cordillera. Darwin collected fossils from the limestone beds he recorded. These indicate a Cretaceous age. Darwin stressed the change in the direction of the Cordillera, that north of the Strait of Magellan runs with a N-S orientation changing south of it to a E-W orientation. This unit includes the present Lemaire and Yaghan Formations in Argentina and Cerro Toro, Yaghan and Punta Barrosa Formations in Chile, spanning the Late Jurassic-Cretaceous (see Olivero et al. 2009). Again, we must stress the precision with which Darwin drew the contact between units 2 and 4 in Tierra del Fuego.
UNIT 5 (Tertiary, Pliocene?). Darwin includes here the Tertiary units exposed in extra-Andean Patagonia right across to the coast of Santa Cruz. He observes that these overlie porphyries of Unit 3. In the map, he represents this unit in the hills of northern Tierra del Fuego and in the northern areas of Brunswick Peninsula. Darwin's observations on this unit include the identification of several faunal assemblages with abundant Turritella, Pecten and Ostrea. He called this complex the "Patagonian Tertiary Formation" (see Parra and Griffin 2009, Casadío and Griffin 2009). From a lithologic point of view he recognized the presence of beds with pumiceous siltstone with abundant gypsum, sandstones, and claystones with calcareous concretions. At the headwaters of the Santa Cruz River he observed that the sedimentary rocks are covered by basaltic lavas of unit 7.
UNIT 6 (Recent). This unit represents Quaternary deposits that Darwin recognized at the southern end of Santa Cruz between the Río Gallegos and Cabo Vírgenes, in the northeastern coast of Tierra el Fuego, along both coasts of the Strait of Magellan in its northeastern sector, and in the sector between Useless Bay and Bahía San Sebastián. He includes here the "Rodados Patagónicos", which he called "Gravel Formation of Patagonia". In his description he indicates that not all the gravel beds are from recent times, particularly those of higher tablelands (see discussion in Martínez et al. 2009). He recorded interesting observations on the origin and transport mode of the gravels, noting the difficulty in explaining the distribution covering the entire area from the Andes to the coast. He did point out, however, that the present distribution of the gravels was due to marine action.
UNIT 7 (Basaltic lava). It represents the basaltic exposures along the middle and upper course of the Río Santa Cruz and the Meseta La Siberia. Darwin placed these outcrops as contemporaneous of the higher sections of the Tertiary formation. In fact, the basalts are of a Pliocene-Quaternary age (see Streling and Malagnino 2009) and include the units presently called Meseta de las Vizcachas Basalt and La Siberia Formation. In his observations he states that the outcrops predominate on the Northern side of the Río Santa Cruz valley and that they reach the foothills of the Andes. He recorded that the basalts were olivinic and that columnar disjunction was common. He identifies at least three overlying beds. Acknowledging the great extension covered by the exposures he concludes that "This great deluge of lava is worthy, in its dimensions, of the great continent to which it belongs. The aggregate streams have flowed from the Cordillera to a distance (unparalleled, I believe, in any case yet known) of about 100 geographical miles" (Darwin 1846, p. 116)
CONCLUSIONS
The brief description presented herein shows one of Darwin's less known facets, i.e., his ability to represent his geological observations in a map. This contrasts Stoddart (1995) who emphasized, when analyzing the geological results of Darwin's voyage on the Beagle, "the contrast between Darwin's verbal facility in describing landscapes and evoking mood, and his general inability to translate his images into visual representations". Lastly, it should be stressed that if it had been published when originally planned, it would have constituted a singular initial contribution to the geological cartography of the South American continent.
WORKS CITED IN THE TEXT
1. Casadío, S. and Griffing, M. 2009. Sedimentology and Paleontology of a Miocene marine succession frst noticed by Darwin at Puerto Deseado (Port Desire). Revista de la Asociación Geológica Argentina 64 (1): 83-89.
2. Darwin, C. 1838. On the connexion of certain volcanic phaenomena, and on the formation of mountain-chains and volcanos, as the effects of continental elevations. Proceedings of the Geological Society 2: 654-660, London.
3. Darwin, C. circa 1840. Geological Map of Patagonia image from the Cambridge University Library (Manuscript MS.DAR.44:13). Available at the Keeping time exhibition web-site: http://www.lib.cam.ac.uk/exhibitions/Keeping_Time/images/large/darwin.jpeg
4. Darwin, C. 1841. On the distribution of erratic boulders and on the contemporaneous unstratified deposits of South America. Proceedings of the Geological Society 3(2): 425-430, London.
5. Darwin, C. 1845. Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. Beagle round the world, under the Command of Capt. Fitz Roy, R.N. John Murray, 2d. edition, 519 p., London.
6. Darwin, C. 1846a. An account of the fine dust which often falls on vessels in the Atlantic ocean. Quarterly Journal of the Geological Society 2: 26-30, London.
7. Darwin, C. 1846b. On the geology of the Falkland Islands. Quarterly Journal of the Geological Society 2: 267-279, London.
8. Darwin, C. 1846c. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 280 p., London.
9. Darwin, C. 1876. Geological observations on the volcanic islands and parts of South America visited during the voyage of H.M.S. 'Beagle'. 2d ed. Smith Elder & Co. 648 p., London.
10. D'Orbigny, A. 1842. Voyage dans l'Amérique Méridionale. Bertrand ed., 3(Géologie), 290 p., Paris.
11. Herbert, S. 1991. Darwin as a prospective geological author. British Journal for the History of Science 24: 159-192.
12. Herbert, S. 1995. From Charles Darwin's Portfolio: An early essay on South American Geology and species. Earth Sciences History 14(1): 23-36.
13. Herbert, S. 2005. Charles Darwin, Geologist. Cornell University Press. 485 p., Ithaca.
14. Lyell, C. 1830-1833. Principles of Geology. John Murray 1(1830) 511 p.; 2(1832) 330 p.; 3(1833) 109 p., London.
15. Martínez, O.A., Rabassa, J. and Coronato, A. 2009. Charles Darwin and the first scientific observations on the Patagonian Shingle Formation ("Rodados Patagónicos"). Revista de la Asociación Geológica Argentina 64(1): 90-100.
16. Parras, A. and Griffin, M. 2009. Darwin's Great Tertiary Patagonian Formation at the mouth of the Río Santa Cruz: a reappraisal. Revista de la Asociación Geológica Argentina 64(1): 70-82.
17. Olivero, E.O., Medina, F.A., and López C., M.I. 2009. The stratigraphy of Cretaceous mudstones in the Eastern Fuegian Andes: New data from body and trace fossils. Revista de la Asociación Geológica Argentina 64(1): 60-69.
18. Rhodes, F.H.T. 1991. Darwin's search for a theroy of the earth: symmetry, simplicity and speculation. British Journal for the History of Science 24: 193-229.
19. Stoddart, D. 1995. Darwin and the Seeing Eye: Iconography and Meaning in the Beagle Years. Earth Sciences History 14(1): 3-22.
20. Strelin, J. and Malagnino, E. 2009. Charles Darwin and the oldest glacial events/episodes in Patagonia: the erratic blocks of the Río Santa Cruz Valley. Revista de la Asociación Geológica Argentina 64(1): 101-108.
Eduardo B. Olivero1, Francisco A. Medina2, and María I. López C.1
1 Centro Austral de Investigaciones Científicas (CADIC-CONICET), B.A. Houssay 200, 9410 Ushuaia, Tierra del Fuego. Email: emolivero@gmail.com 2 Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, UBA, Intendente Güiraldes 2160, Ciudad Universitaria, C1428EGA Buenos Aires.
ABSTRACT: The stratigraphy of Cretaceous marine mudstones in the Fuegian Andes, roughly equivalent to Charles Darwin's clay-slate formation, remains a still unsolved problem. Previous records of Albian, Turonian-Coniacian, and Santonian-Campanian bivalves are combined with new findings of the Late Albian inoceramid Inoceramus anglicus Woods, and the Maastrichtian ammonites Diplomoceras sp., Anagaudryceras sp., Maorites densicostatus (Kilian and Reboul), Maorites sp., and Pachydiscus (Neodesmoceras) sp. to further constrain the Cretaceous stratigraphy of the eastern Fuegian Andes. In addition, new records of distinctive trace fossils and ichnofabric are meaningful for stratigraphic division and delineation of paleoenvironmental settings in these Cretaceous mudstones. The Lower Cretaceous ichnoassemblage of Chondrites targioni (Brongniart) and Zoophycos isp. is consistent with the inferred slope-volcaniclastic apron settings of the Yahgan Formation; Nereites missouriensis (Weller) reflects distal basin plain depositional settings for the Beauvoir Formation. In the Upper Cretaceous, the "Estratos de Buen Suceso" record the earliest extensively bioturbated horizons, reflecting prolonged well-oxygenated bottom conditions. In the Bahía Thetis Formation, organic-rich, channel margin or distal basin slaty mudstones record the last occurrence of inoceramid bivalves in the Austral Basin; the generalized absence of trace fossils is consistent with dysoxic bottom conditions. The thoroughly bioturbated Policarpo Formation, records a marked change in paleoceanographic conditions. The strong contrast in the intensity of bioturbation between the Upper Campanian-Maastrichtian Bahía Thetis Formation, almost devoid of trace fossils, and the highly bioturbated Maastrichtian-Danian Policarpo Formation reflects a change from dysoxic-anoxic to well ventilated conditions, probably associated with a cooling trend of bottom waters in the austral deep oceans.
Keywords: Clay-slate formation; Inoceramid; Ammonites; Cretaceous; Trace fossils; Fuegian Andes.
RESUMEN: Estratigrafía de las fangolitas del Cretácico en los Andes Fueguinos orientales: Nuevos datos de cuerpos y trazas fósiles. En los Andes Fueguinos la estratigrafía de las fangolitas del Cretácico marino (clay-slate formation de Charles Darwin), constituye un problema no enteramente resuelto. Datos previos de bivalvos del Albiano, Turoniano-Coniaciano, y Santoniano-Campaniano, junto con nuevos hallazgos de inoceramidos del Albiano tardío, Inoceramus anglicus Woods, y amonites maastrichianos, Diplomoceras sp., Anagaudryceras sp., Maorites densicostatus (Kilian y Reboul), Maorites sp., y Pachydiscus (Neodesmoceras) sp. se utilizan para obtener una mejor resolución estratigráfica. Además, nuevas asociaciones distintivas de trazas fósiles permiten ajustar la interpretación paleoambiental. La asociación de Chondrites targioni (Brongniart) y Zoophycos isp. del Cretácico inferior es consistente con el modelo inferido de rampa volcaniclástica para la Formación Yahgan. En la Formación Beauvoir la presencia de Nereites missouriensis (Weller) refleja condiciones de depositación en planicies distales de la cuenca. En el Cretácico superior, los "Estratos de Buen Suceso" registran los primeros horizontes bioturbados asociados a fondos con prolongados periodos de condiciones de buena oxigenación. En la Formación Bahía Thetis, fangolitas pizarreñas ricas en materia orgánica depositadas en márgenes de canales o en planicies distales, registran la última aparición de bivalvos inocerámidos en la Cuenca Austral. La ausencia generalizada de trazas fósiles es consistente con condiciones de fondos anóxicos. La Formación Policarpo, densamente bioturbada, marca un profundo cambio en las condiciones paleoceanográficas. El fuerte contraste en la intensidad de la bioturbación entre las Formaciones Bahia Thetis, Campaniano-Maastrichtiano, con escasa presencia de trazas fósiles y Policarpo, Maastrichiano-Daniano, altamente bioturbada, refleja el cambio de condiciones de fondo disóxico-anóxico a óxico, probablemente asociado al comienzo del enfriamiento de las aguas profundas del océano austral.
Palabras clave: Clay-slate formation; Inoceramidos; Amonites; Trazas fósiles; Cretácico; Andes Fueguinos.
INTRODUCTION
The stratigraphic division of Cretaceous marine mudstones in the Fuegian Andes (Fig. 1) is a major and still unsolved problem. These mudstones correspond roughly to the clay-slate formation of Charles Darwin (1846); a very extensive formation of folded, low-grade metamorphic rocks with scarce marine fossils. In the course of the voyage of HMS Beagle Darwin noted that "The great clayslate formation of Tierra del Fuego being Cretaceous, is certainly a very interesting fact,--whether we consider the appearance of the country, which, without the evidence afforded by the fossils, would form the analogy of most known districts, probably have been considered as belonging to the Palaeozoic series,…"(Darwin 1846, p. 152).
 Figure 1: Locality map, tectonic settings, and distribution of Lower and Upper Cretaceous rocks in the Rocas Verdes Marginal basin and Austral/Malvinas Foreland basins.
Figure 1: Locality map, tectonic settings, and distribution of Lower and Upper Cretaceous rocks in the Rocas Verdes Marginal basin and Austral/Malvinas Foreland basins.
The evidence mentioned by Darwin included a few fossils from Mount Tarn, on the western side of the Strait of Magellan, Brunswick Peninsula in Chile (Fig. 2d), particularly the ammonites described by Forbes (in Darwin 1846) as "Ancyloceras simplex" d'Orbigny, from Mount Tarn (Fig. 2f), and "Hamites elatior" Sowerby, from the coastal area nearby Port Famine. We now know that these fossils are not Albian hamitids; the "A. simplex" is a Maastrichtian kossmaticeratid, probably a deformed fragment of Maorites, and "H. elatior" is a Diplomoceras, also of Maastrichtian age (Spath 1953, Lahsen and Charrier 1972). However, the correction in the age of these ammonite-bearing strata does not change the fact that in the Fuegian Andes, Albian to Maastrichtian fine-grained rocks have quite similar lithological and structural features and consequently they are very difficult to distinguish in the field (Fig. 2).
 Figure 2: Structure, lithology, and paleontology of Cretaceous rocks. a) Thin-bedded turbidites, Yahgan Formation, Lower Cretaceous, Beagle Channel close to Ushuaia. b) Black mudstones, Beauvoir Formation, Lower Cretaceous (Albian), Knokeke Hill area. c) Black mudstones and interbedded light-gray tuffs, Estratos de Buen Suceso, Upper Cretaceous (Santonian-Lower Campanian). d) Upper Cretaceous (Maastrichtian) interbedded siltstones and finegrained sandstones, Mount Tarn, Brunswick Peninsula, Chile. e) Highly folded, black mudstones and fine-grained sandstones, Bahía Thetis Formation, Thetis Bay. f) Crushed ?Maorites sp. from Mount Tarn. This is the original specimen, housed at the Natural History Museum, London (B.M. No. 2612), collected by Darwin and described by Forbes as "Ancyloceras simplex" d'Orbigny. Scale bar: 1 cm.
Figure 2: Structure, lithology, and paleontology of Cretaceous rocks. a) Thin-bedded turbidites, Yahgan Formation, Lower Cretaceous, Beagle Channel close to Ushuaia. b) Black mudstones, Beauvoir Formation, Lower Cretaceous (Albian), Knokeke Hill area. c) Black mudstones and interbedded light-gray tuffs, Estratos de Buen Suceso, Upper Cretaceous (Santonian-Lower Campanian). d) Upper Cretaceous (Maastrichtian) interbedded siltstones and finegrained sandstones, Mount Tarn, Brunswick Peninsula, Chile. e) Highly folded, black mudstones and fine-grained sandstones, Bahía Thetis Formation, Thetis Bay. f) Crushed ?Maorites sp. from Mount Tarn. This is the original specimen, housed at the Natural History Museum, London (B.M. No. 2612), collected by Darwin and described by Forbes as "Ancyloceras simplex" d'Orbigny. Scale bar: 1 cm.
In this study, previous records of Albian, Turonian-Coniacian, and Santonian- Campanian bivalves, mostly inoceramids (Olivero and Martinioni 1996a, Olivero and Medina 2001) are combined with new findings of latest Albian inoceramids and Maastrichtian ammonites to further constrain the Cretaceous stratigraphy of the eastern Fuegian Andes. In addition, new records of characteristic trace fossil assemblages with distinctive ichnofabrics, restricted to particular sedimentary successions of the Yahgan Formation, Beauvoir Formation, Bahía Thetis Formation, Policarpo Formation, and "Estratos de Buen Suceso", constitute meaningful data for the stratigraphic division of the relatively homogeneous, mudstone-rich Cretaceous deposits.
STRATIGRAPHIC FRAMEWORK
The stratigraphy of southernmost South America has been controlled since the Mesozoic by a series of contrasting tectonic regimes, represented by Late Jurassic-Early Cretaceous extension, Late Cretaceous-Paleogene compression, and latest Paleogene-Neogene and present day strike-parallel transcurrent motion (Katz 1972, Dalziel et al. 1974, Suárez et al.1985, Kohn et al. 1995, To-rres Carbonell et al. 2008). Accordingly, Cretaceous rocks in the Fuegian Andes are distributed in three tectonostratigraphic domains: 1) the magmatic arc, located along the axis of the southern archipelago, includes Lower Cretaceous andesitic (Hardy Formation) and Upper Cretaceous plutonic rocks (Beagle Channel Plutonic Group); 2) the Rocas Verdes Marginal Basin includes Lower Cretaceous, low-grade metamorphic rocks, mostly slates and sandstones of the Yahgan and Beauvoir formations, and Upper Jurassic-Lower Cretaceous ophiolitic rocks; and 3) the Austral and Malvinas foreland basins, separated in the northern area by the Dungeness or Rio Chico Arch, include Upper Cretaceous rocks dominated by slaty mudstones (Fig. 1). According to a recent review (see Olivero and Malumián 2008, and the bibliography cited therein), the Yahgan Formation represents a volcaniclastic apron of mudstones and deep-marine andesiterich turbidites and the Beauvoir Formation records basin plain and slope mudstones. Both formations reflect Late Jurassic-Early Cretaceous extension and the consequent origination of the Rocas Verdes Marginal Basin. The earliest Late Cretaceous ductile deformation, isoclinal folding, and low-grade regional metamorphism of these rocks indicate a compressional tectonic regime that resulted in the closure of the marginal basin. The Fuegian Andes were uplifted by the Late Campanian; subsequent propagation of the compressional deformation and subsidence by tectonic loading along the northern orogenic margin resulted in the formation of the Austral and Malvinas foreland basins. The Turonian-Lower Campanian "Estratos de Buen Suceso" represent the final stages of the closure of the marginal basin and/or the beginning of the foreland basins. Conglomerate beds in the Bahía Thetis Formation bear foliated clasts of Andean-derived rocks and thus they record uplifting and subaerial erosion of the Fuegian Andes. The youngest Cretaceous rocks are included in the Policarpo Formation, which includes the Maastrichtian/Danian boundary at its top (Olivero et al. 2003). Figure 3 summarizes the paleontological information, including partly new body and trace fossil data, used in this study to differentiate particular horizons within these lithologically homogeneous stratigraphic units. The stratigraphic implications of these data are described for each formation in the next section.

 Figure 3: Stratigraphic distribution of characteristic body and trace fossils in the Cretaceous of the Fuegian Andes (explanation in the text).
Figure 3: Stratigraphic distribution of characteristic body and trace fossils in the Cretaceous of the Fuegian Andes (explanation in the text).
LOWER CRETACEOUS
On the main island of Tierra del Fuego, Darwin (1846) recorded only Cenozoic fossils in the vicinity of Cape San Sebastian, including Nothofagus leaves and mollusks. The first fossils from the region, which are now accepted as uppermost Jurassic or Cretaceous in age, were recorded in 1839 near Nassau Bay (Navarino Island) by James Dwight Dana, a geologist of Charles Wilkes` United States Exploring Expedition (cf. Andersson 1906, Kranck 1932). The first Cretaceous fossils from the main island of Tierra del Fuego were found in 1882 nearby Ushuaia by Lovisato. They include trace fossils (and calcareous algae?) described by Richter (1925), together with Cretaceous radiolaria from the Staten and New Year islands (also collected by Lovisato) and several fossils, including belemnites from the Hito XIX area, collected by the Expedition of University of Buenos Aires to Tierra del Fuego (Doello Jurado 1922).
Yahgan Formation
The Yahgan Formation was established by Kranck (1932) for the slates, slaty mudstones, and andesite-rich greywackes exposed in Mount Olivia and Ushuaia. The Formation is well exposed along the northern margin of the Beagle Channel, from Ushuaia to Bahía Sloggett (Fig. 1); and in Hoste, Navarino, Nueva, Lennox, and Picton islands (Katz and Watters 1966, Dott et al. 1977, Winn 1978, Suárez et al. 1985). Dominant sedimentary facies are: a) black mudstones, fine-grained, thin-bedded turbidites (Fig. 2a) and tuff; b) classical turbidites; and c) massive to graded sandstones. Petrographic composition of the sandstones is uniform and dominated by lithic andesitic fragments and plagioclase, suggesting derivation from the volcanic arc, located to the South in the Fuegian Archipelago (Winn 1978, Suárez et al. 1985, Olivero and Martinioni 1996b). This is well documented with paleocurrent data on the island of South Georgia, now displaced along the North Scotia Ridge to the east (Dalziel et al. 1974, Macdonald 1986). Body and trace fossils are very scarce in the Yahgan Formation (Fig. 3). On Navarino Island Tithonian-Neocomian ammonites and belemnites (Aguirre-Urreta and Suárez 1985); Aptian-Albian corals, gastropods, and bivalves (Dott et al. 1977); and indeterminate inoceramids (Katz and Watters 1966) were reported. Microfossils, particularly radiolaria, are apparently common, but not well preserved due to the strong tectonic deformation of the rocks (Kranck 1932, Olivero and Martinioni 1996b). The trace fossil Chondrites targioni (Brongniart) and the presumed fossil calcareous algae "Lithocaulon antarcticum" Bornemann were described by Richter (1925) from the vicinity of Ushuaia. Biostratigraphically diagnostic Late Albian fossils are known from Moat, where Actinoceramus concentricus (Parkinson) and Inoceramus carsoni Mc Coy were recorded (Olivero and Martinioni 1996a). Fine-grained turbidites and mudstones nearby Ushuaia and adjoining Mount Martial bear scarce and ill-preserved belemnites. Trace fossils are rare and restricted to a few horizons; Chondrites targioni (Fig. 4b) and Zoophycos isp. are the most common trace fossils and are locally abundant in particular beds. A few localities, e.g. on the coast of the Beagle Channel just east from Ushuaia, bear a more diversified ichnoassemblage, including Chondrites targioni, Helminthopsis tenuis, Stelloglyphus isp., Ophiomorpha cf. annulata, and Zoophycos isp. (Fig. 3). A similar trace fossil assemblage, with the addition of Phycosiphon incertum, was recorded in the Yahgan Formation by Winn (1978).
 Figure 4: Typical trace fossils and ichnofabrics of Cretaceous rocks in the Fuegian Andes. a) Nereites missouriensis (Weller) from Albian beds of the Beauvoir Formation near Knokeke Hill, CADIC PI 88. b) Dense ichnofabric of Chondrites targioni (Brongniart) and associated Ophiomorpha cf. annulata (Op), Lower Cretaceous Yahgan Formation, Beagle Channel near Ushuaia, field specimens. c) Localized horizons with a relatively dense ichnofabric in mudstones and isolated specimens of Rhizocorallium isp. (Rh), Estratos de Buen Suceso, Santonian-Lower Campanian, Buen Suceso Bay, field specimens. d) Typical ichnofabric affecting thick packages in the Maastrichtian Policarpo Formation, Zoophycos isp. (Zo) and Schaubcylindrichnus ("Terebellina") isp. (Sc) are the only recognizable structures in the intensely mottled background, field specimens. Scale bar: 1 cm.
Figure 4: Typical trace fossils and ichnofabrics of Cretaceous rocks in the Fuegian Andes. a) Nereites missouriensis (Weller) from Albian beds of the Beauvoir Formation near Knokeke Hill, CADIC PI 88. b) Dense ichnofabric of Chondrites targioni (Brongniart) and associated Ophiomorpha cf. annulata (Op), Lower Cretaceous Yahgan Formation, Beagle Channel near Ushuaia, field specimens. c) Localized horizons with a relatively dense ichnofabric in mudstones and isolated specimens of Rhizocorallium isp. (Rh), Estratos de Buen Suceso, Santonian-Lower Campanian, Buen Suceso Bay, field specimens. d) Typical ichnofabric affecting thick packages in the Maastrichtian Policarpo Formation, Zoophycos isp. (Zo) and Schaubcylindrichnus ("Terebellina") isp. (Sc) are the only recognizable structures in the intensely mottled background, field specimens. Scale bar: 1 cm.
Beauvoir Formation
The Beauvoir Formation (Camacho 1967) consists predominantly of homogeneous black or bluish, massive to faintly laminated dark slates and gray tuffs mainly exposed in Sierra de Beauvoir, in the belt stretching from the eastern part of Lake Fagnano to the Montes Negros, just west of Good Success (Buen Suceso) Bay (Figs. 1 and 2b). Locally, the black slates are rhythmically interbedded with thin, fine-grained sandstones. Stratification is only visible in the latter beds, whereas in the massive slates the bedding is defined only by the presence of thin tuff beds. The fossiliferous marlstones cropping out near Hito XIX in the Argentinean-Chilean border are provisionally included in the Beauvoir Formation. These marlstones contain abundant specimens of the Aptian-Albian bivalves Aucellina "radiotostriata" Bonarelli, A. "andina" Feruglio (Macellari 1979), and A. striata Richter (Richter 1925). Additional fossils include serpulids, brachiopods, inoceramids, probably Inoceramus anglicus Woods or I. sutherlandi MacCoy (cf. Medina 2007) and abundant rostra of the belemnite Parahibolites fuegensis (Stolley) in interbedded sandstone beds (Richter 1925) and large, poorly preserved ammonites. The massive, dark slates of the Beauvoir Formation near Knokeke Hill and Buen Suceso Bay contain several horizons with abundant but ill-preserved inoceramids, which normally are concentrated in very thin beds. Part of this material includes the Albian Mytiloides cf. ipuanus (Wellman), of which the nominal species "Inoceramus" urius Wellman and "I. kapuus" Wellman are now considered as synonyms (see Crampton 2004). Associated, very thin beds include abundant wellpreserved, articulate shells of the Late Albian Aucellina euglypha Woods (Fig. 5b) covered by a thin pyrite coating (Olivero and Medina 2001). Rare, complete but badly deformed echinoids are associated with the latter bivalves. Near the locality of Knokeke Hill, black slates record relatively well preserved and articulated shells of Inoceramus anglicus (Fig. 5a) and delicate back-filled burrows of Nereites missouriensis (Weller) (Fig. 4a). In the subsurface, the stratigraphically equivalent mudstone-dominated rocks of the Nueva Argentina Formation and Arroyo Alfa Formation contain Early-Mid Albian and Late Albian foraminiferal assemblages, respectively (Flores et al. 1973).
 Figure 5: Cretaceous ammonites and bivalves from the Fuegian Andes. a) Inoceramus anglicus Woods, Upper Albian, Beauvoir Formation, right valve, CADIC PI 87; b) Black mudstones with pyritized shells of Aucellina euglypha Woods, Upper Albian, Beauvoir Formation, CADIC PI6 a; c) Fragment of the body chamber of Diplomoceras sp., Maastrichtian, Policarpo Formation, CADIC PI 85; d) Pachydiscus (Neodesmoceras) sp., Maastrichtian, Policarpo Formation, CADIC PI 84; e) and f) Fragments of the body chamber of crushed Maorites spp., Maastrichtian, Policarpo Formation; e) Maorites densicostatus (Kilian and Reboul) CADIC PI 86; f) Maorites sp. CADIC PI 89. Scale bar: 1 cm.
Figure 5: Cretaceous ammonites and bivalves from the Fuegian Andes. a) Inoceramus anglicus Woods, Upper Albian, Beauvoir Formation, right valve, CADIC PI 87; b) Black mudstones with pyritized shells of Aucellina euglypha Woods, Upper Albian, Beauvoir Formation, CADIC PI6 a; c) Fragment of the body chamber of Diplomoceras sp., Maastrichtian, Policarpo Formation, CADIC PI 85; d) Pachydiscus (Neodesmoceras) sp., Maastrichtian, Policarpo Formation, CADIC PI 84; e) and f) Fragments of the body chamber of crushed Maorites spp., Maastrichtian, Policarpo Formation; e) Maorites densicostatus (Kilian and Reboul) CADIC PI 86; f) Maorites sp. CADIC PI 89. Scale bar: 1 cm.
UPPER CRETACEOUS
"Estratos de Buen Suceso"
These beds include a thick sedimentary succession of folded dark gray and black, slaty mudstones, marlstones, and sandy siltstones with common quartz veins. The lower part of the succession, exposed in the southern part of the bay, is dominated by marlstones, impure limestones, and fine-grained silty sandstones. The slaty micritic limestones and marly mudstone record well-preserved, articulated shells of Tethyoceramus madagascariensis (Heinz). This species has been referred to the Middle Coniacian in Madagascar and New Zealand, and to the Upper Turonian-Lower Coniacian in Antarctica (cf. Olivero and Medina 2001). The exact age and identity of inoceramids assigned to "Inoceramus" (=Tethyoceramus) madagascariensis is now debated. Walaszczyk et.al (2004) argue that the New Zealand material is not true "I." madagascariensis and should be referred to "I." nukeus Wellman. However, Crampton (writ. comm. 2008) believes that some New Zealand material is indistinguishable from topotype T. madagascariensis (e.g., compare Crampton 1996 pl. 16M and pl. 17E, with Walaszczyk et al. 2004, Fig. 13B and fig. 14E, respectively). The Tierra del Fuego material is indistinguishable from the Antarctic specimens, and appears to differ from "I."? nukeus, which is almost equivalve and much less inflated than T. madagascariensis. The upper part of the succession, exposed along the northern margin of the bay, is dominated by dark slaty mudstones and silty sandstones, with some interbedded light gray, thin tuff beds (Fig. 2c). The slaty mudstones contain occasional, large specimens (up to 50 cm high) of complete shells of Inoceramus (Platyceramus) sp. and relatively small, well-preserved specimens of Sphenoceramus sp. The association described by Olivero and Medina (2001) was referred to the Santonian-Lower Campanian. The silty sandstones within the Santonian-Lower Campanian beds record several fully bioturbated horizons. The trace fossils Chondrites, Palaeophycus, Planolites, Rhizocorallium (Fig. 4c), and Zoophycos are recognized within a heavily bioturbated background.
Bahía Thetis Formation
The dominant slaty mudstones and sandstones cropping out in the Thetis Bay area were first recognized by Furque and Camacho (1949). Olivero et.al (2003) studied the Bahía Thetis Formation and recognized three packages, more than 250 m thick, of hard, highly deformed rocks including: 1) dark, organic-rich, laminated mudstones and tuffs with incipient cleavage; 2) resedimented conglomerates and pebbly mudstones; and 3) turbidite sandstones and slaty mudstones. The lowest organic-rich package bears abundant radiolaria and more restricted foraminifera. The latter comprise a low diversity, agglutinated assemblage typical of dysoxic environments and are characterized by Rzehakina epigona (Rzehak), R. lata Cushman and Jarvis and R. fissistomata (Grzybowski) (Caramés and Malumián 2006). In the second package, the conglomerates include large clasts of radiolarian-bearing slates and foliated rhyolites, derived from the Beauvoir-Yahgan Formations and Jurassic volcanics, respectively. In the third package, the turbidites (Fig. 2e) bear scarce, ill-preserved ammonites, including flattened fragments of Diplomoceras sp. and kossmaticeratids. Trace fossils are very rare, and only a few specimens of Stelloglyphus were recovered. The absence of trace fossils is consistent with the high organic matter content and the preservation of delicate sedimentary banding in the slaty mudstones. Based on the recorded foraminifera and ammonites the Bahía Thetis Formation was assigned to the Late Campanian-?Early Maastrichtian (Olivero et al. 2003, Caramés and Malumián 2006).
Policarpo Formation
The Policarpo Formation is very well exposed along the Atlantic shore of Península Mitre, between Policarpo Cove and San Vicente Cape (Furque and Camacho 1949, Olivero et al. 2002, 2003). The extensive, but less well-exposed rocks forming the NW belt of strata in the Sierra de Noguera and Sierra de Apen are also included in the Policarpo Formation (Fig. 1). In the type area of Península Mitre the Policarpo Formation includes a minimum thickness of 350 m, and probably more than 700 m, of bioturbated, tuffaceous, monotonous organic-rich, dark gray, very hard sandy mudstones and silty sandstones. Fresh-pyroclastic material is relatively abundant, and some fine-grained tuffaceous sandstones are almost exclusively composed of zoned, euhedral plagioclase crystals and volcanic glass. Most part of the Policarpo Formation consists of a crude alternation of tuffaceous mudstones and silty sandstones but the stratification is not always apparent due to the intense bioturbation. Totally bioturbated thick packages occur repeatedly, and they are commonly characterized by a high abundance of sandy, agglutinated tubes of Schaubcylindrichnus (previously assigned to "Terebellina"). In addition to a dense background mottling, the trace fossils Tasselia, Rhizocorallium, Phycodes, Teichichnus and small Chondrites are occasionally recorded (Fig. 4d). Some horizons bear a dense concentration of large Zoophycos burrows. Relatively abundant invertebrate body fossils are common in certain concretionary horizons, but they are very difficult to recover due to the extreme hardness of the rocks. Irregular echinoids are very common near the faulted contact with the Bahía Thetis Formation near Cabo San Vicente. Solitary corals, scaphopods, gastropods, bivalves, and ammonites are less abundant. One horizon at the contact with the Río Bueno Formation near Puesto Río Bueno, bears relatively wellpreserved specimens of Diplomoceras sp., Anagaudryceras sp., Maorites densicostatus (Kilian and Reboul), Maorites sp. and large Pachydiscus (Neodesmoceras) sp. (Fig. 5c-f). Near Puesto Donata, very well-preserved specimens of the gastropod Stru-thioptera cf. gregaria Wilckens were recovered. The foraminifera are dominated by agglutinated cosmopolitan forms that include the oldest record in the Austral Basin of Spiroplectammina spectabilis (Olivero et al. 2002, 2003, Olivero and Malumián 2008). Dinocysts are scarce and not wellpreserved, but two assemblages of Maastrichtian and Danian age were tentatively recognized (Olivero et al. 2003).
CONCLUDING REMARKS
The first fossils to be discovered in the clay-slate formation were Cretaceous crinoids, gastropods, bivalves, and ammonites found by Darwin in 1834 on Mount Tarn (Darwin 1846). This discovery was followed soon by new records of Cretaceous invertebrates, notably among these were the findings of additional fossil mollusks in Mount Tarn by Hombron and Grange in 1837; belemnites in Nassau Bay by Dana in 1839; and bivalves and ammonites in 1887-1890 in the islands of Saint Peter and Saint Paul, Magellan Channel, reported by White (see Andersson 1906, Bonarelli 1917, Kranck 1932). Despite these discoveries of Cretaceous fossils in the clay-slate formation in distant localities, stretching more than 500 km between Navarino Island and Brunswick Peninsula south of Punta Arenas on the Strait of Magellan, a strong feeling of a Paleozoic age for most of the clay-slate formation still grew. This was based mainly on comparisons of the regional features of the lowgrade metamorphic rocks with similar features in Paleozoic orogenic belts, particularly the basement in the British Isles and in Alpine areas, which at that time was thought to be of Paleozoic age (see Kranck 1932), and on the age interpretations of the dubious fossils found by Lovisato in 1881 in Staten Island (see Harrington 1943). However, the presumed Cambrian quartzites and schists from Staten Island turned out to be Jurassic acidic tuffs and volcaniclastic, foliated rocks, and the dubious Paleozoic fossils were probably part of the fibrous structure of Cretaceous inoceramid shells (Richter 1925) or belemnite rostra (Harrington 1943). As has been confirmed by additional fossil discoveries in the clay-slate formation of the main island of Tierra del Fuego, including the first locality visited by Darwin on the island (Good Success Bay), we now know that all these rocks are of Cretaceous age (Olivero and Martinioni 1996a, Olivero and Medina 2001). The relatively homogeneous lithology is a major obstacle to stratigraphic subdivision; nonetheless, particular combinations of fossil content, ichnofabric, and lithological features make it possible to recognize characteristic sedimentary successions in the Lower and Upper Cretaceous rocks. In the Lower Cretaceous, only Aptian-Albian fossils have been recovered; however, the Yahgan Formation has thick horizons with massive sandstones and sandy turbidites, a lithological association that is not found in the homogenous, black slaty mudstones and tuffs of the Beauvoir Formation. The Yahgan Formation also has localized horizons dominated by the trace fossils Chondrites (Fig. 4b) and Zoophycos, an ichnoassemblage that is consistent with the inferred setting of a low-oxygenated slope (cf. Uchman 2007) in a volcaniclastic apron environment (Olivero and Martinioni 1996b, Olivero and Malumián 2008). The black, fine-grained rocks of the Beauvoir Formation contain localized horizons with Nereites missouriensis (Fig. 4a). This ichnofossil is a typical component of the Nereites ichnosubfacies, which is distributed in distal flysch facies and characterized by the dominance of deposit-feeding invertebrates, such as the tracemaker of Nereites (cf. Seilacher 1974, Uchman 2007). Accordingly, these fine-grained deposits probably reflect distal basin plain settings, occupying an intermediate position between the Pacific volcaniclastic apron of the Yahgan Formation and the typical, South-American cratonic slope settings of the Lower Cretaceous rocks in the Austral Basin (Wilson 1991). In the Upper Cretaceous, four distinctive successions are recognized: 1) the lower part (Turonian-Coniacian) of the "Estratos de Buen Suceso", 2) the upper part (Santonian-Lower Campanian) of the "Estratos de Buen Suceso"; 3) the Upper Campanian-?Lower Maastrichtian Bahía Thetis Formation; and 4) the Maastrichtian-Danian Policarpo Formation (Olivero and Medina 2001, Olivero et al. 2002, 2003, Olivero and Malumián 2008). The lower part of the "Estratos de Buen Suceso" is characterized by marlstones, impure limestones, and fine-grained silty sandstones with T. madagascariensis, whereas the upper part is characterized by dark slaty mudstones, tuffs, and silty sandstones. The latter beds appear to record the earliest successions with extensively bioturbated horizons in the basin, probably reflecting for the first time prolonged periods with well-oxygenated bottom conditions (Fig. 4c). These beds alternate with those showing evidence for more oxygen-deficient bottom conditions, lacking trace fossils and populated locally by Inoceramus (Platyceramus) sp. and Spheno-ceramus sp. (Olivero and Medina 2001). The resedimented conglomerates, pebbly mudstones, and sandy turbidites of the Bahía Thetis Formation record a very distinctive petrographic composition, with clear evidence that the source of clastic material was uplifted Andean rocks, including foliated, Jurassic acidic volcanics and radiolarian-rich Cretaceous slates. These coarse-grained clastic rocks probably reflect deposition in submarine fans, with a source area in the hinterland part of the growing Fuegian Andes. The associated dark, organic-rich slaty mudstones (Fig. 2e) are thought to have originated in channel margin or more distant basin settings. The foraminiferal content and the generalized absence of trace fossils are consistent with the interpreted dysoxic bottom conditions (Olivero et al. 2003, Caramés and Malumián 2006). The Bahía Thetis Formation probably records the last occurrence of inoceramid bivalves in the Austral Basin (Olivero et al. 2003, 2004). The Maastrichtian-Danian Policarpo Formation records a marked, regional change of paleoceanographic conditions in the basin as is evidenced by the thoroughly bioturbated ichnofabric that characterizes thick sedimentary packages. This change in the ichnofabric occurs in the Maastrichtian Policarpo Formation just above the inoceramid extinction level in the Fuegian Andes. The inoceramids were apparently adapted to warm and poorly oxygenated waters and their global extinction pulses, during the mid-Maastrichtian, were related to cooling and enhanced bottom ventilation, promoted by circulation of deep Antarctic waters (MacLeod et al. 1996). The strong contrast in the intensity of bioturbation between the Upper Campanian-Lower Maastrichtian Bahía Thetis Formation, almost devoid of trace fossils, and the highly bioturbated Maastrichtian-Danian Policarpo Formation reflects a change from dysoxic-anoxic to well ventilated bottom conditions, probably associated with a cooling trend of bottom waters in the austral deep oceans (Olivero et al. 2003, 2004). In addition, the Policarpo Formation also records a distinctive petrographic composition, which is dominated by fresh volcaniclastic material implying a coeval volcanic pulse in the magmatic arc.
ACKNOWLEDGMENTS
We thank N. Malumián (SEGEMARCONICET) for fruitful discussion over the years on the geology of Tierra del Fuego. D. Martinioni (CADIC-CONICET) helped with part of the fieldwork and analysis of information in German papers. We thank P. Torres Carbonell (CADIC-CONICET) for a critical review of an early manuscript. M.B. Aguirre- Urreta (UBA-CONICET) provided the pictures of the figured original material collected by Darwin in Brunswick Peninsula and housed in the Natural History Museum, London. We thank the reviewers A. Crame, J.S. Crampton, I.W.D. Dalziel, and M.D. Suárez for constructive observations that improved the original manuscript. This study was financed by PIP 5100 CONICET and PICTO 36315 FONCYT.
WORKS CITED IN THE TEXT
1. Aguirre Urreta, M.B. and Suárez, M. 1985. Belemnites de una secuencia turbidítica volcanoclástica de la Formación Yahgan-Titoniano-Cretácico inferior del extremo sur de Chile. 4° Congreso Geológico Chileno (Antofagasta), Actas 1: 1-16.
2. Andersson, J.G. 1906. Geological fragments from Tierra del Fuego. Bulletin of the Geological Institution, University of Upsala 8: 169-183.
3. Bonarelli, G. 1917. Tierra del Fuego y sus turberas. Anales del Ministerio de Agricultura de la Nación, Sección Geología, Mineralogía y Minería 12(3): 1-119, Buenos Aires.
4. Camacho, H.H. 1967. Las transgresiones del Cretácico superior y Terciario de la Argentina. Revista de la Asociación Geológica Argentina 22: 253-280.
5. Caramés, A. and Malumián, N. 2006. La Familia Rzehakinidae (Foraminifera) en el Cretácico superior-Paleógeno de la cuenca Austral y la plataforma continental atlántica adyacente, Argentina. Ameghiniana 43: 649-668.
6. Crampton, J.S. 1996. Inoceramid bivalves from the Late Cretaceous of New Zealand. Institute of Geological and Nuclear Sciences Monograph 14: 1-188.
7. Crampton, J.S. 2004. Shell composition, cryptic costae, complex composite molds, and taphonomic chicanery in Mytiloides (Inoceramidae, Bivalvia). Journal of Paleontology 78: 1091-1096.
8. Dalziel, I.W.D., De Wit, M.J. and Palmer, K.F. 1974. Fossil marginal basin in the southern Andes. Nature 250: 291-294.
9. Darwin, C. 1846. Geological observations on South America. Being the third part of The Geology of the Voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith, Elder and Co., 279 p., London.
10. Doello Jurado, M. 1922. Note préliminaire sur les résultats géologiques de l'Expédition de l'Université de Buenos-Ayres a la Terre du Feu (1921). Compte Rendu du 13eme Congrès Géologique International (Liege) 3: 1519-1520.
11. Dott, R.H., Winn, R.D., De Wit, M.J. and Bruhn, R.L. 1977. Tectonic and sedimentary significance of Cretaceous Tekenika beds of Tierra del Fuego. Nature 266: 620-622.
12. Flores, M.A., Malumián, N., Masiuk, V. and Riggi, J.C. 1973. Estratigrafía cretácica del subsuelo de Tierra del Fuego. Revista de la Asociación Geológica Argentina 28: 407-437.
13. Furque, G. and Camacho, H.H. 1949. El Cretácico superior de la costa Atlántica de Tierra del Fuego. Revista de la Asociación Geológica Argentina 4: 263-297.
14. Harrington, H.J. 1943. Observaciones geológicas en la isla de los Estados. Anales del Museo Argentino de Ciencias Naturales 41 (Geología), Publicación 29: 29-52.
15. Katz, H.R. 1972. Plate tectonics and orogenic belts in the South-East Pacific. Nature 237: 331-332.
16. Katz, H.R. and Watters, W.A. 1966. Geological investigations of the Yahgan Formation (Upper Mesozoic) and associated igneous rocks of Navarino Island, southern Chile. New Zealand Journal of Geology and Geophysics 9: 323-359.
17. Kohn, M.J., Spear, F.S., Harrison, T.M. and Dalziel, I.W.D. 1995. AR40/AR39 geochronology and P-T-t paths from Cordillera Darwin metamorphic complex, Tierra del Fuego, Chile. Journal of Metamorphic Geology 13: 251-270.
18. Kranck, E.H. 1932. Geological Investigations in the Cordillera of Tierra del Fuego. Acta Geographica 4: 1-231.
19. Lahsen, A. and Charrier, R. 1972. Late Cretaceous ammonites from Seno Skyring-Strait of Magellan area, Magallanes Province, Chile. Journal of Paleontology 46: 520-532.
20. Macdonald, D.I.M. 1986. Proximal to distal sedimentological variation in a linear turbidite trough: implications for the fan model. Sedimentology 33: 243-259.
21 Macellari, C.E. 1979. La presencia del género Aucellina (Bivalvia, Cretácico) en la Formación Hito XIX (Tierra del Fuego, Argentina). Ameghiniana 16: 143-172.
22. MacLeod, K.G., Huber, B.T., and Ward, P.D. 1996. The biostratigraphy and paleobiogeography of Maastrichtian inoceramids. In Ryder, G., Fastovsky, D. and Gartner, S. (eds.) The Cretaceous-Tertiary Event and Other Catastrophes in Earth History, Geological Society of America, Special Paper 307: 361-373.
23. Medina, F.A. 2007. Inocerámidos Albianos de la Isla James Ross, Antártica. 6º Simposio Argentino y 3º Latinoamericano sobre Investigaciones Antárticas, CD-ROM Resumen Expandido GEORE802, 4 p., Buenos Aires.
24. Olivero, E.B. and Malumián, N. 2008. Mesozoic-Cenozoic stratigraphy of the Fuegian Andes, Argentina. Geologica Acta 6: 5-18.
25. Olivero, E.B. and Martinioni, D.R. 1996a. Late Albian Inoceramid bivalves from the Andes of Tierra del Fuego. Age implications for the closure of the Cretaceous marginal basin. Journal of Paleontology 70: 272-274.
26. Olivero, E.B. and Martinioni, D.R. 1996b. Sedimentología de la Formación Yahgán (Jurásico-Cretácico) en Tierra del Fuego. 13º Congreso Geológico Argentino y 3° Congreso de Exploración de Hidrocarburos (Buenos Aires), Actas 2: 45-59.
27. Olivero, E.B. and Medina, F.A. 2001. Geología y paleontología del Cretácico marino en el sureste de los Andes Fueguinos, Argentina. Revista de la Asociación Geológica Argentina 56: 344-352.
28. Olivero, E.B., Malumián, N., Palamarczuk, S. and Scasso, R. 2002. El Cretácico superior-Paleógeno del área del Río Bueno, costa atlántica de la Isla Grande de Tierra del Fuego. Revista de la Asociación Geológica Argentina 57: 199-218.
29. Olivero, E.B., Malumián, N. and Palamarczuk, S. 2003. Estratigrafía del Cretácico superior-Paleoceno del área de bahía Thetis, Andes Fueguinos, Argentina: acontecimientos tectónicos y paleobiológicos. Revista Geológica de Chile 30: 245-263.
30. Olivero, E.B., Malumián, N. and López C., M.I. 2004. Changes in bioturbation intensity near the inoceramid extinction horizon: new data from Antarctica and Tierra del Fuego. First International Congress on Ichnology, Ichnia 2004 (Trelew), Abstract Book: 63-64.
31. Richter, M. 1925. Beiträge zur Kenntniss der Kreide in Feuerland. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 52B: 524-568.
32. Seilacher, A. 1974. Flysch trace fossils: Evolution of behavioural diversity in the deep-sea. Neues Jahrbuch für Geologie und Paläontologie Monatshefte, Jahrgang 1974: 233-245.
33. Spath, L.F. 1953. The Upper Cretaceous cephalopod fauna of Graham Land. Falkland Island Dependencies Survey, Scientific Report 3: 1-60.
34. Suárez, M.D., Hervé, M.A. and Puig, G.A. 1985. Carta geológica de Chile. Hoja Isla Hoste e islas adyacentes. 12 Región. Servicio Nacional de Geología y Minería 65: 1-113.
35. Torres Carbonell, P.J., Olivero, E.B. and Dimieri, L.V. 2008. Control en la magnitud de desplazamiento de rumbo del Sistema Transformante Fagnano, Tierra Del Fuego, Argentina. Revista Geológica de Chile 35: 65-79.
36. Uchman, A. 2007. Deep-sea ichnology: development of major concepts. In Miller, W. (ed.) Trace Fossils Concepts, Problem, Prospects, Elsevier B.V.: 248-263, Amsterdam.
37. Walaszczyk, I., Marcinowski, R., Praszkier, T., Dembicz, K., and Bienkowska, M. 2004. Biogeographical and stratigraphical significance of the latest Turonian and Early Coniacian inoceramid/ammonite succession of the Manasoa section on the Onilahy River, south-west Madagascar. Cretaceous Research 25: 543-576.
38. Wilson, T.J. 1991. Transition from back-arc to foreland basin development in the southernmost Andes: Stratigraphic record from the Ultima Esperanza District, Chile. Geological Society of America, Bulletin 103: 98-111.
39. Winn, R.D. 1978. Upper Mesozoic flysch of Tierra del Fuego and South Georgia Island. A sedimentologic approach to lithosphere restoration. Geological Society of America, Bulletin 89: 553-547.
Darwin's great patagonian tertiary formation at the mouth of the río Santa Cruz: a reappraisal
Ana Parras and Miguel Griffin
Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa y CONICET, Santa Rosa, La Pampa. Emails: aparras@exactas.unlpam.edu.ar, miguelgriffin@aol.com
ABSTRACT
The classical section described by Darwin at the mouth of the Río Santa Cruz was a landmark in the later stratigraphy of the marine Cenozoic of Patagonia. Sedimentological and paleontological studies of rocks from the Monte León Formation at the Darwin and Mount Entrance sections - mouth of the Río Santa Cruz - suggest that they were deposited in an early Miocene inner shelf to subtidal - or even intertidal at the top - environment. The base of the sections is covered and the top is unconformably overlain by Quaternary deposits. The invertebrate fauna (mainly mollusks) mentioned herein - the earliest of which were collected by Darwin and described by Sowerby - were collected from the two sections with a detailed stratigraphic control. It includes brachiopods, echinoderms, scaphopods, bivalves, and gastropods. The similarity of this fauna with that one from the sections at Restinga Norte, Cabeza de León, Las Cuevas, Yegua Quemada and Monte Observación, all south of the study area, allow correlation with them. In addition to the equivalence of the faunas, sedimentological features at all these localities are also the same. Therefore, all the shell beds involved should be referred to the Punta Entrada Member of the Monte León Formation. The Monte Observación Member should be restricted to its original use and included as a member at the base of the overlying Santa Cruz Formation, outcrops at Cerro Monte León and Cerro Observación.
Keywords: Darwin Section; Mount Entrance Section; Mouth of Santa Cruz River; Paleoenvironments; Invertebrates; Patagonia.
RESUMEN: La Gran Formación Terciaria Patagónica de Darwin en la desembocadura del río Santa Cruz: una revaluación. La sección clásica descripta por Darwin en la desembocadura del río Santa Cruz fue un hito en el posterior desarrollo de la estratigrafía del Cenozoico marino de Patagonia. Los estudios sedimentológicos y paleontológicos realizados en sedimentitas de la Formación Monte León, en las secciones Darwin y Monte Entrada, en el área de la desembocadura del río Santa Cruz, sugieren que las mismas fueron depositadas en un ambiente de plataforma interna a submareal en la base, hasta intermareal en el techo, durante el Mioceno temprano. La base de las secciones se encuentra cubierta y su techo es cortado en forma discordante por depósitos del Cuaternario. La fauna de invertebrados (mayormente moluscos) mencionada en este trabajo, cuyos primeros representantes fueran coleccionados por Darwin y descriptos por Sowerby, fue colectada con un detallado control estratigráfico a lo largo de las dos secciones. La misma incluye braquiópodos, equinodermos, escafópodos, bivalvos y gasterópodos. La similitud en la composición taxonómica, como así también en las características sedimentológicas, entre estas secciones y aquellas ubicadas más al sur como son Restinga Norte, Cabeza de León, Las Cuevas, Yegua Quemada y Monte Observación, sugiere que las mismas se correlacionan lateralmente. Por lo tanto estos niveles deben ser referidos al Miembro Punta Entrada de la formación Monte León, restringiéndose el Miembro Monte Observación a los niveles basales de la suprayacente Formación Santa Cruz, aflorantes en Cerro Monte León y Cerro Observación.
Palabras clave: Sección Darwin; Sección Monte Entrada; Desembocadura del río Santa Cruz; Paleoambientes; Invertebrados; Patagonia.
INTRODUCTION
On April 13th 1834, HMS Beagle anchored within the mouth of the Río Santa Cruz, a few miles upstream from the Atlantic Ocean. During his numerous excursions ashore, Darwin made geological observations on the cliffs that rise along the southern bank of the river. He also assembled a collection of invertebrate marine fossils later described by Sowerby (1846). Thus, the locality from where this material was collected became the type locality of most of the taxa described by Sowerby. The main purposes of this work are to pinpoint and re-describe as accurately as possible the locality at the mouth of the Río Santa Cruz where Charles Darwin (1846, p. 112-113) described the geological section of marine Tertiary rocks, and discuss the possible paleoenvironmental conditions in which it was deposited. This should aid in the formulation of more precise correlations of the beds mentioned by Darwin with others exposed in the area, and thus clarify the meaning of the formal names proposed by later authors for the rocks involved. Darwin (1839, 1846) himself included these rocks in his "great Patagonian tertiary formation" or "Patagonian Tertiary Formation", a unit subject to very different interpretations ever since. According to his description of the beds exposed and after a survey of the area to try to identify the exact location of his section, we were able to identify it on the southern bank of the Río Santa Cruz, a short distance away from its mouth in the Atlantic Ocean. The rocks exposed in this area are now included in the lithostratigraphic unit known as Monte León Formation (Bertels 1970) and deemed to be of late Oligocene-early Miocene age (Malumián 1999, 2002, Barreda and Palalmarczuk 2000). This unit is exposed along the coast of Patagonia between Puerto San Julián and Coy Inlet, at the mouth of the Río Coyle (or Coig) (Fig. 1). It comprises mainly sandstones and siltstones with a high content of pyroclastic material, and carries numerous shell beds.
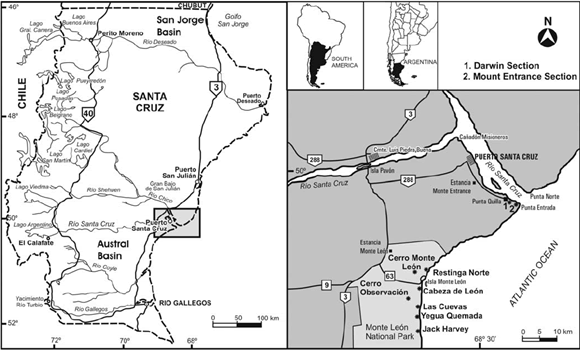 Figure 1: Location map of the Darwin Section and the Mount Entrance Section at the mouth of the Río Santa Cruz.
Figure 1: Location map of the Darwin Section and the Mount Entrance Section at the mouth of the Río Santa Cruz.
In order to clarify the exact location of Darwin's section, we surveyed the coast along the southern bank of the river. Based on his writings (1846, p. 112-113) it became clear the section is the one lying just east of the present port of Punta Quilla (S 50° 07' 39.9"; W 68° 24' 10.04"), a point where the marine Tertiary rocks are best exposed (Fig. 2), and where they best meet the description and measurements he offers in his "Geological Observations…".
 Figure 2: Monte León Formation at the mouth of the Río Santa Cruz, Darwin Section, showing Darwin's strata and facies associations described herein.
Figure 2: Monte León Formation at the mouth of the Río Santa Cruz, Darwin Section, showing Darwin's strata and facies associations described herein.
As there has been some confusion as to the exact location of Darwin's locality, we also measured and described the section lying further east, approximately one kilometer away from Mount Entrance, at the very mouth of the river (S 50° 08' 32.2"; W 68° 22' 23.6"). The two localities lie within the present Department of Corpen Aike, in the province of Santa Cruz (Fig. 1). They both carry a rich fauna of exceptionally well preserved mollusks that can be counted among the best in southern South America. They have been mentioned in the literature (Becker 1964, Bertels 1970, 1980, Di Paola and Marchese 1973, Olivera 1990, del Río and Camacho 1998, Barreda and Palamarczuk 2000), but detailed descriptions that could enable correlation and paleoenvironmental reconstructions are still lacking. Study of these two stratigraphic sections allowed recognition of sedimentary facies, and hence the dominant deposition mechanisms could be interpreted. These facies were grouped into three facies associations and a depositional model thus emerged. Detailed analysis of the taxonomic composition of the fossil assemblages revealed the presence of at least 38 species of bivalves and 70 species of gastropods, in addition to scaphopods, echinoderms, and brachiopods. Diffe-rences among the different shell beds suggest a complex history. The different fossil concentrations were originated in different settings, ranging from inner shelf to nearshore subtidal environments - and even intertidal, representing deposition during the early Miocene transgression in Patagonia.
GEOLOGICAL FRAMEWORK
The surface of Patagonia is covered mainly by marine rocks deposited during the Atlantic transgressions that occurred during the late Cretaceous, Paleogene and Neogene. These are intercalated with non marine rocks and non deposition and erosion intervals (Malumián 1999). One of these transgressions is commonly referred to as "Patagoniano" and occurred during the high sea level period that began in the latest Oligocene and ended in the early middle Miocene. The final phases of this transgression are coincident with the Neogene climatic optimum. It covered the largest extent of land among the various Cenozoic transgressions that occurred in Patagonia (Malumián 2002). This transgression deposited rocks containing the Paleogene/Neogene boundary in Patagonia and in the Cordillera sector of southern Chile. They are presently included in the San Julián and Monte León formations along the coast of Santa Cruz and in the Centinela Formation in the Andean region of this province. Exposures of these units lie within the Austral Basin. Further north, the rocks deposited during the Patagonian transgression are included in the Chenque and Gaiman formations, in the San Jorge Basin. In southern Chile, these rocks are known as Guadal Formation, and are exposed south of Lago General Carrera. The basement rocks underlying the Cenozoic deposits are a Jurassic lava-pyroclastic complex known as Bahía Laura Group, including the Chon Aike Formation (ignimbrites and scarce rhyolitic lavas and tuffs) and the La Matilde Formation (tuffs, claystones, siltstones and coal beds). These two units are interdigitated and unconformably overlain, in the San Julián area, by the San Julián Formation. This late Oligocene unit includes predominantly marine clastic -sandstone to siltstone - rocks (Parras and Casadío 2005, Parras et al. 2008). Deposition of the probably unconformably overlying Monte León Formation occurred during the late Oligocene and early Miocene. This unit includes siltstones, sandstones and tuffs (Bertels 1970, 1980), and is overlain by the early to middle Miocene continental rocks of the Santa Cruz Formation. Unconformably overlying these are variably thick Quaternary deposits of the Shingle Formation described by Darwin too (Darwin 1839, 1846). The earliest record of marine rocks in Patagonia was by d'Orbigny (1842 a, b), who explored this part of South America during the years 1826-1833. D'Orbigny himself never went ashore further south than the coast of northern Patagonia. However, he named all the deposits that constitute the cliffs along the coast between Carmen de Patagones (Río Negro) and the Strait of Magellan as "Tertiaire Patagonien". Although he did not inspect the cliffs further south, he did have available the large oysters from San Julián, which he believed were the same ones as he had collected in Río Negro and further north in Entre Ríos. Darwin was aware of this, and therefore used an equivalent name for these rocks. However, he also noticed that there were differences between the faunas from the southern and northern areas in which his "Patagonian Tertiary Formation" was exposed. After these initial surveys the area remained largely unexplored from a geological point of view. A Chilean settlement near the present town of Puerto Santa Cruz was headed by Vidal Gormaz, who collected a few fossils from the cliffs surrounding the settlement and sent them to R.A. Philippi, then at the Museo Nacional de Historia Natural in Santiago de Chile. These fossils were described by Philippi (1887), but hardly any comments were made on the geology of the area, except to note that the specimens came from the Tertiary marine beds there exposed. It was not until the final years of the 19th Century that this region was again surveyed and its geology described in more detail. This was due mainly to three early explorers who visited the mouth of the Río Santa Cruz and assembled the data that were then - and for many years - used to formulate the first stratigraphic divisions and the earliest paleogeographic and paleobiogeographic interpretations about the rocks in the area. In the first place should be mentioned Carlos Ameghino, brother of Florentino Ameghino-the famed Argentine paleontologist. C. Ameghino surveyed the area and collected fossils that were subsequently studied by his brother. Although he did not publish his results extensively, his keen and detailed geological observations were later used by Florentino to formulate the earliest biostratigraphic scheme of the Tertiaire Patagonien or the Great Patagonian Formation. Thus, based on the fossil content (mainly mollusks) of these marine beds at the mouth of the Río Santa Cruz and in the surroundings of Puerto San Julián, F. Ameghino (1898) subdivided the marine beds he had named (Ameghino 1896) "Formación Patagónica" into two chronostratigraphic units he called "Juliense" and "Leonense". Overlying these (Ameghino 1900-1902) he recognized the continental "Formación Santacruceña", at the base of which he could distinguish a marine stage he called "Superpatagónico". The faunas of marine invertebrates that were the basis of Ameghino's subdivisions were collected by Carlos and described by Maurice Cossmann (1899) and Hermann von Ihering (1897, 1899, 1902, 1907, 1914). During the same years, the area was also visited by J. B. Hatcher, geologist in charge of the Princeton Expedition to Southern Patagonia. Hatcher's observations were published in the Reports and Geology of this expedition (Hatcher 1903) and in a series of papers in the American Journal of Science (Hatcher 1897, 1900). The large collection of Tertiary marine fossils that he assembled was described by Ortmann (1902). Hatcher's conclusions on the geology of the area were different from those of Ameghino. Hatcher did not recognize the existence of the "Superpatagónico", as the fauna described by Ortmann was essentially the same one as the one in the underlying "Formación Patagónica". This initial controversy pervaded all further work on these units and has never been satisfactorily resolved. The reasons for this are varied and include imperfect knowledge on the sedimentology and stratigraphy at the localities involved, inaccurate location of some of these localities, outdated taxonomic placement of some of the fossil material on which the biostratigraphy was based, among other. At about the same time, C. Bicego was entrusted by H. von Ihering with the task of assembling further collections at the localities initially visited by C. Ameghino. His large collection of mollusks was described by von Ihering, but Bicego himself did not provide further geological data beyond that mentioned by Ameghino. Wichmann (1922), Windhausen (1931) and Feruglio (1949) published detailed accounts on the geological features of Patagonia, including the marine Cenozoic rocks. Their detailed sections and geological observations are generally easy to corroborate in the field. However, interpretations largely reflect the ideas of Ameghino and von Ihering. After these initial studies, the use of "Patagonian Formation" for the Cenozoic marine rocks in southern South America was widespread, although the exact chronostratigraphic and lithostratigraphic meaning of this formational name remained somewhat unstable. Accounts on the uses and controversies attached to such usage can be found in Camacho (1974, 1979), Zinsmeister (1981), Legarreta and Uliana (1994), and Malumián (1999, 2002). At the mouth of the Río Santa Cruz, these rocks were formally named Monte León Formation by Bertels (1970). The unit includes mainly yellowish-grey siltstones and fine sandstones with a high pyroclastic content and intercalated shell beds. These rocks record a relative sea level increase characterized by the greatest extent of the Patagonian Cenozoic sea, thus allowing input of corrosive Antarctic water onto the Argentine continental shelf (Malumián 2002). Most authors agree that in the study area, i.e. the mouth of the Río Santa Cruz, these rocks were deposited in a shallow marine environment, under littoral and neritic conditions (Panza et al. 1995, Barreda and Palamarczuk 2000). Bertels (1980) suggested outer shelf conditions for the lower part of this formation. Panza et al. (1995) stated that towards the top of the unit conditions became progressively shallower, ending in a marshy environment just before the onset of the continental environment in which the overlying Santa Cruz Formation was deposited. The abundant tuff beds and cineritic material in these rocks led Bertels (1970) and Di Paola and Marchese (1973) to suggest they originated in coetaneous volcanic events in the Cordillera. The ash was probably transported towards the coast by the western winds and deposited in an environment that was protected from the action of marine currents. Although discrepancies still exist concerning the age of the Monte León Formation, most authors refer it to the late Oligocene (Bertels 1970) or late Oligocene - early Miocene (Malumián 1999, 2002). Yet, Náñez (1988) restricted it to the Oligocene (middle section of P19 Zone and top of P21 Zone) while suggesting that the top may reach into the early Miocene. This was later confirmed by 40Ar/39Ar ages published by Fleagle et al. (1995). Their sample came from the top of the unit at Cerro Monte León and it yielded an age of 19.33 Ma (Burdigalian, early Miocene). More recently Barreda and Palamarczuk (2000) considered that in the study area the Monte León Formation should be restricted to the early Miocene on the basis of its palynomorph content, an age we agree with herein.
METHODOLOGY
Just east of Punta Quilla (Darwin section; S50°07'39.9"; W68°24'10.04") and at Mount Entrance (S50°08'32.2"; W68° 22'23.6") we measured detailed (1:10) stratigraphic-sedimentologic sections (Fig. 3). We recorded bed geometry, lithology, grain size, sedimentary structures, and color. Each bed was sampled and fossils and trace fossils were photographed and collected. Taphonomic observations were recorded too. Close-packing and size-sorting were estimated following the semi-quantitative categories for coarse bioclastic fabrics of Kidwell and Holland (1991). Terminology for geometry of concentrations and orientation of bioclasts follows Kidwell et al. (1986). Sedimentary facies were distinguished and grouped in facies associations for paleoenvironmental analysis. The invertebrate fauna collected was identified taxonomically. The different beds mentioned by Darwin (1846, p. 112-113) were identified as was the exact provenance of the type material of the species described by Sowerby (1846).

 Figure 3: Studied stratigraphic sections showing identified facies associations.
Figure 3: Studied stratigraphic sections showing identified facies associations.
SEDIMENTOLOGY
Sections
The type section of the Monte León Formation lies on the right bank of the Río Santa Cruz, one kilometer upstream of its mouth into the Atlantic Ocean (Bertels 1970, p. 499). However, Bertels (1970) did not describe or illustrate her section until a decade later (Bertels 1980, p. 214-217), when she also defined the Punta Entrada Member of the Monte León Formation. This locality is precisely where our Mount Entrance section lies. According to her description (Bertels 1980) the section measures there 103.8 m, but this could not be confirmed in the field by us, as the bottom of the section is completely covered by modern foothill deposits and only 60 m are exposed. Therefore, the base of Bertels' section - as inferred from her description - must have been measured at Darwin's section, where the exposure is complete, although the top part is here extremely difficult to reach. Riggi (1978, p. 166) measured 89 m but it remains unclear exactly where his section is (his Mount Entrance section lies "on the right bank of the Río Santa Cruz and its mouth in the ocean"). Barreda and Palamarczuk (2000) measured 80 m south of the mouth of the river, but most other published sections are compounds based on partial sections measured at various points ranging between south of the mouth of the river to 30 km upstream along the right bank (Olivera 1990, del Río and Camacho 1998). Other authors based their stratigraphic framework on unpublished data (Becker 1964) or previous work by others (Di Paola and Marchese 1973). According to Darwin's description, the section mentioned by him is the one just east of Punta Quilla (our Darwin Section, Figs. 2 and 3). This is where the cliff is best exposed and where the description and measurements provided by Darwin himself best agree with field observations. In order to refer the beds described by Darwin to the type section, we provide herein a description of both, even if lithology is very similar - as is fossil content. The Darwin Section is more complete because the lower beds lie exposed at low tide. However, access to the topmost beds is extremely difficult. This is probably the reason why the section at Mount Entrance (and nearby) has been better studied.
a) Darwin Section (S50º07'39.9"; W68º 24'10.04") 0-1.2 m: 1.2 m Grey tuffaceous clayish siltstones with abundant fossils and numerous Ophiomorpha isp. Dominant species is "Turritella" patagonica. It appears in clump-geometry packets. Also recorded are Nucula sp., Limopsis insolita, Cucullaea alta, Crassostrea? hatcheri, Fasciculicardia patagonica, Pleuromeris cruzensis, Dosinia laeviuscula, Ameghinomya darwini. Close-packing within these clumps is dense/loose (25%), size-sorting is poor, orientation chaotic, disarticulation is total, fragmentation low in some clumps and high in others, abrasion is nil. Encrusting and bioerosion is observed on the valves of C.? hatcheri. The taxa recorded were en-crusting bryozoans, sponge borings (En-tobia isp.), polychaetes (Maeandropolydora isp.), and bivalve borings such as Gastro-chaenolites isp. 1.2-6 m: 4.8 m Grey tuffaceous clayish siltstones similar to the previous ones. This bed carries three lenses, each ca. 0.2 m thick and dominated by "Turritella" patagonica. These lenses are separated by siltstones with very few clumps that carry almost exclusively "T". patagonica. Also present are, among many other taxa, Nucula (Lamellinucula) reticularis, Iheringinucula crassirugata, Scaeoleda? ortmanni, Neilo ornata, Arca patagonica, Cucullaea alta, Limopsis insolita, Glycymeris cuevensis, Atrina magellanica, Neopanis quadrisulcata, Swiftopecten nodosoplicatus, Zygochlamys geminata, Zygochlamys quemadensis, Reticulochlamys proxima, Jorgechlamys centralis, Crassostrea? hatcheri, Pteromyrtea crucialis, Pleuromeris cruzensis, Fasciculicardia patagonica, Spissatella lyelli, Hedecardium? ameghinoi, Hedecardium? puelchum, Trachycardium pisum, Cardium patagonicum, Lahillia patagonica, Maorimactra indistincta, Macoma? santacruzensis, Retrotapes striatolamellata, Dosinia laeviuscula, Ameghinomya darwini, Ameghinomya meridionalis, Caryocorbula hatcheri, Panopea quemadensis, Liotia scotti, Gibbula (Phorcus) margaritoides, Valdesia dalli, Solariella dautzenbergi, Calliostoma santacruzense, Calliostoma peraratum, Calliostoma cossmanni, Calliostoma garretti, "Turritella" ambulacrum, Struthiochenopus santacruzensis, Perissodonta ornata, Perissodonta patagoniensis, Crepidula gregaria, Cerithioderma patagonica, Polinices santacruzensis, Polinices ortmanni, Glossaulax vidali, Lunatia consimilis, Sassia bicegoi, Cerithiopsis juliana, Cirsotrema rugulosum, Cirsotrema quemandense, Trophon santacruzensis, Xymenella dautzenbergi, Xymene cossmanni, Enthacanthus monoceros, Crassilabrum hatcheri, cf. Ocenebra iheringi, Urosalpinx archipatagonica, Peonza torquata, "Cominella" annae, Penion subrectus, Miomelon petersoni, Miomelon gracilior, Adelomelon pilsbryi, Neoimbricaria patagonica, Dentimargo deuterolivella, Austroimbricaria quemadensis, Zeadmete ameghinoi, Zeadmete cruzialis, Antimelatoma quemadensis, Splendrillia santacruzensis, Austrotoma cuevensis, Fusiguraleus iheringi, Terebra quemadensis, Terebra santacruzensis, Odostomia synarthrota, Turbonilla cuevensis, Turbonilla observatonis, Semiacteon argentinus, Cylichna juliana, Kaitoa patagonica, "Odontostomia" euryope, Dentalium sp. Within the lens close-packing is dense (55%), size-sorting poor - with juvenile and adult specimens alike - , orientation is chaotic, disarticulation is total, fragmentation very low, and abrasion nil. Encrusting and bioerosion occur on the oysters. 6-27 m: 21 m Yellowish-brown tuffaceous siltstones with at least two beds dominated by "Turritella" patagonica. They measure 1 and 2.5 m thick respectively. The top bed is the most conspicuous and shows sectors in which it is hardened into concretions that outstand in the section. Faunal composition almost identical to that of the underlying bed. Contacts are wavy, close-packing is dense/loose (25%) at some places and dense (55%) at others, size-sorting is poor (full size range of "T." patagonica), orientation is chaotic, disarticulation high with very few specimens keeping conjoined valves, fragmentation is high in some sectors and low in others, abrasion nil. Between 15 and 18 m above the base of this bed there are thin intercalated lenses of 0.05-0.3 m thickness dominated also by "T." patagonica. At the very top there is Dentalium in life position and Limopsis insolita. 27-30 m: 3 m Grey tuffaceous siltstones. Lithology similar to previous ones, but the color change from yellowish to grey is notable. 30-34 m: 4 m Grey tuffs. The first 0.4 m carry large Ophiomorpha isp. The rest of the bed is massive, with only a few sparse fragments of shells. 34-56 m: 22 m Yellow tuffaceous siltstone highly bioturbated, with numerous very large Thalassinoides isp. (up to 0.08 m diameter). Sparse body fossils include crabs, very small Lucinoidea in life position and larger fragments of maybe the same taxon. Preservation is gypsum replacement. The top 8 m carry very sparse valves of Cucullaea alta and Dosinia laeviuscula, in all cases disarticulate and convex-up, some of them broken. Numerous concretions (as recorded by Darwin) of 0.8 to 1.4 m diameter and conjoined in a "necklace" manner. An especially conspicuous bed of concretions is at 6 m from the bottom of this bed. 56-56.5 m: 0.2-0.5 m Very fine sandstone with abundant specimens of Dosinia laeviuscula. This is a monospecific bed with a wavy bioturbated contact at the bottom and a wavy one at the top. Close-packing is dense (55%), size-sorting is well sorted, orientation chaotic, disarticulation high, fragmentation is moderate to low, abrasion nil. At the base there are numerous tubes that reach down up to 0.7 m into the underlying bed. This shell bed is replaced laterally by light grey to white tuff of approximately the same thickness. 56.5-75.5 m: 19 m Interstratified siltstone, claystone and fine sandstone. At the bottom it carries right valves of Crassostrea? hatcheri, in all cases convex-down. At 11 m from the bottom the color changes from brown to yellowish-green.
b) Mount Entrance Section (S50º08'32.2"; W68º22'23.6") 0-17 m: 17 m Grey and greenish tuffaceous clayish siltstones with a few specimens of "Turritella" patagonica that may be isolated or forming clumps. At 7 m from the base there is a sandy lens dominated by this species but also containing a few brachiopods and bivalves. At 14 m there are large "necklace" concretions of about 0.3 m diameter. Between 16 and 17 m the bed is strongly bioturbated (Ophiomorpha isp. and other small tubes in all directions). 17-27 m: 10 m Very bioturbated tuffaceous sandstones. 27-27.8m: 0.8 m Fine tuffaceous sandstones, dark grey and very hard, with crossbedding. At the base it carries load structures. These sandstones are very bioturbated, with many tubes of about 0.01 m diameter placed obliquely to the stratification planes. 27.8-38.8 m: 11 m Tuffaceous bioturbated sandstones similar to the underlying bed. 38.8-39.4 m: 0.6 m Very fine sandstones with abundant fossils. This is a shell bed with a wavy contact at the base and a gradual transition at the top. Close-packing is dense (55%) at the base and loose (15%) at the top, size-sorting is well sorted, as complete specimens and fragments show approximately the same size, a few large whole bivalves are also observed, orientation is chaotic, disarticulation complete, fragmentation high, and many naticid and muricid boreholes are evident on the shells. This bed carries a very rich fauna of well preserved mollusks, including among others Nucula (Lamellinucula) reticularis, Iheringinucula crassirugata, Scaeoleda? ortmanni, Neilo ornata, Arca patagonica, Cucullaria darwini, Cucullaea alta, Limopsis insolita, Glycymeris cuevensis, Atrina magellanica, Neopanis quadrisulcata, Swiftopecten nodosoplicatus, Zygochlamys geminata, Zygochlamys quemadensis, Reticulochlamys proxima, Jorgechlamys centralis, Crassostrea? hatcheri, Pteromyrtea crucialis, Pleuromeris cruzensis, Fasciculicardia patagonica, Spissatella lyelli, Spissatella kokeni, Hedecardium? ameghinoi, Hedecardium? pauciradiatum, Hedecardium? puelchum, Trachycardium pisum, Cardium patagonicum, Lahillia patagonica, Maorimactra indistincta, Serratina jeguaenis, Macoma? santacruzensis, Retrotapes ortmanni, Retrotapes striatolamellata, Dosinia laeviuscula, Ameghinomya darwini, Ameghinomya meridionalis, Caryocorbula hatcheri, Panopea quemadensis, Liotia scotti, Homalpoma philippii, Gibbula (Phorcus) margaritoides, Valdesia dalli, Solariella dautzenbergi, Calliostoma santacruzense, Calliostoma peraratum, Calliostoma cossmanni, Calliostoma garretti, "Turritella" ambulacrum, "Turritella" patagonica, Struthiochenopus santacruzensis, Perissodonta ornata, Perissodonta ameghinoi, Perissodonta patagoniensis, Calyptraea eleata, Sigapatella americana, Crepidula gregaria, Cerithioderma patagonica, Vermetus incertus, Polinices santacruzensis, Polinices ortmanni, Glossaulax vidali, Lunatia consimilis, Sconsia ovulum, Sassia bicegoi, Sassia morgani, Cerithiopsis juliana, Cirsotrema rugulosum, Cirsotrema qemandense, Trophon santacruzensis, Xymenella dautzenbergi, Xymene cossmanni, Enthacanthus monoceros, Crassilabrum hatcheri, cf. Ocenebra iheringi, Urosalpinx archipatagonica, Peonza torquata, "Cominella" annae, Penion subrectus, Austrocominella cosmanni, Retizafra improvisa, Miomelon petersoni, Miomelon gracilior, Adelome lon pilsbryi, Neoimbricaria patagonica, Dentimargo deuterolivella, Austroimbricaria quemadensis, Zeadmete ameghinoi, Zeadmete cruzialis, Sveltia maior, Scalptia vidali, Antimelatoma quemadensis, Splendrillia santacruzensis, Austrotoma cuevensis, Fusiguraleus iheringi, Eoturris santacruzensis, Borsonia patagonica, Terebra quemadensis, Terebra santacruzensis, Odostomia synarthrota, Odostomia suturalis, Turbonilla cuevensis, Turbonilla observatonis, Turbonilla iheringi, Semiacteon argentinus, Cylichna juliana, Kaitoa patagonica, "Odontostomia" euryope, Dentalium sp. 39.4-59.4 m: 20 m Interstratified fine sandstone, siltstone and claystone. It carries a few interstratified concretions with bivalves and echinoderms. At 12 m from the base of this bed there is a color change from brown to yellowish green.
Facies Analysis
Vertical and lateral facies distribution at the two studied sections allows recognition of three facies associations (Figs. 2 and 3). Facies Association 1 (FA1): Siltstones and fine sandstones rich in pyroclastic material and clumps, lens and beds of skeletal fossils concentrations. Facies Association 2 (FA2): Highly bioturbated siltstones and tuffs with concretions. Facies Association 3 (FA3): Interstratified siltstones, fine sandstones and claystones, thinly laminated in some sectors.
Facies Association 1 (FA1)
This facies association is roughly equivalent to Darwin's (Darwin 1846, p. 112) lowermost stratum (i.e., "… to a thickness of fifty or sixty feet, consists of a more or less hardened, darkish, muddy, or argillaceous sandstone…"). It includes yellowish grey siltstones and fine muddy sandstones with abundant pyroclastic material and skeletal concentrations forming clumps, lens or beds, dominated by "Turritella" patagonica (Fig. 4a). Also intercalated are specimens of Crassostrea? hatcheri in life position or slightly rotated but still with both valves articulated. This facies association also carries most of the rich and diverse fauna of the Monte León Formation (see description of Sections for a list of the most conspicuous taxa and their stratigraphic distribution). Also present are abundant bioturbations as small tubes running in all directions, and galleries assigned to Ophiomorpha isp. Concretions of variable size and shape are abundant, sometimes joined together forming "necklaces". This facies association measures 30 m thick at Darwin's Section and 17 m thick at Mount Entrance Section.
 Figure 4: a) Clumps of "Turritella" patagonica (FA1); b) Ophiomorpha isp. (FA2); c) Concretions (FA2); d) Skeletal concentration dominated by Dosinia laeviscula at the top of FA2.
Figure 4: a) Clumps of "Turritella" patagonica (FA1); b) Ophiomorpha isp. (FA2); c) Concretions (FA2); d) Skeletal concentration dominated by Dosinia laeviscula at the top of FA2.
Some of the skeletal concentrations occur in laterally continuous beds, with wavy contacts and hardened concretionary sectors that form ledges in the section. There are a least three beds 1 - 2.5 m thick. The shell beds are densely to loosely packed (25%) in some of the sectors, and densely packed (55%) in others. Size-sorting is poor; although by far the numerically most abundant taxon is "T." patagonica; it is represented by specimens of all sizes. Orientation is chaotic, disarticulation is very high and only a few shells are conjoined. Fragmentation is low in some sectors and high in others, while abrasion is nil. Encrusting and bioerosion are evident in valves of C.? hatcheri, which carry encrusting bryozoans, sponge borings (Entobia isp.), polychaetes (Maeandropolydora isp.) and bivalve borings (Gastrochaenolites isp.). In addition to the aforementioned shell beds also common are lenses of between 0.05 and 0.3 m thickness. The fossil content of these lenses is the same as the beds, as are all taphonomic attributes too. The uppermost part of this facies association shows a sharp decrease in fossil content, as the beds and lenses become much less frequent. However, a few disperse fossils such as a Dentalium in life position and Limopsis insolita can be observed. Facies Association 1 represents a transition environment between the inner shelf and the subtidal part of a tidal plain, below fair weather wave base but above storm wave base. The mollusks contained in these beds are commonly considered to be indicative of shallow, fully marine conditions with normal salinity. Well over half the taxa present in this fauna are either very shallow infaunal (Turritella, Cucullaea, Cardium, etc) or epifaunal (Zygochlamys, Reticulochlamys, Swiftopecten, Trophon, Austrotoma, etc.). Most trophic levels are represented, including suspension feeders, deposit feeders and carnivores. Taxonomic composition of the assemblage and the beds with articulate and highly bored oyster clusters suggest that the background sedimentation rate was low. The presence of thin consolidated lenses with fragments of shells or specimens lying chaotically is interpreted as of deposits formed by sporadic events recording a sharp increase in current velocity. The lenticular structure of some of these skeletal concentrations and their taphonomic features suggests they belong to the infilling of small scale tidal channels in a subtidal to inner shelf environment.
Facies Association 2 (FA2)
This facies association overlies FA1 and is equivalent to Darwin's lower two strata of the upper part of the cliff. It comprises massive and bioturbated tuffaceous siltstones and very fine sandstones, and tuffs. These are grey at the base and light yellow towards the top. The thickness is approximately 27 m at Darwin's section and 22 m at the Mount Entrance Section. The base carries large galleries assigned to Ophiomorpha isp. (Fig. 4b) and also few isolated shell fragments. Towards the top there are frequent galleries assigned to Thalassinoides isp. (up to 8 cm diameter). It also carries very sparse remains of crabs and small monospecific clumps of a small bivalve possibly referable to Pteromyrtea, and very few fragments of larger bivalve shells. The uppermost 8 m carry a few isolated valves of Dosinia laeviuscula. These are disarticulate, convex- up, and mostly unbroken. Throughout this facies association are common concretions of 0.8 - 1.4 m diameter that may be isolated or joined forming "necklaces" (Fig. 4c). Darwin had already noticed them as a conspicuous feature in this part of the section: "…Of the three beds, the central one is the most compact, and more like ordinary sandstone: it includes numerous flattened spherical concretions, often united like a necklace, composed of hard calcareous sandstone, containing a few shells: some of these concretions were four feet in diameter, and in a horizontal line nine feet apart…"(Darwin 1846, p. 113). At the Mount Entrance Section there is a hard ledge of about 0.8 m thickness with cross-bedding. It is highly bioturbated with numerous tubes of about one cm diameter placed obliquely to stratification. At the Darwin Section this facies association ends with a monospecific shell concentration of 0.2 - 0.5 m thickness, with a very fine sandstone matrix, and containing adult specimens of Dosinia laeviscula (Fig. 4d). Contacts of this bed are wavy and bioturbated, with many tubes reaching up to 0.7 m down into the underlying bed. Laterally it gives way to a white tuff of equal thickness. Close-packing is dense (55%), size-sorting good, orientation chaotic, disarticulation high, fragmentation moderate to low, abrasion nil. At the Mount Entrance Section this facies association ends with a skeletal concentration of about 0.6 m thickness, also with a fine sandstone matrix, but with a polyspecific taxonomic composition, carrying an abundant fauna of gastropods and bivalves (see description of Sections for a list of the most conspicuous taxa and their stratigraphic distribution). The lower contact is wavy and the upper one transitional, as close-packing is dense at the base and dispersed at the top. Size-sorting is good, as the whole specimens and the shell fragments are of the same size with only a few interspersed large bivalves and gastropods. Orientation is chaotic, disarticulation is complete, and fragmentation of large specimens is high but with a few whole specimens, while small specimens are largely unbroken. Many specimens show muricid and naticid boreholes. Facies association 2 represents a subtidal environment at the lowest part of a tidal plain. The low diversity in the fauna throughout the facies association (Pteromyrtea and Dosinia), suggests a more marginal shallow environment than FA1. The high degree of bioturbation and dominance of trace fossils assigned to Thalassinoides isp. and Ophiomorpha isp. suggests low sedimentation rate and an environment with moderate to relatively low energy conditions. Taphonomic features of the specimens in the skeletal concentrations at the top, together with the presence of apparently unselected material suggests that these concentrations were formed during a high energy event, perhaps representing a storm event.
Facies Association 3 (FA3)
This facies association comprises the uppermost bed of Darwin's "upper part of the cliff" (Darwin 1846, p. 113). It includes 19 - 20 m of interstratified siltstones, fine sandstones and claystones, sometimes with heterolithic lamination. At the base there are convex-down right valves of Crassostrea? hatcheri. It carries a few concretions with bivalves and echinoderms. At 11 m above the base color changes abruptly from brown to yellowish green. Fine grain-size and sedimentary structures produced by variations between calm and turbulent water indicates deposition by combined traction and decantation processes. Such a process suggests that this facies association developed in a subtidal to intertidal environment.
Paleonvironments
Facies associations together with the invertebrate fauna recorded in each bed allow a detailed paleoenvironmental reconstruction of the rocks of the Monte León Formation in the Río Santa Cruz area. These rocks can be included in the regressive stage of the high sea-level period that began at the end of the Oligocene. The proposed deposition model suggests that it took place in a shallow sea, with variations that range from the inner shelf to subtidal or even intertidal environments. At the base of the section (FA1) - i.e., the inner shelf to subtidal environment - there was a continuous deposition of siltstone with volcanic material (bioturbated) as result of decantation, with a few intercalated episodes recording higher energy events. These were recorded as beds and lenses of skeletal concentrations interpreted as having been deposited by channeled high energy unidirectional currents, perhaps tidal channels. Towards the top of the sections (FA2) these lenses become sparser, while the bioturbation and pyroclastic content of the deposit increase. The top carries finely laminated siltstones and sandstones (FA3) that suggest a subtidal - or even intertidal - environment.
PALEONTOLOGY
One of the most conspicuous features of the Monte León Formation is the rich and excellently preserved mollusk fauna contained in the beds and lenses within the Punta Entrada Member; it occurs mainly in FA1 and the most conspicuous and common elements are mentioned above. This fauna has been known ever since Darwin collected the first specimens at the mouth of the Río Santa Cruz, and has been used over the years in the formulation of diverse hypothesis on the biostratigraphic and paleobiogeographic distributions of the taxa involved (del Río 2004 and references therein). Darwin's initial collection of 16 species of mollusks has been increased to over 110 recognized from that locality nowadays. Of these, virtually all are also known from the shell beds that were described later by Ameghino and Hatcher. The type locality of the species described by Sowerby (1846) is the section described here as Darwin Section. Thus, all his taxa come from beds lying within the Punta Entrada Member. It is important to note that taxa later described by von Ihering (1897, 1907, 1914) and Ortmann (1902) came from further south. The beds bearing these taxa were then included in the Superpatagonian, although Hatcher and Ortmann could see no difference between the Patagonian and Superpatagonian fauna. This is not surprising, as the Superpatagonian included two suites of taxa from entirely different sources. In the first place it included the species that is unique to the lowermost beds of the Santa Cruz Formation, i.e., Crasssotrea orbignyi. This oyster is quite different from Crassostrea? hatcheri, and is widespread in Patagonia and always appears associated to the marginal marine rocks at the base of the Santa Cruz Formation or its equivalent units. Secondly, the Superpatagonian included many taxa common to the "Leonense" chronostratigrapic unit of Ameghino, together with a suite of exclusive taxa not known from the underlying strata. However, these were based on material coming from two different sources. In the first place, it included material collected from the shell beds at the top of the Punta Entrada member, at localities where this unit is exposed along the coast between the mouth of the Río Santa Cruz and Monte Observación, just off limits to the south of Monte León National Park. It also includes material collected from beds overlying the former and containing Crassostrea orbignyi, exposed a few kilometers further inland and clearly at the base of the Santa Cruz Formation. The taphonomic attributes of the material other than the autochtonous accumulations of C. orbignyi suggest that it is in all cases reworked material from the underlying shell beds. In most cases, taxonomic differences can be attributed to the differential preservation of the two suites of material.
MEANING OF THE GREAT PATAGONIAN FORMATION AT THE MOUTH OF THE RÍO SANTA CRUZ
Upstream the Río Santa Cruz, the rocks described by Darwin at the mouth of the river are gradually replaced by continental beds nowadays included in the Santa Cruz Formation (Darwin 1846, p. 113-115). Although he described a section at about 160 km west of Santa Cruz, Darwin was doubtful that the rocks could be included in "the great Patagonian tertiary formation", as the only fossils he could find were very worn pieces of Ostrea patagonica (i.e. Crassostrea? hatcheri or more likely C. orbignyi). As Darwin never went ashore further south than Mount Entrance, he was unable to observe neither the beds at Las Cuevas nor those within the Monte Observación Member. Therefore, his concept of "great Patagonian tertiary formation" or "Patagonian Tertiary Formation" only included those rocks exposed at Santa Cruz (and other places further north). At Mount Entrance Bertels (1980, p. 214) defined a lower member of the Monte León Formation that she named Punta Entrada Member and included in it all the marine beds exposed there. Darwin's Section is only 2.6 km away and no major structural features alter the stratigraphic relations between the beds exposed at the two localities. Consequently, we can confidently assume that (as confirmed by descriptions above) the two sections are equivalent. About 40 km further south along the coast and within the present Monte León National Park, these rocks continue to form part of the coastal cliff and lithological features remain overall constant. However, a short distance inland a second higher ledge is composed mainly by the continental Santa Cruz Formation. The contact between the two units has been subject of heated debates over the years, ever since the first subdivisions of the Patagonian Tertiary were made by Ameghino. The Patagonian Tertiary Formation in Darwin's sense was restricted by Ameghino (1894) to the marine beds exposed along the coast between Chubut and Monte Observación in Santa Cruz. His subdivision of this unit (Ameghino 1898) into "Juliense" and "Leonense" was followed shortly afterwards (Ameghino 1900-1902) by his recognition that the overlying "Formación Santacruceña" included a lower marine bed he called "Superpatagoniano". In this he included the shell beds at the top of the cliff exposed at Las Cuevas, Yegua Quemada, Jack Harvey and Monte Observación, and also the oyster beds at the base of the Santa Cruz Formation exposed further inland at Cerro Monte León and Cerro Observación. He based such a subdivision on the fossil content, a fact that was later disputed by other authors (Wilckens 1905, Hatcher 1900, Ortmann 1902) who could see no difference in taxonomic composition between the Superpatagonian beds and those appearing in the typical Patagonian Formation. Di Paola and Marchese (1973) did not recognize Bertels' introduction of a formational status for Ameghino's Juliense and Leonense (i.e., her San Julián and Monte León Formations). Instead they subdivided the Patagonia Formation (sensu Ameghino 1894) into three members based on lithological differences among them. The uppermost one, overlying the Monte León Member, they named Monte Observación Member. They defined this unit as the rocks appearing below the Santa Cruz Formation at Cerro Observación and that showed features common to the Monte León Member at the base and to the Santa Cruz Formation at the top. The bottom of this member is - according to them - an oyster bed of Crassostrea orbignyi, and the top is the last bed of this same oyster before giving way completely to the continental mammal-bearing rocks of the Santa Cruz Formation (the contact here is transitional). The total thickness measured by them between the two oyster beds was 27 m. When introducing her Punta Entrada Member, Bertels (1980, p. 214) also acknowledged the difference between this unit and the topmost beds of the Monte León Formation. Therefore she admitted the formal status given by Di Paola and Marchese (1973, p. 214) to the Monte Observación Member. However, she stated that a) the Monte Observación Member included all the shell beds at the top of the sections at Monte León (Cabeza de León), Las Cuevas, and Monte Observación (i.e., Cerro Observación, just west of Las Cuevas), and b) it replaced Ameghino's chronostratigraphic term "Piso Superpatagónico". However, these assumptions appear to be unwarranted. The lithological features described by Di Paola and Marchese (1973) for the Monte Observación Member are quite clearly distinct from those observed (as noted by Bertels herself; Bertels 1980, p. 216) in the shell beds at the top of the sections at Monte León (Cabeza de León) and Las Cuevas. The lithology in these shell beds is clearly the same as that in the underlying beds attributed by her to her Punta Entrada Member. The Superpatagonian of Ameghino originally included not only the shell beds at Monte León and Las Cuevas, but also those at Yegua Quemada, Jack Harvey, and Monte Observación all of them south of Las Cuevas. He also included the shell beds with Crassostrea orbignyi at Cañadón de los Artilleros, inland from San Julián, also underlying the continental Santa Cruz Formation. Neither Ameghino nor any author after him considered the fact that the shell beds at these places are thicker, with a far more abundant and better preserved fauna than those at the mouth of the Río Santa Cruz, the most likely reason for them supporting the difference between the Patagonian and Superpatagonian beds. Such a difference was minimized by Ortmann (1902) and Hatcher (1900) based on the very abundant fauna from Santa Cruz (possibly near Mount Entrance) that they had available. In fact, these authors were right in stating that the different composition of the fauna was due to local facies variations. Moreover, the far more abundant fauna at Las Cuevas is also surely the cause of bias in this sense, as the chances of collecting uncommon taxa are naturally much higher there. Di Paola and Marchese (1973), while correctly defining their Monte Observación Member, never mentioned the fact that between the base of this unit (i.e., the lowest oyster bed at Cerro Observación) and the top of the shell beds at Las Cuevas there are ca. 20 m of rocks that are not exposed anywhere between Santa Cruz and the last exposures with marine shells at Puerto Coyle. As the lithology (and sedimentary structures as well) of the Monte Observación Member (other than the in situ beds with oysters in life position) is identical with that of the Santa Cruz Formation and the beds with other marine shells included in this unit are clearly reworked material with taxa that appear at Las Cuevas too, we suggest that the Monte Observación Member should be restricted to the rocks described by Di Paola and Marchese (1973, p. 214) and included in the Santa Cruz Formation. Likewise, the shell beds at the top of the sections at Monte León (Cabeza de León) and Las Cueva, Yegua Quemada, Jack Harvey and Monte Observación should be definitely referred to the Monte León Formation as they are equivalent to the shell beds lying within the Punta Entrada Member at Mount Entrance and Darwin's Section. Therefore, the use of the term Superpatagonian becomes superfluos.
CONCLUSIONS
As a result of this study we were able to pinpoint the most probable location where Charles Darwin made his observations in the Río Santa Cruz mouth. At this locality, i.e., our Darwin Section, we also identified the beds mentioned by him, including the location of the beds from where he collected the fossil mollusks that were described by Sowerby. The strata recognized by him are equivalent to those described by us and of which a detailed lithological description is provided. Rocks belonging to the Monte León Formation exposed at the mouth of the Río Santa Cruz can be included in the regressive stage of the Atlantic transgressive-regressive cycle that flooded Patagonia during the late Oligocene-early Miocene. The deposition model suggests that it took place in a shallow sea, with variations that range from the inner shelf to subtidal or even intertidal environment. Analysis of sedimentological features of these rocks, together with the mollusk fauna contained in them, and comparison with those from nearby exposures further south along the coast and traditionally referred to the "Superpatagonian", suggest that the Monte Observación Member of the Monte León Formation should be restricted to its original concept. As a consequence, the term "Superpatagonian" becomes meaningless, at least as a chronostratigraphic unit in the area studied.
ACKNOWLEDGEMENTS
We would like to thank Sergio Martínez (Montevideo, Uruguay) and Jeff Stilwell (Melbourne, Australia) for their thoughtful comments that improved the original version of this manuscript. Help in the field by Juan Griffin is duly acknowledged too. The Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa supported part of the fieldwork (Project PI188). CONICET financed field and lab work (Project PIP1442 /05).
WORKS CITED IN THE TEXT
1. Ameghino, F. 1894. Ennumération synoptique des espèces des mammifères fossiles des formations eocenes de Patagonie. Boletín de la Academia Nacional de Ciencias 13: 259-445, Córdoba.
2. Ameghino, F. 1896. Notas sobre cuestiones de geología y paleontología argentina. Boletín del Instituto Geográfico Argentino 17: 87-119.
3. Ameghino, F. 1898. Sinopsis Geológico-Paleontológica. Segundo Censo de la República Argentina 1: 111-225.
4. Ameghino, F. 1900-1902. L'age des formations sédimentaires de Patagonie. Anales de la Sociedad Científica Argentina 50: 109-130, 145-165, 209-229 (1900); 51: 20-39, 65-91 (1901); 52: 189-197, 244-250 (1901); 54: 161-180, 220-249, 283-342 (1902).
5. Barreda, V. and Palamarczuk, S. 2000. Palinomorfos continentales y marinos de la Formación Monte León en su área tipo, provincia de Santa Cruz, Argentina. Ameghiniana 37(1): 3-12.
6. Becker, D. 1964. Micropaleontología del Superpatagoniense de las localidades Las Cuevas y Monte Entrance (provincia de Santa Cruz). Ameghiniana 3(10): 319-351.
7. Bertels, A. 1970. Sobre el "Piso Patagoniano" y la representación de la época del Oligoceno en Patagonia austral, República Argentina. Revista de la Asociación Geológica Argentina 25(4): 495-501.
8. Bertels, A. 1980. Estratigrafía y foraminíferos (Protozoa) bentónicos de la Formación Monte León (Oligoceno) en su área tipo, provincia de Santa Cruz, República Argentina. Actas 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires, 1978) 2: 213- 273.
9. Camacho, H. H. 1974. Bioestratigrafía de las formaciones marinas del Eoceno y Oligoceno de la Patagonia. Anales de la Academia de Ciencias Exactas, Físicas y Naturales 26: 39-57.
10. Camacho, H. H. 1979. Significados y usos de "Patagoniano", "Patagoniense", "Formación Patagonica", "Formación Patagonia" y otros términos de la estratigrafía del Terciario marino argentino. Revista de la Asociación Geológica Argentina 34(3): 235-242.
11. Cossmann, M. 1899. Description des quelques coquilles de la formation Santacruzienne en Patagonie. Journal de Conchiliologie 47(3): 223-241.
12. Darwin, Ch. 1839. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle, between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Colburn, H. Ed., London, vol. 3, 615 p.
13. Darwin, Ch. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R. N. during the years 1832 to 1836. Smith Elder and co Ed., London, 279 p.
14. del Río, C. J. 2004. Tertiary marine Molluscan Assemblages of Eastern Patagonia (Argentina): a biostratigraphic analysis. Journal of Paleontology 78(6):1097-1122.
15. del Río, C. J. and Camacho, H. H. 1998. Tertiary nuculoids and arcoids of eastern Patagonia (Argentina). Palaeontographica Abt. A, 250: 47-88.
16. Di Paola, E. C. and Marchese, H. G. 1973. Litoestratigrafía de la Formación Patagonia en el área tipo (Bajo de San Julián-desembocadura del río Santa Cruz). Provincia de Santa Cruz. República Argentina. 5º Congreso Geológico Argentino, Actas 3: 207-222.
17. d'Orbigny, A. 1842a. Voyage dans l'Amérique Méridionale (le Brésil, la République Orientale de l'Uruguay, la République Argentine, la Patagonie, la République du Chili, la République du Bolivia, la République du Perou), exécuté pendant les années 1826,1827,1828,1829, 1830,1831, 1832 et 1833. P. Bertrand. Paris, V. Levrault, Strasbourg, vol. 3, part. 3, 290 p.
18. d'Orbigny, A. 1842b. Voyage dans l'Amérique Méridionale (le Brésil, la République Orientale de l'Uruguay, la République Argentine, la Patagonie, la République du Chili, la République du Bolivia, la République du Perou), exécuté pendant les années 1826,1827,1828,1829, 1830,1831, 1832 et 1833. P. Bertrand. Paris, V. Levrault, Strasbourg, vol. 3, part. 4, 96 p.
19. Feruglio, E. 1949. Descripción Geológica de La Patagonia. Ministerio de Industria y Comercio de La Nación, Dirección General de Yacimientos Petrolíferos Fiscales 2: 1-349.
20. Fleagle, J.G., Bown, T.M., Swisher, C. and Buckley, G. 1995. Age of the Pinturas and Santa Cruz Formations. 6º Congreso Argentino de Paleontología y Bioestratigrafía (Trelew), Actas: 129-135.
21. Hatcher, J.B. 1897. On the geology of Southern Patagonia. American Journal of Science 4(23): 327-354
22. Hatcher, J. B. 1900. Sedimentary rocks of Southern Patagonia. American Journal of Sciences (4) 9(50): 85-107.
23. Hatcher, J.B. 1903. Geography of Southern Patagonia. In Scott, W.B. (ed.) Reports of the Princeton University Expedition to Patagonia 1896-1899, J. Pierpoint Morgan Publishing Foundation, Narrative of the expeditions 1: 45-332, Princeton.
24. Ihering von, H. 1897. Os Molluscos dos terrenos terciarios da Patagonia. Revista do Museo Paulista 2: 217-382.
25. Ihering von, H. 1899. Die Conchylien der patagonischen Formation. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1899: 1-46.
26. Ihering von, H. 1902. Historia de las ostras argentinas. Anales del Museo Nacional de Buenos Aires 7: 109-123.
27. Ihering von, H. 1907. Les mollusques fossiles du Tertiaire et du Crétacé superieur de l'Argentine. Anales del Museo Nacional de Buenos Aires, serie 3(7): 1-611.
28. Ihering von, H. 1914. Catálogo de molluscos Cretáceos e Terciarios da Argentina da collecçao do auctor [with appendix in German]. Notas Preliminares da Revista do Museu Paulista 1(3): 1-148.
29. Kidwell, S. M. and Holland, S. M. 1991. Field description of coarse bioclastic fabrics. Palaios 6: 426-434.
30. Kidwell, S. M., Fürsich, F. T. and Aigner, T. 1986. Conceptual Framework for the Analysis and Classification of Fossil Concentrations. Palaios 1: 228-238.
31. Legarreta, L. and Uliana, M. A. 1994. Asociaciones de fósiles y hiatos en el Supracretácico-Neógeno de Patagonia: una perspectiva estratigráfico-secuencial. Ameghiniana 31(3): 257-281.
32. Malumián, N. 1999. La sedimentación y el volcanismo terciarios en la Patagonia extraandina. In Caminos, R. (ed.) Geología Argentina, Anales del Instituto de Geología y Recursos Minerales 29(18): 557-612.
33. Malumián, N. 2002. El Terciario marino. Sus relaciones con el eustatismo. In Haller, M.J. (ed.) Geología y Recursos Naturales de Santa Cruz, 15º Congreso Geológico Argentino, Relatorio I-15: 237-244.
34. Náñez, C. 1988. Foraminíferos y bioestratigrafía del Terciario Medio de Santa Cruz oriental. Revista de la Asociación Geológica Argentina 43(4): 493-517.
35. Olivera, A. M. 1990. Aspectos paleoecológicos de asociaciones de moluscos fósiles de la Formación Monte León (Oligoceno). Puerto Santa Cruz (Santa Cruz). 5º Congreso Argentino de Paleontología y Biostratigrafía, Actas 1: 155-160.
36. Ortmann, A. E. 1902. Tertiary Invertebrates. In Scott, W. B. (ed.) Reports of the Princeton University Expedition to Patagonia 1896-1899, J. Pierpoint Morgan Publishing Foundation, Paleontology volume 4(I), Part 2: 45- 332, Princeton.
37. Panza, J. L., Irigoyen, M. V. and Genini, A. 1995. Hoja Geológica 4969-IV, Puerto San Julián, provincia de Santa Cruz, República Argentina. Boletín de la Secretaría de Minería de la Nación, Dirección Nacional del Servicio Geológico 211: 1-77.
38. Parras, A. and Casadío, S. 2005. Taphonomy and sequence stratigraphic significance of oysterdominated concentrations from the San Julián Formation, Oligocene of Patagonia, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 217: 47-66.
39. Parras, A., Griffin, M., Feldmann, R., Casadío, S., Schweitzer, C. and Marenssi, S. 2008. Correlation of marine beds based on Sr- and Ar- date determinations and faunal affinities across the Paleogene/Neogene boundary in southern Patagonia, Argentina. Journal of South American Earth Sciences 26: 204-216.
40. Philippi, R. A. 1887. Los fósiles terciarios i cuartarios de Chile. F. A. Brockhaus, Leipzig and Museo Nacional de Historia Natural, 256 p., Santiago.
41. Riggi, J. C. 1978. La importancia de los sedimentos piroclásticos y de la sílice biogenética en la estratigrafía de la Formación Patagonia. Revista de la Asociación Geológica Argentina 33(2): 158-171.
42. Sowerby, G. B. 1846. Descriptions of the Tertiary fossil shells from South America. In Darwin, Ch. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R. N. during the years 1832 to 1836. Smith Elder and Co. Ed., Appendix, p. 249-264, London.
43. Wichmann, R. 1922. Observaciones geológicas en el Gran Bajo de San Julián y sus alrededores (Territorio de Santa Cruz). Boletín Dirección General de Minas 30B: 1-34.
44. Wilckens, O. 1905. Die Meeresablagerugen der Kreide und Tertiärformation in Patagonien. Neues Jahrbuch für Geologie, Mineralogie und Paläontologie 21: 98-195.
45. Windhausen, A. 1931. Geología Argentina. Segunda Parte: Geología Histórica y Regional del territorio Argentino. Editorial Peuser, 676 p., Buenos Aires.
46. Zinsmeister, W. J. 1981. Middle to late Eocene invertebrate fauna from the San Julián Formation at Punta Casamayor, Santa Cruz province, Southern Argentina. Journal of Paleontology 55: 1083-1102.
Silvio Casadío and Miguel Griffin
Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Santa Rosa, La Pampa. Email: miguelgriffin@aol.com
ABSTRACT
Rocks exposed just south of Puerto Deseado (Port Desire), Santa Cruz Province, were surveyed by Darwin during his journey on board HMS Beagle. The fossil mollusks collected there were studied later by Sowerby, who described four species based on the material from "Port Desire". Sedimentological and stratigraphical observations suggest that the marine rocks cropping out at Darwin's locality should be assigned to the Monte León Formation (early Miocene). The rocks were deposited in subtidal environments, most precisely in an offshore environment near fair weather wave base at the bottom to a lower shoreface at the top of the section.
Keywords: Darwin; Early Miocene; Monte León Formation; Sedimentology; Paleontology.
RESUMEN: Sedimentología y paleontología de una sucesión marina miocena citada por primera vez por Darwin en Puerto Deseado. Las rocas expuestas al sur de Puerto Deseado, provincia de Santa Cruz, fueron relevadas por Darwin durante su viaje a bordo del HMS Beagle. Los moluscos fósiles que coleccionó fueron estudiados por Sowerby quien describió cuatro especies basadas en ejemplares procedentes de Puerto Deseado. Las observaciones sedimentológicas y estratigráficas sugieren que estos depósitos marinos, estudiados originalmente por Darwin, son asignables a la Formación Monte León (Mioceno temprano). En general, las rocas representan ambientes submareales que en el tramo inferior de la sucesión corresponden a un costa afuera, próximo al nivel de base de olas de buen tiempo, mientras que hacia el techo pasan a un shoreface inferior.
Palabras clave: Darwin; Mioceno temprano; Formación Monte León; Sedimentología, Paleontología.
INTRODUCTION
The Cenozoic marine deposits of Patagonia have been known since the early 19th century and have been studied ever since d'Orbigny (1842) first described his Tertiaire Patagonien. This author also described the fossils that he collected during his explorations in southern South America. All of the specimens he described came from the marine rocks exposed in northern Patagonia - in the vicinity of the mouth of the Río Negro, and in the area surrounding the present city of Paraná, in northeastern Argentina. Likewise, he did mention the presence of a large oyster similar to Crassostrea patagonica (d'Orbigny 1842) in Puerto San Julián, which had been given to him by a sailor. However, he never actually visited the coast further south of Río Negro. During his journey to Chile, Bolivia and Peru around Cape Horn, he must have seen from afar the cliffs rising along the coast, as he included all these exposed rocks in his Tertiare Patagonien. A few years later, HMS Beagle sailed along the coast of Patagonia, and on this occasion the geology of the region was studied by Charles Darwin, who published his observations (Darwin 1846) together with descriptions of the fossil taxa he collected during this trip (Sowerby 1846). In addition to the localities already mentioned by d'Orbigny, Darwin visited four other places that became classic fossiliferous localities over the years, and that were crucial in establishing the basis of Patagonian Cenozoic stratigraphy. These four localities are San José, Puerto Deseado, Puerto San Julián, and Santa Cruz. San José lies on the northern coast of Península Valdés in Chubut (northern Patagonia). The other three localities lie much further south along the coast of the present province of Santa Cruz. Darwin included all the rocks with marine fossils from these localities, and from Paraná and Uruguay, in what he called the Great Patagonian Formation. Although this appears to be roughly equivalent to d'Orbigny's Tertiaire Patagonien, Darwin was aware that this succession included two distinct units, the lower one marine, and the second one, continental with land vertebrate remains. Nowadays, the four localities visited by Darwin include different formal units, deposited in different basins and environments. For example, the marine deposits at San José locality belong to the Puerto Madryn Formation (late Miocene). The marine rocks and fossils mentioned by Darwin at San Julián belong either to the late Oligocene San Julián Formation (Bertels 1970), or to the early Miocene Monte León Formation (Bertels 1970). At the mouth of the Río Santa Cruz, the rocks described by Darwin belong to the Monte León Formation. Darwin's observations and fossil collections drew the interest of later explorers, geologists and paleontologists alike, especially during the second half of the 19th century and early decades of the 20th century. During this period, Ameghino (1896, 1898, 1900-1902) established the earliest stratigraphic subdivision of the Cenozoic rocks of Patagonia. This subdivision was mostly based on the fossil content of these richly fossiliferous beds. However, since Ameghino's subdivision, there has been a lot of controversies, not only because of the age of the rocks, but also because of the stratigraphic relationships and correlations of several outcrops that lie spread over a huge area in Patagonia, and that were attributed to the "Patagonian beds". Most of the rocks exposed there are now included in different formal units, although the exact relationships among them are still partially unresolved (Parras et al. 2008). Detailed history of these controversies can be found in Camacho (1974, 1979), Zinsmeister (1981), Legarreta and Uliana (1994), and Malumián (1999, 2002). Many of the rich faunas contained in the marine and continental Cenozoic rocks are crucial in the reconstruction of the evolutionary history of the South American biota. Therefore, a clear understanding of the stratigraphic relationships - especially for those type localities of taxa that are used with biostratigraphic and/or paleobiogeographic purposes - is prerequisite to any further inference involving these rocks and the fossils that they contained. During its voyage around the world, the HMS Beagle anchored in the Río Deseado mouth. There, Darwin had the opportunity to explore the riverbanks upstream and to provide detailed descriptions of the rocks he encountered. The northern bank of the river - where the modern town of Puerto Deseado (Fig. 1) lies and along where the road and railroad run - is mainly composed of Jurassic volcanic rocks for at least 30 km upstream. These rocks were well recorded by Darwin, who was aware of their lack of fossils. However, a few kilometers further upstream of "Darwin's Rock", the furthest upstream point reached by him, marine Cenozoic rocks rich in fossils become exposed.
 Figure 1: Location map, showing Darwin's locality at Port Desire
Figure 1: Location map, showing Darwin's locality at Port Desire
These rocks remained unknown to Darwin. Nevertheless, among the fossils collected by him and described by Sowerby (1846), there are several that come from "Port Desire". Darwin mentioned as the fossiliferous locality, the cliffs lying a few miles south of the river mouth. This locality comprises two very small outcrops that ever since remained almost unexplored by geologists or paleontologists, as they are in a relatively inaccessible area and are not visible from the town. Four of the species of mollusks described by Sowerby come from this locality. Despite the fact that it is a type locality of taxa later used in biostratigraphic and paleobiogeograpic reconstructions, it has remained obscure and little known ever since Darwin discovered it. The late Oligocene - early Miocene marine beds exposed along the Atlantic coast of Patagonia are presently included in the San Julián (late Oligocene - early Miocene) and Monte León (early Miocene) formations (in the Austral Basin), and in the Chenque Formation (in the San Jorge Basin). These rocks were deposited during Atlantic transgressions that covered large portions of Patagonia. The Monte León Formation was formally described by Bertels (1970), and it includes those deposits at the mouth of the Río Santa Cruz, which are characterized by their abundant and well preserved fauna. These rocks and faunas were extensively studied throughout the years (Ihering 1897, 1899, 1907, 1914, Ortmann 1902, Feruglio 1949, Di Paola and Marchese 1973, Bertels 1980, Náñez 1988, del Río 2004, del Río and Camacho 1998, Crawford et al. 2008, and references therein), but most of these studies were carried out on the exposures found around the mouth of the Río Santa Cruz and in the San Julián area. In contrast, outcrops with similar rocks in the Puerto Deseado area received far less attention (Frenguelli 1931, Feruglio 1949, del Río 2004, Giacosa et al. 1998). In this area, the Neogene marine rocks overlie Jurassic volcanic rocks of the Bahía Laura Group, and are reduced to patchy thin exposures isolated one another. Giacosa et al. (1998) used the informal name "Patagoniano" to designate these outcrops and suggested that these were equivalent to the San Julián and Monte León formations. However, del Río (2004) included them in the Chenque Formation, a unit formally described for rocks exposed about 150 km to the northwest, in the vicinity of Comodoro Rivadavia city. Sedimentological and stratigraphical observations suggest that the Neogene marine rocks at Darwin's locality near Puerto Deseado should be assigned to the Monte León Formation. In this study we present a detailed description of these rocks and comments on its paleontological content. Institutional abbreviations used in this work are as follows: NHM-L (The Natural History Museum, London, Department of Palaeontology); GHUNLPam (Colección Paleontológica del Departamento de Ciencias Naturales de la Universidad Nacional de La Pampa).
GEOLOGICAL SETTING
South of Puerto Deseado, the Monte León Formation (early Miocene) is exposed between Punta Guanacos and Punta Norte. The lowermost portion of the section is exposed during low tide along the coast, with the remainder of the formation outcropping along the base of cliffs. At the studied locality (47º48'37''S; 65º52'32''W, see Fig. 1), the Monte León Formation is 10 m thick (Fig. 2) and unconformably overlies Jurassic volcanic rocks of the Bahía Laura Group. The contact between the two units is only visible during low tide. The Monte León Formation is unconformably overlain by Quaternary conglomerates (Fig. 3a).
 Figure 2: Stratigraphic section at Darwin's locality.
Figure 2: Stratigraphic section at Darwin's locality.
 Figure 3: a) Puerto Deseado section. The Monte León Formation is unconformably overlain by Quaternary conglomerates; b) Lenticular shell concentrations dominated by "Turritella" patagonica; c) Specimens - mostly articulated - of Crassostrea? hatcheri; d) Thalassinoides isp. Filled with alternating sandy and muddy laminae.
Figure 3: a) Puerto Deseado section. The Monte León Formation is unconformably overlain by Quaternary conglomerates; b) Lenticular shell concentrations dominated by "Turritella" patagonica; c) Specimens - mostly articulated - of Crassostrea? hatcheri; d) Thalassinoides isp. Filled with alternating sandy and muddy laminae.
Facies associations (FA)
FA1: This association includes fine bioturbated sandstone and mudstone, intercalated with lenticular shell concentrations (less than one meter wide and 0.4 m thick) with trough-cross stratification, dominated by "Turritella" patagonica (Fig. 3b). At the top of these concentrations there are specimens - mainly articulated - of Crassostrea? hatcheri (Fig. 3c). Specimens of Crassostrea? hatcheri in life position are also abundant. They are mainly obvious du-ring low tide on the wide abrasion platform, where some specimens are found forming clumps. Commonly, the valves of this oyster are bioeroded by sponge (Entobia isp.), polychaetes (Maeandropoly-dora isp.) and Pholadidea patagonica (Gastro-chaenolites isp.). Large burrow systems, with horizontal and vertical components are frequent in this facies association. These excavations are 5 cm in average diameter, and are assigned to Thalassinoi-des isp. The fill of these structures consists of alternating snady and muddy laminae (Fig. 3d).
Interpretation FA1 represents an environment lying above storm wave base and below normal wave base. The alternating sandy and muddy laminae filling the Thalassinoides burrows probably indicate tidal cyclicity. The abundance of Thalassinoides isp. suggests that the benthos was well-oxygenated during deposition (Bottjer et al. 1986, Bromley 1990) and the substrate was probably firm. However, the abundance of "Turritella" patagonica - member of a very shallow infaunal group - suggests that the surface sediments were most likely poorly consolidated. The concentrations of "Turritella" patagonica may represent the infilling of small tidal channels. The beds with articulated and highly bored oyster clusters point towards periods of lower sedimentation rates. FA2: comprises mainly fine- to mediumgrained massive tuffaceous sandstones. Bioturbation completely obliterated the sedimentary structures. There are large burrow systems mainly with vertical components, with an average diameter of 5 cm, and more than 1.5 m long, branching points not observed, precluding ichnoespecific identification and therefore referred to Thalassinoides isp. The fill of these burrows is mainly composed by sand and abundant fragments of "Turritella" patagonica. Most of the fossils are disarticulate and the degree of fragmentation is low. In addition to semi-infaunal and epifaunal taxa such as Atrina magellanica, Neopanis quadrisulcata, Swiftopecten nodosoplicatus, Zygochlamys geminata, Zygochlamys quemadensis, Reticulochlamys proxima, Jorgechlamys centralis among others, specimens of the deep burrowing bivalve Panopea quemadensis and shallow infaunal Cucullaea alta and Lahillia patagonica were found articulated and in life position.
Interpretation FA2 represents a lower shoreface. Bioturbation was reported for mixed sediments to a depth of as much as 1 m in shallow marine environments (Smith and Nelson 2003) resulting in a homogenized, poorly-sorted layer (Fürsich and Pandey 2003). Mollusks suggest that background sedimentation rate was low. Most of the taxa found in FA2 represent either very shallow infaunal (e.g. arcoids, cardiids) or epifaunal forms (e.g. pectinids, muricids), indicating that the substrate was stable for extended periods. In general, the rocks exposed at Darwin's locality represent subtidal facies from an offshore environment near the fair weather wave base at the bottom, to a lower shoreface at the top of the section.
PALEONTOLOGY
The most conspicuous feature of the rocks described here is the rich and well preserved fauna of mollusks contained in them. This fauna lies almost entirely within FA 2 and amongst the most abundant elements are Nucula (Lamellinucula) reticularis, Neilo ornata, Cucullaea alta, Limopsis insolita, Glycymeris cuevensis, Neopanis quadrisulcata, Swiftopecten nodosoplicatus, Zygochlamys geminata, Zygochlamys quemadensis, Reticulochlamys proxima, Jorgechlamys centralis, Crassostrea? hatcheri, Pteromyrtea crucialis, Pleuromeris cruzensis, Fasciculicardia patagonica, Spissatella lyelli, Spissatella kokeni, Hedecardium? puelchum, Cardium patagonicum, Lahillia patagonica, Dosinia laeviuscula, Ameghinomya darwini, Panopea quemadensis, Valdesia dalli, Solariella dautzenbergi, Calliostoma santacruzense, "Turritella" ambulacrum, "Turritella" patagonica, Polinices santacruzensis, Glossaulax vidali, Cirsotrema rugulosum, Trophon santacruzensis, Xymenella dautzenbergi, Xymene cossmanni, Enthacanthus monoceros, Crassilabrum hatcheri, cf. Ocenebra iheringi, Urosalpinx archipatagonica, Neoimbricaria patagonica, Antimelatoma quemadensis, and Dentalium sp. All these taxa are also found in the Punta Entrada Member of the Monte León Formation, at the mouth of the Río Santa Cruz, the only difference is the relative abundance of taxa in the two areas. They are far less numerous at Puerto Deseado, but this can be due to the fact that the exposures are also very much reduced there. Darwin's specimens were described by Sowerby and four of the species described by him were based on material from "Port Desire". Three of these were bivalves and one is a gastropod. The latter is the most conspicuous fossil at both localities: Neilo ornata (Sowerby, 1846) (Fig. 4a). It is not strange that Sowerby had only "fragments" (see Griffin and Nielsen 2008), as most of the specimens of this species, although complete in the rock, are very easily broken when collected, possibly as a result of the shell structure in this group. The species is quite common in Puerto Deseado, but it also occurs in the Santa Cruz area (sensu Darwin, i.e., just upstream from the mouth of the river in the Atlantic Ocean), where it is generally found at the base of the section. Other species of Neilo are known from Paleogene and Neogene rocks exposed in different areas of Patagonia and Tierra del Fuego, but the relationships among them remain rather obscure, as in most cases the material consists of fragmentary molds.
 Figure 4: a) Neilo ornata (Sowerby, 1846). Lectotype, NHM-L 27974; b) Cucuallaea alta Sowerby, 1846, GHUNLPam 16859, external view; c) Jorgechalmys centralis (Sowerby, 1846), GHUNLPam 16860, left valve; d) "Turritella" patagonica Sowerby, 1846, GHUNLPam 16861.
Figure 4: a) Neilo ornata (Sowerby, 1846). Lectotype, NHM-L 27974; b) Cucuallaea alta Sowerby, 1846, GHUNLPam 16859, external view; c) Jorgechalmys centralis (Sowerby, 1846), GHUNLPam 16860, left valve; d) "Turritella" patagonica Sowerby, 1846, GHUNLPam 16861.
Cucullaea alta Sowerby, 1846 (Fig. 4b). Also common at Monte León, this species appears sparsely in FA2, and is less abundant than at the localities further south. Darwin's original material - i.e., Sowerby's syntypes - included three specimens, one of them a juvenile from "Santa Cruz", and the other two (possibly the two separate valves of the same specimen) are from "Port Desire". One of the latter is a large well preserved specimen, similar to those found by us at this locality. One of Darwin`s specimens from Puerto Deseado is an adult, but the two are fairly well preserved. This species also appears at other Patagonian localities in which rocks of similar age are exposed, although it is never quite as abundant as it is at the exposures at the mouth of the Río Santa Cruz and environs. Jorgechlamys centralis (Sowerby, 1846) (Fig. 4c). This large pectinid is known from rocks belonging to the Monte León Formation, or to its equivalents such as the Centinela and Chenque Formations. The type material comes from "Santa Cruz" and "Port Desire", but lectotype designation by Griffin and Nielsen (2008) has restricted the type locality to Puerto Deseado. It is an important species as it has been used in biotratigraphic zonations of Patagonian Cenozoic units (del Río 2004). "Turritella" patagonica Sowerby, 1846 (Fig. 4d). This species is the most common mollusk in the Monte León Formation. It appears in great numbers and constitutes a significant part of the deposits in which it occurs. The exact generic placement of this species remains as yet uncertain, as pointed out by Griffin and Nielsen (2008). This taxon occurs in many other localities where marine rocks of this age are exposed. It is very common in the Chenque, Centinela, and Carmen Silva formations, albeit in none of these it reaches the abundance showed in the Monte León Formation.
CONCLUSIONS
South of Puerto Deseado the rocks exposed along the cliffs show a lithological composition that is not distinguishable from those of the Monte León Formation at its type locality further south. The succession exposed in the Puerto Deseado area contains subtidal facies from an offshore environment near fair weather wave base at the bottom to a lower shoreface at the top of the section. The type specimens of Neilo ornata, Cucullaea alta, Jorgechlamys centralis and "Turritella" patagonica described by Sowerby (1846) come - in all cases - from the cliffs south of Puerto Deseado. The mollusks collected by us from FA2 are also found in the Punta Entrada Member of the Monte León Formation at the mouth of the Río Santa Cruz suggesting an early Miocene age for Darwin's Port Desire locality.
ACKNOWLEDGMENTS
Prefectura Naval Argentina (Puerto Deseado) was instrumental in carrying out field work at Darwin's locality. The help of Miguel Santillán is also acknowledged. J. Todd (The Natural History Museum, London) provided photographs of Sowerby's type material. The manuscript benefited from thorough reviews by Noelia Carmona, Sven Nielsen and Guido Pastorino. Financial support of CONICET and Universidad Nacional de La Pampa are also duly acknowledged.
WORK CITED IN THE TEXT
1. Ameghino, F. 1896. Notas sobre cuestiones de geología y paleontología argentina. Boletín del Instituto Geográfico Argentino 17: 87-119.
2. Ameghino, F. 1898. Sinopsis Geológico-Paleontológica. Segundo Censo de la República Argentina 1: 111-225.
3. Ameghino, F. 1900-1902. L'age des formations sédimentaires de Patagonie. Anales de la Sociedad Científica Argentina 50: 109-130, 145-165, 209-229 (1900); 51: 20-39, 65-91 (1901); 52: 189-197, 244-250 (1901); 54: 161-180, 220 -249, 283-342 (1902).
4. Bertels, A. 1970. Sobre el "Piso Patagoniano" y la representación de la época del Oligoceno en Patagonia austral, República Argentina. Revista de la Asociación Geológica Argentina 25: 495-501.
5. Bertels, A. 1980. Estratigrafía y foraminíferos (Protozoa) bentónicos de la Formación Monte León (Oligoceno) en su área tipo, provincia de Santa Cruz, República Argentina. 2º Congreso Argentino de Paleontología y Bioestratigrafía y 1º Congreso Latinoamericano de Paleontología (Buenos Aires, 1978), Actas 2: 213-273.
6. Bottjer, D.J., Arthur, M.A., Deanm, W.E., Hattin, D.E. and Savrda, C.E. 1986. Rhythmic bedding produced in Cretaceous pelagic carbonate envirnments: sensitive recorders of climatic cycles. Paleoceanography 1: 467-481.
7. Bromley, R.G. 1990. Trace Fossils. Unwin Hyman, 280 p., London.
8. Camacho, H.H. 1974. Bioestratigrafía de las formaciones marinas del Eoceno y Oligoceno de la Patagonia. Anales de la Academia de Ciencias Exactas, Físicas y Naturales 26: 39-57, Buenos Aires.
9. Camacho, H.H. 1979. Significados y usos de "Patagoniano", "Patagoniense", "Formación Patagonica", "Formación Patagonia" y otros términos de la estratigrafía del Terciario marino argentino. Revista de la Asociación Geológica Argentina 34: 235-242.
10. Crawford, R.S., Casadío, S., Feldmann, R.M., Griffin, M., Parras, A. and Schweitzer, C. 2008. Mass mortality of fossil decapods within the Monte León Formation (Early Miocene), Southern Argentina: Victims of Andean volcanism. Annals of the Carnegie Museum of Natural History 77: 259-287.
11. Darwin, Ch. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R. N. during the years 1832 to 1836. Smith, Elder and Co., 279 p., London.
12. del Río, C.J. 2004. Tertiary marine molluscan assemblages of eastern Patagonia (Argentina): a biostratigraphic analysis. Journal of Paleontology 78: 1097-1122.
13. del Río, C.J. and Camacho, H.H. 1998. Tertiary nuculoids and arcoids of eastern Patagonia (Argentina). Palaeontographica Abt. A 250: 47-88.
14. Di Paola, E.C. and Marchese, H.G. 1973. Litoestratigrafía de la Formación Patagonia en el área tipo (Bajo de San Julián-desembocadura del río Santa Cruz). Provincia de Santa Cruz. República Argentina. 5º Congreso Geológico Argentino, Actas 3: 207-222.
15. Feruglio, E. 1949. Descripción Geológica de La Patagonia. Ministerio de Industria y Comercio de La Nación, Dirección General de Yacimientos Petrolíferos Fiscales 2: 1-349.
16. Frenguelli, J. 1931. Observaciones estratigráficas en Bahía Sanguinetti (Santa Cruz, Patagonia). Anales de la Sociedad Científica de Santa Fe 3: 237-283.
17. Fürsich, F.T. and Pandey, D.K. 2003. Sequence stratigraphic significance of sedimentary cycles and shell concentrations in the Upper Jurassic and Lower Cretaceous of Kachchh, western India. Palaeogeography, Palaeoclimatology, Palaeoecology 193:285-309.
18. Giacosa, R.E., Césari, O. and Genini, A. 1998. Hoja Geológica 4766-II y IV Puerto Deseado. Servicio Geológico Minero Argentino, Boletín 240, 72 p., Buenos Aires.
19. Griffin, M. and Nielsen, S.N. 2008. A revision of the type specimens of Tertiary molluscs from Chile and Argentina described by d'Orbigny (1842), Soweby (1846) and Hupé (1854). Journal of Systematic Palaeontology 6: 251-316.
20. Ihering von, H. 1897. Os Molluscos dos terrenos terciarios da Patagonia. Revista del Museo Paulista 2: 217-382.
21. Ihering von, H. 1899. Die Conchylien der pataognischen Formation. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie 1899: 1-46.
22. Ihering von, H. 1907. Les mollusques fossiles du Tertiaire et du Crétacé superieur de l'Argentine. Anales del Museo Nacional de Buenos Aires, serie 3: 1-611.
23. Ihering von, H., 1914. Catalogo de molluscos Cretáceos e terciarios da collecçao do auctor. Notas do Museu Paulista 1: 1-113.
24. Legarreta, L. and Uliana, M.A. 1994. Asociaciones de fósiles y hiatos en el Supracretácico-Neógeno de Patagonia: una perspectiva estratigráfico-secuencial. Ameghiniana 31: 257-281.
25. Malumián, N. 1999. La sedimentación y el volcanismo terciarios en la Patagonia extraandina. In Caminos, R. (ed.) Geología Argentina, Anales del Instituto de Geología y Recursos Minerales 29(18): 557-612.
26. Malumián, N. 2002. El Terciario marino. Sus relaciones con el eustatismo. In Haller, M.J. (ed.) Geología y Recursos Naturales de Santa Cruz, Relatorio 15º Congreso Geológico Argentino I-15: 237-244.
27. Náñez, C. 1988. Foraminíferos y bioestratigrafía del Terciario Medio de Santa Cruz oriental. Revista de la Asociación Geológica Argentina 43: 493-517.
28. Orbigny, A. d'. 1834-1847. Voyage dans l'Amérique méridionale (le Brésil, la République orientale de l'Uruguay, la République Argentine, la Patagonie, la République du Chili, la République de Bolivia, la République du Pérou), exécuté pendant les années 1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833, 5 (3): Mollusques: i-xliii, 1-758, láms. 1-85, vol. 3, part. 3, 290 p., P. Bertrand. Paris, V. Levrault, Strasbourg,
29. Ortmann, A.E. 1902. Tertiary Invertebrates. In Scott, W.B. (ed.) Reports of the Princeton University Expedition to Patagonia 1896-1899, J. Pierpoint Morgan Publishing Foundation, volume 4, Paleontology I, Part 2: 45-332, Princeton.
30. Parras, A., Griffin, M., Feldmann, R., Casadío, S., Schweitzer, C. and Marenssi, S. 2008. Correlation of marine beds based on Sr- and Ardate determinations and faunal affinities across the Paleogene/Neogene boundary in southern Patagonia, Argentina. Journal of South American Earth Sciences 26: 204-216.
31. Smith, A.M. and Nelson, C.S. 2003. Effects of early sea-floor processes on the taphonomy of shelf skeletal carbonate deposits. Earth Science Reviews 63:1-31.
32. Sowerby, G.B. 1846. Descriptions of the Tertiary fossil shells from South America. In: Darwin, Ch. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R. N. during the years 1832 to 1836. Smith, Elder and Co., Appendix: 249-264, London.
33. Zinsmeister, W.J. 1981. Middle to late Eocene invertebrate fauna from the San Julián Formation at Punta Casamayor, Santa Cruz province, Southern Argentina. Journal of Paleontology 55: 1083-1102.
Charles Darwin and the first scientific observations on the patagonian shingle formation (Rodados Patagónicos)
Oscar A. Martínez1, Jorge Rabassa2,3, Andrea Coronato2,3
1Universidad Nacional de la Patagonia-San Juan Bosco, Sede Esquel, Esquel, Chubut. Email: oam1958@gmail.com 2CADIC, CONICET, C.C.92, 9410 Ushuaia, Tierra del Fuego. 3Universidad Nacional de la Patagonia-San Juan Bosco, Sede Ushuaia, Tierra del Fuego.
ABSTRACT
The Rodados Patagónicos is one of the most intriguing lithostratigraphic units in the Late Cenozoic of Patagonia. Charles Darwin named these gravels as the "Patagonian Shingle Formation", when he discovered them during his trip to Patagonia on board HMS Beagle in 1832. According to the prevailing paradigm of the time, he assigned these deposits to a giant transgression during the Great Universal Déluge epoch, considering that their formation was related to wave action along the beach in ancient times. The name of Rodados Patagónicos, as they are generally known in the Argentine geological literature, is usually confusing since it has been applied to a wide number of geological units of multiple origin and age. Many authors have discussed the nature and origin of these gravels, considering them to have been formed by piedmont, alluvial, colluvial, glaciofluvial, and/or marine processes. Today, it is accepted that the term Rodados Patagónicos includes gravel deposits of varied nature and age, perhaps with a prevalence of piedmont genesis in northern Patagonia and glaciofluvial dynamics in southern Patagonia and Tierra del Fuego.
Keywords: Charles Darwin; Patagonian Shingle Formation; Late Cenozoic; Patagonia; Tierra del Fuego.
RESUMEN: Charles Darwin y las primeras observaciones científicas sobre los Rodados Patagónicos. Los Rodados Patagónicos son algunas de las unidades litoestratigráficas más sorprendentes del Cenozoico tardío de Patagonia. Charles Darwin dio a estas gravas el nombre de Patagonian Shingle Formation, cuando las descubrió durante su viaje a Patagonia en el HMS Beagle en 1832. De acuerdo con los paradigmas dominantes de la época, asignó estos depósitos a una transgresión gigantesca durante el "Gran Diluvio Universal", considerando que su formación estaba relacionada a la acción del oleaje a lo largo de la playa en tiempos antiguos. El nombre de Rodados Patagónicos, como generalmente se los conoce en la literatura geológica argentina, es usualmente confuso, ya que ha sido aplicado a un amplio número de unidades geológicas, de múltiple origen y edad. Muchos autores han discutido la naturaleza y génesis de estas gravas, considerándolas como formadas por procesos diversos, ya sea pedemontanos, aluviales, coluviales, glaciofluviales, y/o marinos. En la actualidad, se acepta que el término Rodados Patagónicos incluye a depósitos de grava de naturaleza y edad muy variadas, quizás con una predominancia de aquellos de génesis pedemontana en Patagonia septentrional y debidos a la dinámica glaciofluvial en Patagonia austral y Tierra del Fuego.
Palabras clave: Charles Darwin; Rodados Patagónicos; Cenozoico tardío; Patagonia; Tierra del Fuego.
INTRODUCTION
The outstanding work of Charles Darwin in the biological sciences has concealed his significant contributions to geology and other earth sciences. Perhaps because of this reason, the great influence of his findings in South American earth sciences is seldom appraised in the literature beyond his biological theories. The publication of The Origin of Species (Darwin 1859) was the onset of a period in which there were so many radical changes in the structure of western knowledge that it can be considered an authentic scientific revolution. However, the strengthening of this new paradigm on the origin and evolution of living beings was accompanied, and mostly complemented, by the formulation of new approaches to the great geological dilemmas of the times. By then, the paramount work of Charles Lyell (1830-1833) represented the gradualist principles within the geological sciences, which appeared as a reaction and antipode position against the catastrophist theories, that postulated that natural history was, essentially, a succession of universal cataclysm that had dramatically modeled the surface of the Earth, generating mass extinctions and the rise of new species different to the previously existing ones. Darwin's work, basically of a gradualist nature, fired catastrophism the final blow. Almost at the same time that Darwin was traveling on board HMS Beagle, the last steps towards the presentation of the Glacial Theory were being fulfilled in central Europe (Louis Agassiz, in 1837; Agassiz 1840, in Imbrie and Imbrie 1979), which would deeply modify the ideas about the origin and evolution of the landscape in the northern hemisphere and, as it happened later on, on the understanding of the global climate system. This new theory did not adjust to Bible principles that underpinned the Great Universal Déluge as the main cause of most of the present landscape features, and strongly supported by the aforementioned catastrophist conception. The first volume of Lyell's Principles of Geology was published in 1830, only one year before Darwin set out on his 5-year voyage to the Southern Hemisphere. This volume, and the second one that he received when HMS Beagle was in Buenos Aires in 1832, became the conceptual platform from which Darwin made his observations and formulated his principal hypotheses on the geological sciences in general and of South American geology in particular. It is frequently believed that Darwin's main contributions to earth sciences are his works on plutonic and metamorphic rocks and his ideas on the origin of the volcanic islands and reef barriers. In this article, we want to emphasize the thoughts dedicated by this great scientist to one of the more interesting and intriguing geological units, not only of those days but even today, as are the so called Rodados Patagónicos. For a review of Darwin's work as a Quaternary geologist and as a glaciologist see Rabassa (1995). The discussion of Darwin's process of identification, description and interpretation of the Rodados Patagónicos, which are ubiquitous over most of the surface of Argentine Patagonia (Fig. 1), reveals once again his scientific talent and pioneer activity in the area and also allows recognition of relevant aspects of the historical-scientific background in which such process occurred.
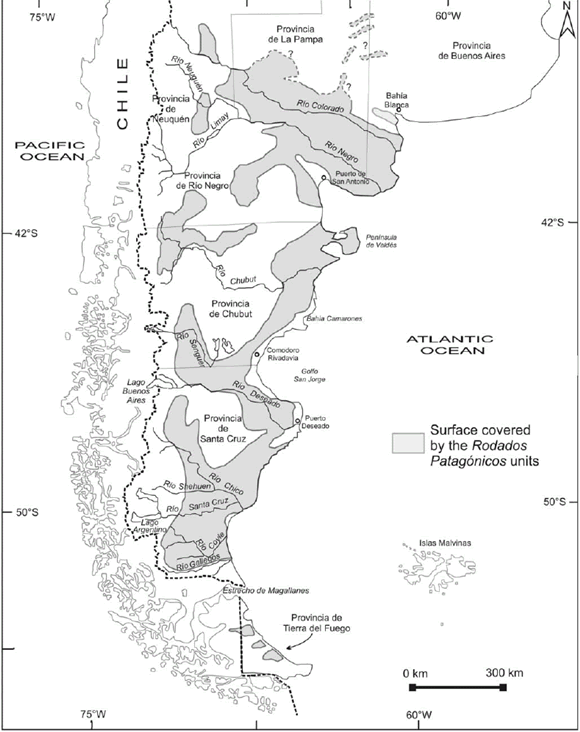 Figure 1: Location map of Patagonia and Tierra del Fuego showing the areas covered by the Rodados Patagónicos (modified from Clapperton 1993).
Figure 1: Location map of Patagonia and Tierra del Fuego showing the areas covered by the Rodados Patagónicos (modified from Clapperton 1993).
CHARLES DARWIN AND THE "DISCOVERY" OF THE RODADOS PATAGÓNICOS
The voyage of the HMS Beagle took place between 1831 and 1836. The first opportunity in which Charles Darwin identified gravel deposits that are today known as Rodados Patagónicos was in 1833, during his expedition to the surroundings of the present city of Bahía Blanca, southern Buenos Aires province (Fig. 1). There, he observed a layer that was less than a meter thick, composed of small pebbles, essentially porphyritic rocks, that were lying on top of the "Pampean beds" and that were the base over which the frequent large dunes in the area are deposited (Darwin 1846). Starting here, and later during different landings as they sailed southwards, such as San Antonio, the mouth of Río Chubut, Puerto Deseado, San Jorge Gulf and the mouth of Río Santa Cruz (Fig. 1), Darwin described the outcrops at the scarps of tablelands and terraces stretching along the sea. At the same time, he began working on the hypothesis that these gravels were the product of alluvial accumulation at the foot of the Andean Cordillera and later spread out by wave action during a marine transgression. He verified the vast continuity of these gravel beds he named as the "Gravel Formation" or "Patagonian Shingle Formation", concluding that they represented one of the main physical features of this region. The term "shingle" referred to the gravels which are the result of wave action on the cliffs along many sectors of the British coasts (Fig. 2).
 Figure 2: Beach gravel deposits (shingle beach) in a classical locality along the English coast, representing the kind of deposits that Darwin had seen before starting his voyage on board HMS Beagle. Darwin's home memories about these kind of deposits lead him to interpret the gravel deposits that he found everywhere along the Patagonian coasts and rivers as"shingle formations", thus giving support to the original, historical name of these units. Reproduced with permission of www.beenthere-donethat.org.uk, copyright © by Barry Samuels.
Figure 2: Beach gravel deposits (shingle beach) in a classical locality along the English coast, representing the kind of deposits that Darwin had seen before starting his voyage on board HMS Beagle. Darwin's home memories about these kind of deposits lead him to interpret the gravel deposits that he found everywhere along the Patagonian coasts and rivers as"shingle formations", thus giving support to the original, historical name of these units. Reproduced with permission of www.beenthere-donethat.org.uk, copyright © by Barry Samuels.
To Darwin's eyes, the vast expanse of the Patagonian gravel beds was awesome and astounding, in comparison to what he had seen in Europe before, to a point that he considered that these units were the largest ones of this kind in the entire world. He assumed that a clear evidence of the marine origin (in fact, submarine for him) of these strata was the frequent finding of Recent marine shells scattered on top and even within these terraces. Although several authors later discarded this genetic interpretation, it was Feruglio (1950) who confirmed that the shells had been accumulated by human action and they were actually archaeological sites. There are other elements that contributed to Darwin's choice of his marine (submarine) process interpretation. Firstly, the great widening of the Río Santa Cruz valley nearby its sources at Lago Argentino (Fig. 1), more than 300 km from the Atlantic coast, was wrongly interpreted as an ancient estuary. This large landform is today known to have been generated due to recurrent Pleistocene glaciation and the action of glaciofluvial streams (Mercer 1976, Clapperton 1993, Schellmann 1998, among others). It is also accepted today that the building up of the extensive, step-like terraces and/or tablelands of the region is also due to the same glaciofluvial processes. However, Darwin linked these landforms to the impact of Atlantic Ocean transgressions that reached locations very close to the Andean Cordillera. Besides, he also considered that it was very likely that Patagonia would have been crossed by many sea passages in the past, similar to the present Magellan Straits (Fig. 1), which connected both oceans. It should be considered that almost simultaneously with Darwin's pioneer scientific observations in Patagonia (1833-1834), the ideas that led Louis Agassiz to postulate his Glacial Theory in 1837, were growing steadily. For an ample discussion of this epistemological process, see Imbrie and Imbrie (1979). Concerning Darwin's geological background, it was probably not conceivable that glaciers would have had in the recent geological past a larger extent that today. Even less conceivable was that areas which are ice free today and very far from the glacier boundaries could have been covered by large ice masses in the past; a feature that has been shown was a distinctive characteristic of Patagonia (Caldenius 1932, Feruglio 1950, Clapperton 1993, Coronato et al. 2004, Rabassa 2008). Darwin focused his geological analysis accepting Lyell's statements as a foundation, albeit in a critical manner, as Lyell was still supporting the hypothesis of a large flooding -a phenomenon of Biblical roots- which had had a key role in the origin of many features of the Earth's surface. It is interesting to consider that in those years Lyell believed that the erratic boulders, today accepted as essentially of strict glacial origin, had been transported through usually very long distances by icebergs generated by such flooding, to be later abandoned on land as the sea withdrew. This interpretation, which Lyell abandoned a few years later, was very influential on Darwin's intellectual work. The large boulders of foreign rocks (of Andean origin) that are lying on or partially buried in the gravel beds along extra-Andean areas of southernmost Patagonia (Darwin 1848), were thus of marine origin for Darwin, becoming so another strong line of reasoning in favor of a similar or identical origin for the Shingle Formation.
THE SEDIMENTARY MATERIALS THAT HAVE BEEN NAMED AS RODADOS PATAGÓNICOS
Previous works
After Darwin's early contributions there were several authors that documented the existence of these characteristic gravel and sand beds in Patagonia (Table 1). Doering (1882) named them as Piso Tehuelche and in a pioneer manner interpreted them as of glaciofluvial origin in a moment in which the Glacial Theory was well accepted by the scientific community. This author correlated them with the lower section of the Pampean sediments, based on the occurrence of calcareous duricrusts locally known as tosca, and assigned them an Early Pliocene age. Carlos Ameghino (1890) was the first geologist to discard a single origin for these materials and he differentiated between the marine deposits forming the high terraces and the low terrace sedimentary beds, referring the first ones as the Formación Araucanense, deposited in successive epochs since the Early Miocene. Mercerat (1893) studied these accumulations in the southernmost part of Patagonia between the Río Santa Cruz and the Magellan Straits. He named them as Rodados Tehuelches and assigned them a marine origin and a pre-Pliocene age. Hatcher (1897) also considered them of marine origin and attributed them to a sea transgression that would have covered all of Extra-Andean Patagonia during the Pliocene, reinstating the Darwinian name of Shingle Formation. Nordenskjöld (1897), who was strongly influenced by the recently introduced Glacial Theory and his wide knowledge of the glacial landscapes in Scandinavia and northern Europe, correctly proposed a glaciofluvial origin for the gravel deposits in southern Santa Cruz province and the Magellan Straits, but he did not discuss the origin of similar units farther north. Florentino Ameghino (1906) returned to the topic from a regional perspective, insisting that it was not appropriate to assign a unique origin to all gravel deposits and that they could have a different genesis according to their location.
TABLE 1: Summary table of the main contributions to the knowledge on the Rodados Patagónicos, since the first descriptions (Darwin 1848) up to the end of the 20th century. The great historical controversy -which still persists- may be identified, concerning the genesis and age of these units.
The first author to relate the Rodados Patagónicos to the development of the glacial periods in the Patagonian Andes was Rovereto (1912), who recognized a link to four hypothetical glaciations according to the Alpine scheme then in use. According to him these glaciations were related to different marine terraces with a mollusk fauna quite similar to the present one, as suggested by his studies along the Atlantic coast. Keidel (1917) disagreed with the hypotheses of the previous workers, postulating that the gravels that cover much of the tablelands and terraces of northwestern Patagonia represented alluvial bajadas built by fluvial streams coming from the Andes, during the Pliocene and the Quaternary, in response to regional uplift events. Keidel was the first to note the unconformity between the gravels and the underlying Late Tertiary marine and continental sedimentary rocks. Later, Bonarelli and Nágera (1922) returned to the ideas about the marine origin of the gravels and assumed that the so-called Rodados Tehuelches of the highest terraces were at least of Pliocene age, which had been dispersed later by the action of marine waters pertaining to a transgression that reached the foothill of the Andes. These later were the source of the fluvial deposits of the lower terraces, carved after successive episodes of river base drop. Windhausen (1931) suggested that the higher beds were deposited in an alluvial manner over a rather flat relief with a very gentle slope, whereas the topographically lower, terraced gravels were the consequence of glaciofluvial deposition in different stages of uplift that occurred during the Quaternary. Based on the ideas of Rovereto (1912), Frenguelli (1931) distinguished the Tehuelchiano beds, composed of three orders of marine terraces and other continental ones corresponding to the Post-Tehuelchiano, formed by low terrace gravels, of postglacial age. Caldenius (1932, 1940) assigned a fluvial and glaciofluvial origin to the Rodados Tehuelches, originally deposited in the shape of piedmont glaciofluvial cones and he suggested that these units had undergone certain amount of reworking due to solifluction processes. Likewise, he recognized the existence of higher level gravel beds and of an older age than even the oldest glaciations, which he named as Initioglacial. Groeber (1936) proposed a mixed alluvial and colluvial origin for these gravels. Feruglio (1950) recognized the existing relation among the fluvial terraces of the different fluvial systems of the southernmost Patagonian meseta, in the valleys of the Chubut, Deseado, Shehuen, Coyle, Santa Cruz and Gallegos rivers (Fig. 1). The great dimensions of the terraces, the thickness of their alluvial mantles and the marked relief that separated them justified his interpretation linked to the glacial and interglacial periods that affected the mountain ice sheet of the Patagonian Andes since the Pliocene, and to a lesser extent, to phases of tectonic uplift. On these terraces Feruglio (1950) identified moraine deposits and glaciofluvial gravels of varied lithology, but mostly of eruptive rocks. Frenguelli (1957) agreed in general terms with Feruglio's (1950) interpretations. The first really rigorous systematic and solid studies on the gravels were done by Fidalgo and Riggi (1965, 1970), who based their interpretations upon geomorphological and sedimentological observations in the surroundings of Lago Buenos Aires (Santa Cruz province; Fig. 1). In agreement with Caldenius (1932), they classified these materials into two large groups: (a) those of fluvial and piedmont origin (Rodados Patagónicos, sensu stricto), located at higher altitude and covering the tablelands and pediments, and (b) those that form the glaciofluvial plains that are found within the valleys or depressions around the mesetas and therefore of younger age. According to Fidalgo and Riggi (1965, 1970), all other deposits of more restricted extent as those building up the flanking pediments should also be considered as Rodados Patagónicos, a proposal that Clapperton (1993) considered as of little value. The development of absolute dating and the consequent confirmation of the occurrence of glaciations older than the Pleistocene in Santa Cruz province allowed Mercer (1976) to identify accumulations of glaciofluvial origin, referring them to the Rodados Patagónicos, with an age equivalent or even older than that of those of piedmont origin that had been mentioned as the oldest by some authors. González Díaz and Malagnino (1984) and Malagnino (1989) centered their observations in northern Patagonia and they concurred in assigning a polygenetic character to the Rodados Patagónicos at these latitudes, proposing an essentially glaciofluvial origin for the younger ones, and broadly a piedmont genesis, possibly associated to tectonic pulses for the older ones. Clapperton (1993) and later Lapido and Pereyra (1999), reviving the essentials of Ameghino's (1906) hypothesis, proposed classifying the deposits in (a) those located in northern Patagonia, between the Negro and Colorado rivers (Fig. 1), to which they assigned a dominantly piedmont origin and (b) the gravels of southern Patagonia, in the provinces of Chubut and Santa Cruz, which were interpreted as of predominantly glaciofluvial nature. During the second half of the 20th century the geological surveys of Extra-Andean Patagonia became more frequent and many authors have proposed a series of lithostratigraphic units corresponding to the Rodados Patagónicos. Among many others should be mentioned the contributions of Volkheimer (1963, 1964, 1965 a and b, 1973), Cortelezzi et al. (1965, 1968), González (1971, 1978), Coira (1979), Fidalgo and Rabassa (1984), Page (1987), Cortés (1987), González Díaz (1993a, b and c), Panza (1994a, 1994b), Panza and Irigoyen (1994) and more recently, Strelin et al. (1999), Caminos (2001), González Díaz and Tejedo (2002), Pereyra et al. (2002) and Leanza and Hugo (1997, 2005). Meglioli (1992) mapped as Patagonian Gravels -without distinguishing about their genesis- the plains located along the southern margin of the Río Gallegos, the Río Chico de Santa Cruz basin and several basins in Tierra del Fuego Island (Fig. 1). The slender relief of these gravelly plains, undifferentiated from a genetic point of view, is interrupted by the Quaternary volcanic cones that form the Pali-Aike volcanic field. The glaciofluvial gravels from the Pleistocene glacial advances are distributed according to the moraine morphology, either in frontal or marginal position. Although Meglioli (1992) did not present details of the location of each one of the glaciofluvial terraces, he defined their spatial setting and assigned them to the Cabo Vírgenes, Punta Delgada, Primera Angostura and Segunda Angostura glaciations, or the Post-GGP I, II and III glaciations and Last Glacial Maximum, according to Coronato et al. (2004) in the Magellan Straits, Skyring and Otway sounds ice lobes (Fig. 1). In high topographic positions, Meglioli (1992) identified a thin gravel bed that is part of the Sierra de los Frailes Drift, corresponding to the Great Patagonian Glaciation (GPG, according to Coronato et al. 2004), whose age was established in ca. 1 Ma (Ton That et al. 1999, Rabassa 2008). Meglioli (1992) defined several units of rounded and subrounded gravels of similar origin in Tierra del Fuego and named them as Rodados Fueguinos, thus recognizing that this type of unit is also present in the southernmost end of the continent. Finally, the work of Panza (2002) provided an integrated view of the Cenozoic gravels within the province of Santa Cruz, whereas Martínez and Coronato (2008) extended this analysis to the rest of Patagonia.
CHARACTERIZATION OF THE RODADOS PATAGÓNICOS
The Rodados Patagónicos are accumulations of gravelly clasts (Figs. 3 and 4), cemented or not, substantially rounded, with pebbles and cobbles as the dominant size fractions, in a sandy or silty/clayish matrix, of highly variable lithology, although with a certain predominance of basic and mesosilicic volcanics and acid plutonic rocks. They range between the Andean Cordillera and the Atlantic Ocean coast, and from the northern flank of the Río Colorado valley to the island of Tierra del Fuego (Fig. 1). They tend to form horizontal to subhorizontal mantles of varied extension and thickness, which are located in different topographical positions, usually showing an east-west dominant gradient, and the genesis of which may be variable according to the considered unit or geographical area. They were generated at some time during the Late Cenozoic. They may be forming different landforms or their relicts, such as inactive flood plains, alluvial terraces, alluvial fans, bajadas, pediment covers, proglacial plains and structural plains covers (Fig. 5). Hence, the great diversity of the many variables that play a part in the definition of these units (Table 2), i.e. (a) sedimentological / petrological (composition, grain size, shape, selection, among other parameters), (b) spatial (shape, elevation, slope, size, extent, thickness of the beds), (c) chronological (tentatively between the Late Miocene and the Holocene) and (d) genetic (fluvial, piedmont, glaciofluvial, periglacial, among other possible environments).
 Figure 3: A Rodados Patagónicos outcrop in the area of Lago Argentino, Río Santa Cruz valley, Province of Santa Cruz. Photograph by A. Coronato, 2006.
Figure 3: A Rodados Patagónicos outcrop in the area of Lago Argentino, Río Santa Cruz valley, Province of Santa Cruz. Photograph by A. Coronato, 2006.
 Figure 4: Outcrop of the Rodados Patagónicos in the tablelands of Central Chubut Province. Photograph by O.A. Martínez, 2007.
Figure 4: Outcrop of the Rodados Patagónicos in the tablelands of Central Chubut Province. Photograph by O.A. Martínez, 2007.
 Figure 5: Structural terraces and other landforms covered by the Rodados Patagónicos in southeastern Chubut Province, mapped on a Landsat satellite image (from Martínez and Coronato 2008).
Figure 5: Structural terraces and other landforms covered by the Rodados Patagónicos in southeastern Chubut Province, mapped on a Landsat satellite image (from Martínez and Coronato 2008).
TABLE 2: Genetic classification and general sedimentological characterization of the Rodados Patagónicos (modified from Martínez and Coronato 2008).
It is clear then that the concept of Rodados Patagónicos is ample enough, and thus ambiguous, so as to hamper its use in a regional stratigraphic sense. However, it may have a useful practical application as a generic term in those cases - not infrequent-, in which it would be impossible or unnecessary to establish the age and/or genesis of these gravel layers. As suggested by Lapido and Pereyra (1999) the lack of chronostratigraphic studies and of absolute datings in the different Quaternary units of the region renders any predetermined time framework and/or geographical location pattern of these deposits only tentative and incomplete. When the gravel mantles are grouped more or less in a parallel manner with respect to the present drainage networks, they might be genetically related to fluvial valley processes. This possible genesis should be considered as the result of both climatic fluctuations (glacial and interglacial periods) and base level modifications in response to Late Cenozoic tectonic and epeirogenic uplift (Strelin et al. 1999). Besides, it seems relevant to consider that major piedmont aggradation events should have followed and, in some cases, even coincided at the regional level with those of glaciofluvial nature, at least since the late Miocene (Martínez and Coronato 2008). The general idea of advocating an older age for the piedmont deposits in relation to those formed by glaciofluvial action (Fidalgo and Riggi 1965, 1970) seems inconvenient at least, considering the complexity in the tectonic and climatic evolution of such extensive a region as Patagonia (Lapido and Pereyra 1999). J.L. Panza (pers. comm., while acting as a reviewer of an earlier version of this manuscript) did not agree with some of our conclusions. He considered that most of, if not all, those deposits assigned to the Rodados Patagónicos of ages older than 1.2-1.0 Ma in the Province of Santa Cruz are not related to glaciofluvial proceses or genetically or timely associated to the major Patagonian glaciations, being much older than these. He understands that there is no synchronism between the main aggradational events and those of glaciofluvial nature, particularly in Northern Patagonia. He also considers inappropriate our discussion of the relative ages of piedmont and glaciofluvial deposits. J.L. Panza's comments are very valuable and worthy. However, we would like to state that we have never denied the fluvial/ aggradational/piedmont origin for some of the Rodados Patagónicos units. Moreover, we have clearly maintained (see for instance Tables 1 and 2) that this genesis is one of the possible major sources for these units. Our intention has been just to make noticeable that some of the accumulations of Rodados Patagónicos, and particularly those of Early Pleistocene and older ages (Rabassa et al. 2005), may have been generated by glaciofluvial action during very ancient glaciations, older than the Great Patagonian Glaciation, even though these glacial events were growing small, isolated ice caps before the Patagonian Mountain Ice Sheet finally developed around ca. 1.2 Ma (Rabassa 2008). Though on-going and future research will undoubtedly elucidate this puzzle, the scale and complexity of this problem has kept this discussion open for over a century and obviously it will probably remain so for a long time.
FINAL REMARKS
This article intends to give renewed importance to the historic role that the work of Charles Darwin on the Rodados Patagónicos had at his time, precisely in a profoundly revolutionary moment within the earth sciences, when new ideas were thriving and new paradigms were precipitously put forward. The Darwinian production concerning the Rodados Patagónicos compels us to recognize the enormous merits of this author as an intuitive geologist of great intellectual audacity and who conceived science, as many other naturalists of those times, as an essentially integral and multidisciplinary activity. Thus, Darwin achieved a prominent position in this discipline in Argentina, perhaps unintentionally, since his most insightful interests were in the fields of biology and anthropology. Nevertheless he is widely recognized in the earth sciences particularly as a petrologist (some of the first descriptions of plutonic and metamorphic rocks), sedimentologist (pioneer reconnaissance of old sedimentary rocks and modern sediments), geomorphologist (identification and characterization of terraces, tablelands, dunes, estuaries, moraines, erratic boulders, etc.), stratigrapher (a visionary definition of the Pampean units), paleontologist (transcendental discoveries of relevant localities for Tertiary and Pleistocene fossil mammals in the Pampean region) and glaciologist (innovative observations of the Patagonian and Fuegian glaciers). Darwin was one of the most important geologists and geomorphologists of the 19th century, very far ahead of his time, and his forerunner ideas needed over a century to be revised, incorporated, confirmed, or dismissed. Even today we continue revisiting his ideas and still work pursuing the search of valuable, ground-breaking concepts which may still be hidden within his unforgettable writings.
ACKNOWLEDGEMENTS
The authors would like to thank Beatriz Aguirre-Urreta for her kind invitation to contribute to this volume, as well as for her permanent help and collaboration during the editorial process of the manuscript. This paper has been largely improved by important observations and criticism to earlier versions of this manuscript by Raúl De Barrio, José Luis Panza and an anonymous reviewer. We are greatly indebted and deeply grateful to all of them for their thorough, meticulous and comprehensive comments. Needless to say, inaccuracies and mistakes that may still be present in this contribution are solely the authors' responsibility.
WORKS CITED IN THE TEXT
1. Agassiz, L. 1840. Études sur les glaciers. Jent et Gassmann Libraires, 346 p., Neuchâtel.
2. Ameghino, C. 1890. Los plagiaulacídeos argentinos y sus relaciones zoológicas, geológicas y geográficas. Boletín del Instituto Geográfico Argentino 11:143-201.
3. Ameghino, F. 1906. Les formations sedimentaires du Crétacé supérieur et du Tertiarie de Patagonie. Anales del Museo Nacional Buenos Aires 15: 45-76.
4. Bonarelli, G. and Nágera, J. 1922. Observaciones geológicas en las inmediaciones del Lago San Martín (Territorio de Santa Cruz). Dirección General de Minas, Boletín 27B, 39 p., Buenos Aires.
5. Caldenius, C. 1932. Las glaciaciones cuaternarias en la Patagonia y Tierra del Fuego. Dirección General de Minas y Geología, Publicación 95, 152 p., Buenos Aires.
6. Caldenius, C. 1940. The Tehuelche or Patagonian shingle-formation. A contribution to the study of its origin. Geografiska Annaler 22: 160-181.
7. Caminos, R. 2001. Hoja Geológica 4166-I, Valcheta, Provincia de Río Negro. Servicio Geológico Minero Argentino, Boletín 310, 73 p., Buenos Aires.
8. Clapperton, C. 1993. Quaternary Geology and Geomorphology of South America. Elsevier Science Publishers B.V., 779 p., Amsterdam.
9. Coira, B. 1979. Descripción Geológica de la Hoja 40d, Ingeniero Jacobacci, Provincia de Río Negro. Servicio Geológico Nacional, Boletín 168, 94 p., Buenos Aires.
10. Coronato, A., Martínez, O. and Rabassa, J. 2004. Pleistocene Glaciations in Argentine Patagonia, South America. In Ehlers, J. and Gibbard, P. (eds.) Quaternary Glaciations - Extent and Chronology. Elsevier, Quaternary Book Series, Part 3, 49-67, Amsterdam.
11. Cortés, J.M. 1987. Descripción geológica de la Hoja 42h, Puerto Lobos, Provincia del Chubut. Servicio Geológico Nacional, Boletín 202, 68 p., Buenos Aires.
12. Cortelezzi, C., De Francesco, F. and De Salvo, O. 1968. Estudio de las gravas Tehuelches de la región comprendida entre el Río Colorado y el Río Negro, desde la costa atlántica hasta la cordillera. 3º Jornadas Geológicas Argentinas, Actas 3: 123-145.
13. Cortelezzi, C., De Salvo, O. and De Francesco, F. 1965. Estudio de las gravas Tehuelches de la región comprendida entre el Río Colorado y el Río Negro, desde la costa de la Provincia de Buenos Aires hasta Choele-Choele. Acta Geológica Lilloana 6: 65-86.
14. Darwin, C. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 280 p., London.
15. Darwin. C. 1848. On the distribution of the erratic boulder and on the contemporaneous unstratified deposits of South America. Transactions Geological Society London 6 (1842): 415-431.
16. Darwin, C. 1859. The origin of species by means of natural selection, or the preservation of races in the struggle for life. John Murray, 490 p., London.
17. Doering, A. 1882. Informe oficial de la Comisión Científica agregada al Estado Mayor General de la expedición al Río Negro (Patagonia). Entrega III, Geología: 295-530, Buenos Aires.
18. Feruglio, E. 1950. Descripción Geológica de la Patagonia. Yacimientos Petrolíferos Fiscales (YPF). 3, Ediciones Coni, 431 p., Buenos Aires.
19. Fidalgo, F. and Riggi, J.C. 1965. Los Rodados Patagónicos en la Meseta de Guenguel y alrededores. Revista de la Asociación Geológica Argentina 20: 273-325.
20. Fidalgo, F. and Riggi, J.C. 1970. Consideraciones geomórficas y sedimentológicas sobre los Rodados Patagónicos. Revista de la Asociación Geológica Argentina 25: 430-443.
21. Fidalgo, F. and Rabassa, J. 1984. Los depósitos cuaternarios. In Ramos, V.A. (ed.) Geología y Recursos Naturales de la provincia de Río Negro, 9º Congreso Geológico Argentino, Relatorio: 301-316, Buenos Aires.
22. Frenguelli, J. 1931. Nomenclatura estratigráfica patagónica. Anales de la Sociedad Científica 3, 115 p., Santa Fe.
23. Frenguelli, J. 1957. Geografía de la República Argentina. Neozoico. 2(3), 218 p., Buenos Aires.
24. González, R. 1971. Descripción Geológica de la Hoja 49c, Sierra de San Bernardo, Provincia de Chubut. Dirección Nacional de Geología y Minería, Boletín 112, 103 p., Buenos Aires.
25. González, R. 1978. Descripción Geológica de las Hojas 49a, Lago Blanco y 49b, Paso Río Mayo, Provincia de Chubut. Servicio Geológico Nacional, Boletín 154-155, 68 p., Buenos Aires.
26. González Díaz, E.F. 1993a. Mapa geomorfológico del sector de Cushamen (NO de Chubut): reinterpretación genética y secuencial de sus principales geoformas. 12º Congreso Geológico Argentino (Mendoza), Actas 6: 56-65.
27. González Díaz, E.F. 1993b. Propuesta evolutiva geomórfica para el sector de Cushamen (NO de Chubut) durante el lapso Terciario Superior-Cuaternario. 12º Congreso Geológico Argentino, (Mendoza), Actas 6: 66-72. .
28. González Díaz, E.F. 1993c. Nuevas determinaciones y mayores precisiones en las localizaciones de los términos glaciarios del "Inicio" y "Daniglacial" en el sector de Cushamen (Chubut), Noroeste del Chubut. 12º Congreso Geológico Argentino (Mendoza), Actas 6: 48-55.
29. González Díaz, E.F. and Ferrer, J. 1986. Geomorfología de la Provincia de Neuquén. C.F.I. - Expediente 181 (unpublished report), 11 p., Buenos Aires.
30. González Díaz, E.F. and Malagnino, E.C. 1984. Geomorfología de la Provincia de Río Negro. 9º Congreso Geológico Argentino, Relatorio, 154 p., Bariloche.
31. González Díaz, E.F. and Tejedo, A. 2002. Mapa geomorfológico de la Hoja 4569-IV, Escalante, Provincia de Chubut. 15º Congreso Geológico Argentino (El Calafate), Actas 2: 667-671.
32. Groeber, P. 1936. Oscilaciones del clima en la Argentina desde el Plioceno. Holmbergia, Revista Centro Estudiantes Ciencias Naturales 1: 71-84, Buenos Aires.
33. Hatcher, J.B. 1897. On the geology of southern Patagonia. American Journal of Science 4(23): 327-354.
34. Imbrie, J. and Imbrie, K.P. 1979. Ice Ages, solving the mistery. The Macmillan Press, 224 p., London.
35. Keidel, J. 1917. Ueber das patagonische Tafelland, das patagonische Gëroll und ihre Beziehungenzu den geologischen Erscheinungen im argentinischen Andengebiet und Litoral. Zeitschrift Deutsche Wissenschaftliche Verein 3(5-6): 219-245.
36. Lapido, O.R. and Pereyra, F. 1999. Cuaternario de la Patagonia Argentina. In Caminos, R. (ed.) Geología Argentina, Instituto de Geología y Recursos Minerales, Anales 23(7): 704-709, Buenos Aires.
37. Leanza, H.A. and Hugo, C. 1997. Hoja Geológica 3969-III. Picún Leufú. Provincias de Neuquén y Río Negro. Servicio Geológico Minero Argentino, Boletín 218, 135 p., Buenos Aires.
38. Leanza, H.A. and Hugo, C. 2005. Hoja Geológica 3969-I, Zapala, Provincia de Neuquén. Instituto de Geología y Recursos Minerales, Servicio Geológico Minero Argentino, Boletín 275, 128 p., Buenos Aires.
39. Lyell, C. 1830-33. Principles of Geology. John Murray, Albemarle-Street, (1830) 1, 511 p.; (1832) 2, 330 p.; (1833) 3, 109 p., London.
40. Malagnino, E.C. 1989. Paleoformas de origen eólico y sus relaciones con modelos de inundación de la Provincia de Buenos Aires. 4º Simposio Latinoamericano de Percepción Remota and 9º Reunión Plenaria Selper (Bariloche), Actas 2: 611-620.
41. Martínez, O.A. and Coronato, A. 2008. The fluvial deposits of Argentine Patagonia. In Rabassa, J. (ed.) The Late Cenozoic of Patagonia and Tierra del Fuego. Elsevier, Developments in Quaternary Sciences 11: 205-226, Amsterdam.
42. Meglioli, A. 1992. Glacial Geology of Southernmost Patagonia, the Strait of Magellan and Northern Tierra del Fuego. PhD Dissertation, Lehigh University, Department of Geological Sciences, (unpublished), 216 p., Bethlehem.
43. Mercer, H. 1976. Glacial history of southernmost South America. Quaternary Research 6: 125-166.
44. Mercerat, A. 1893. Contribuciones a la geología de la Patagonia. Anales de la Sociedad de Ciencias Argentinas 36: 65-103, Buenos Aires.
45. Moreno, F.P. 1976. Viaje a la Patagonia Austral. Editorial El Elefante Blanco - Imprenta de La Nación, 480 p., Buenos Aires.
46. Nordenskjöld, O. 1897. Algunos datos sobre la naturaleza de la región Magallánica. Anales de la Sociedad Científica Argentina 44: 190-240.
47. Page, R. 1987. Descripción Geológica de la Hoja 43g, Bajo de la Tierra Colorada, Provincia de Chubut. Dirección Nacional de Minería y Geología, Boletín 200, 85 p., Buenos Aires.
48. Panza, J.L. 1994a. Hoja Geológica 4969 - II, Tres Cerros, Provincia de Santa Cruz. Dirección Nacional del Servicio Geológico, Boletín 213, 103 p., Buenos Aires.
49. Panza, J.L. 1994b. Hoja Geológica 4966 - I y II, Bahía Laura, Provincia de Santa Cruz. Dirección Nacional del Servicio Geológico, Boletín 214, 83 p., Buenos Aires.
50. Panza, J.L. 2002. La cubierta detrítica del Cenozoico superior. In Haller, M.J. (ed.) Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino, Relatorio: 259-284, El Calafate.
51. Panza, J.L. and Irigoyen, M.V. 1994. Hoja Geológica 4969-IV, Puerto San Julián, Provincia de Santa Cruz. Dirección Nacional del Servicio Geológico, Boletín 211, 77 p., Buenos Aires.
52. Pereyra, F., Fauqué, L. and González Díaz, E.F. 2002. Geomorfología. In Haller, M.J. (ed.) Geología y Recursos Naturales de Santa Cruz. 15º Congreso Geológico Argentino, Relatorio: 325-352, El Calafate.
53. Rabassa, J. 1995. Charles Darwin, el primer geólogo y glaciólogo de Tierra del Fuego. Ciencia Hoy 6 (31): 24-36.
54. Rabassa, J. 2008. Late Cenozoic glaciations in Patagonia and Tierra del Fuego. In Rabassa, J. (ed.) The Late Cenozoic in Patagonia and Tierra del Fuego, Elsevier, Developments in Quaternary Sciences 11: 151-204, Amsterdam.
55. Rabassa, J., Coronato, A. and Salemme, M. 2005. Chronology of the Late Cenozoic Patagonian Glaciations and their correlation with biostratigraphic units of the Pampean Region (Argentina). Journal of South American Earth Sciences 20: 81-103.
56. Rovereto, G. 1912. Studi di geomorfologia Argentina. III. La valle del Río Negro: 2. Il lago Nahuel Huapi. Societta Geologica Italiana Bolletino 31: 181-237.
57. Schellmann, G. 1988. Jungkänozoische Landschaftsgeschichte Patagoniens (Argentinien). Andine Vorlandvergletscherungen, Talentwicklung und marine Terrasen. Essener Geographische Arbeiten 29, 218 p., Essen.
58. Strelin, J.A., Re, G., Keller, R. and Malagnino, E. 1999. New evidence concerning the Plio-Pleistocene landscape evolution of southern Santa Cruz region. Journal of South American Earth Sciences 12: 333-341.
59. Ton-That,T., Singer, B., Mörner, N. and Rabassa, J. 1999. Datación de lavas basálticas por 40Ar/ 39Ar y geología glacial de la región del Lago Buenos Aires. Revista de la Asociación Geológica Argentina 54: 333-352.
60. Volkheimer, W. 1963. El Cuartario Pedemontano en el noroeste de Chubut (zona Cushamen). 2° Jornadas Geológicas Argentinas, Actas 2: 439-457.
61. Volkheimer, W. 1964. Estratigrafía de la región extrandina del Departamento de Cushamen (Chubut) entre los paralelos 42° y 42°30' y los meridianos 70° y 71°. Revista de la Asociación Geológica Argentina 20: 85-107.
62 Volkheimer, W. 1965a. El Cuaternario pedemontano en el noroeste del Chubut (zona Cushamen). 2° Jornadas Geológicas Argentinas, Actas 2: 439-451.
63. Volkheimer, W. 1965b. Bosquejo geológico del noroeste del Chubut extraandino (zona Gastre-Gualjaina). Revista de la Asociación Geológica Argentina 20: 326-350.
64. Volkheimer, W. and Lage, J. 1981. Descripción geológica de la Hoja 42c, Cerro Mirador, Provincia de Chubut. Secretaría de Estado de Minería, Boletín 181, 71 p., Buenos Aires.
65. Windhausen, A. 1931. Geología Argentina. Editorial Peuser, 2, 486 p., Buenos Aires.
Jorge Strelin1 and Eduardo Malagnino2
1 Instituto Antártico Argentino, Universidad Nacional de Córdoba, Departamento de Geología Básica, Facultad de Ciencias Exactas Físicas y Naturales, Universidad Nacional de Córdoba, E-mail: jstrelin@yahoo.com.ar 2 Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires.
ABSTRACT
Although the depositational environment assigned by Darwin to the large erratic blocks and gravels in the Río Santa Cruz valley has been reinterpreted, his geomorphological and stratigraphic observations are still in force. The large erratic blocks he described as crowning the Condor Cliff terrace and spread at the bottom of the valley just east of this locality (Sites 2 and 3), are now interpreted as indicators of the maximum glacial expansion in Patagonia. Similar blocks, though of a different lithology, accumulated over a lower terrace located up-valley (Site 4), are now linked to moraines and glacifluvial terraces of the Penultimate Glaciation. Finally, in addition to the erratic block discovered by Darwin in the lower Río Santa Cruz valley (Site 1), there are others - recently discovered - which probably account for a catastrophic event ascribed to a big glacier-lake outburst during the last interglacial.
Keywords: Patagonia; Glaciations; Moraines; Erratic blocks; Darwin.
RESUMEN: Charles Darwin y las glaciaciones más antiguas de Patagonia: los bloques erráticos del alto valle del Río Santa Cruz. No obstante haber sido reinterpretado el ambiente depositacional asignado por Darwin a los grandes bloques erráticos y rodados del valle del río Santa Cruz, siguen vigentes sus observaciones geomorfológicas y estratigráficas. Los grandes bloques erráticos que describe coronando la terraza de Condor Cliff y dispersos en el fondo del valle inmediatamente al este de esta localidad (Sitios 2 y 3), son interpretados ahora como indicadores de la máxima expansión glaciaria de la Patagonia. Bloques similares, aunque de dispar litología, acumulados sobre una terraza más baja situada río arriba (Sitio 4), se vinculan actualmente a morenas y terrazas glacifluviales de la Penúltima Glaciación. Finalmente, al bloque errático descubierto por Darwin en el tramo inferior del valle del río Santa Cruz (Sitio 1), se le suman otros -de reciente descubrimiento- que probablemente den cuenta de un evento catastrófico atribuible al vaciamiento de un gran lago glaciar durante el último interglaciar.
Palabras clave: Patagonia; Glaciaciones; Morenas; Bloques erráticos; Darwin.
INTRODUCTION
Although Charles Darwin was a naturalist with a remarkably penetrating, ingenious and broad mind in the field of biology, the not so well-known geologist in Darwin -eclipsed by the former- did not lag far behind. Many of the observations that he made and the geological theories he formulated when still a youth during his scientific voyage to South America and the Pacific Ocean are still current and go beyond pure historical interest. As an example of his innovative theories, it is worth mentioning one which arose from his published observations on coral reefs (Darwin 1842a), which made him famous among geologists. It was only a century later, with the help of modern technology, that his deductions on the great oceanic subsidence could be proved. This work, together with his one on volcanoes (Darwin 1844) and South American Geology (Darwin 1846), make up Darwin's most prolific geological period - from 1841 to 1846. During his journey on board the HMS Beagle, in his own words "one of the most important events" in his life, was when he reached the valley of the Río Santa Cruz. It was here, in 1834 and when only 26 years old, that he made observations of great scientific value. Darwin (1842b) was the first one to describe and discuss the origin of the extensive gravel and shingle layers scattered with erratic blocks, occasionally connected with till deposits, which he discovered during the Beagle's itinerary along the coasts of Patagonia, Tierra del Fuego and Chiloé Island. His interest in these deposits was not fortuitous. Charles Lyell, one of the foremost geologists at the time, had not only sent him a copy of the first edition of his Principles of Geology but had also asked him to pay special attention to the presence of erratic blocks in these regions. By the end of the 18th century, Cuvier's catastrophism theory had been dogmatically accepted by geologists. The first one to seriously oppose it was von Hoff (1834). However, it was Charles Lyell who, following Hoff's steps, introduced actualism in geology. In his Principles of Geology (Lyell 1830), together with his revolutionary contributions to this science and loyal to his uniformitarian ideas, Lyell suggested that the big erratic blocks found in the Northern European plains had been transported there by icebergs in shallow seas and had accumulated on the bottom jointly with non-stratified gravel of chaotic grain sizes which he called till. Just before Darwin's work on gravel beds and erratic blocks in Patagonia was published, Lyell (1840) discussed the origin of this type of erratic blocks in Northern Europe in clear criticism of the neocatastrophic theory proposed by Agassiz (1840), according to which extensive land-grounded ice sheets acted as gatherers of erratic blocks. After his journey to Patagonia and strongly influenced by Lyell (1830-1833), Darwin wrote to his cousin and friend Darwin Fox (July 1835) from Lima, "I have become an enthusiast of Mr. Lyell's ideas and I have tried to explain them in my geological work in South America". Twelve years after Lyell's work, Darwin (1842b) also supported a glaciomarine origin for the extensive gravel deposits and erratic blocks he had discovered in the southern part of South America. Darwin obtained the most complete vision of the geomorphology and stratigraphy of gravel beds and erratic blocks when, together with FitzRoy, he traveled up the valley of the Río Santa Cruz. During this journey, which took place between April 3rd and May 8th, 1834, he focused his descriptions on four sites (Fig. 1). He established the geographic location of these sites by indicating their distance from the Atlantic Ocean and the Andes Mountains in geographical miles. Herein we will see how, 166 years later, Darwin's observations of those sites are still in force and play a key role in the interpretation of the oldest glacial events in Patagonia.
Figure 1: Satellite image of the Río Santa Cruz Valley showing the sites described by Darwin (1842b).
DESCRIPTION OF THE SITES
Site 1 - The Lower Río Santa Cruz Valley
The first isolated erratic block which drew young Darwin's attention as he traveled up the Río Santa Cruz was partially buried in the alluvial plain of the river, 57 miles (105 km) away from the Atlantic Ocean and 110 miles (204 km) from the Cordillera (Site 1 - Fig. 1). However, as he could not find any other similar blocks nearby, he did not consider it important and only described it as a feldspathic rock of about 0.70 m diameter. This feature was pointed out again by Moreno (1879) 45 years later. He also mentioned erratic blocks in the lower valley of the Río Santa Cruz and particularly described a 1m2 block partially buried in thin gravel, two thirds from the top of a terrace level (elevation 40 m a.s.l., i.e., 30 m above the river level) and 6 miles (11 km) away from Isla Pavón. Our recent observations have allowed the discovery of numerous blocks at several locations close to this site. One example are the large erratic blocks, partially buried in alluvium, that can be found on the northern side of the Río Santa Cruz valley, on a terrace at an elevation of 65 and 70 m a.s.l. (40 m above the river valley), 50 miles (91 km) away from the Atlantic Ocean (19 miles - 35 km - from Isla Pavón). The most outstanding are samples 6LA1 of andesitic composition, striated, 1.70 m maximum diameter (Fig. 2) and sample 6LA2 of basaltic composition, 0.80 m maximum diameter. There is also a series of blocks between 1.00 and 0.80 m diameter, partly disaggregated in situ, of rhyodacitic and basaltic composition. East-west oriented fluvial bars, ten to twenty meters long and around 1 to 2 m high, can be observed on the surface of this terrace. Seven erratic boulders up to 1.2 m long have been found to the east of this locality (written communication from Mr. Segovia), at the bottom of a depression which lies partly below sea level. Another block was identified at the foot of a terrace (elevation 40 m a.s.l.) located on the northern margin of the river, near the point mentioned by Moreno (1879) (written communication from Mr. Segovia).
 Figure 2: Erratic block located 15 km downstream of Site 1, at the top of a 65 m. a.s.l. high terrace.
Figure 2: Erratic block located 15 km downstream of Site 1, at the top of a 65 m. a.s.l. high terrace.
Site 2 - The Middle Río Santa Cruz Valley
Darwin (1842b) mentioned the reappearance of erratic blocks 100 miles (185 km) away from the Atlantic Ocean and 67 miles (124 km) away from the Cordillera (Site 2 - Fig. 1), a presence that is continuous and gradually increasing towards the Cordillera. According to our observations, the erratic blocks reappear on the southern side of the valley, 7.8 miles (13 km) east of the point indicated by Darwin. They are partly buried and partly lying on the fluvial terrace at an elevation of 200 m.a.s.l., approximately 100 m above the current river bed. The erratic blocks, which can reach 0.90 m in diameter, are partially weathered and mostly of basaltic and rhyodacitic composition, accompanied by smaller blocks (0.20 - 0.25 m in diameter), some of which seem to be noticeably faceted.
Site 3.- Condor Cliff
It was here, at the narrowest point of the Río Santa Cruz valley, where Darwin (1842b) described and sketched in more detail (Fig. 3a) the morphologic and stratigraphic characteristics of the gravel beds and erratic blocks in Patagonia. Following Darwin's description, the site is located 112 miles (207 km) from the Atlantic Ocean and 55 miles (102 Km) from the Cordillera (Site 3 - Fig. 1). Site 3 was plotted on figure 1 using the intersection of the river with the meridian of 70º50' W, also provided by Darwin (1842b). The distance that we measured following the river track from this geodetic point to the Atlantic Ocean is 116 miles (215 km), only 4 miles longer than the distance measured by Darwin.
 Figure 3: Condor Cliff according to (a) Darwin's interpretation (1842b), (b) Mercer et al. (1975) and (c) the proposal in this paper. 1) Santa Cruz Formation 2) San Fernando Terrace alluvium (Strelin et al. 1999) 3) Condor Cliff Basalts 4) Erratic blocks of the Estancia La Fructuosa Moraine (Strelin1995) 5) post Pliocene gravel strata (Mercer et al. 1975)
Figure 3: Condor Cliff according to (a) Darwin's interpretation (1842b), (b) Mercer et al. (1975) and (c) the proposal in this paper. 1) Santa Cruz Formation 2) San Fernando Terrace alluvium (Strelin et al. 1999) 3) Condor Cliff Basalts 4) Erratic blocks of the Estancia La Fructuosa Moraine (Strelin1995) 5) post Pliocene gravel strata (Mercer et al. 1975)
Darwin's (1842b) observations include a stratigraphic section of the outcrop on the northern side of the valley, which ends at the top of a terrace (at an elevation of 425 m a.s.l.) covered by erratic blocks (Fig. 3a). The covered base of this outcrop begins at an elevation of 90 m a.s.l. with 180 m of small round pebbles composed of clay-slate, feldspathic rock and quartzose chlorite schist. The grain size decreases towards the upper half where thin layers of a variety of colours can be seen. Overlying these are 100 m of basaltic lava which are in turn overlain by 65 m of rounded, coarsely stratified gravel, similar in composition to the underlying gravel but also including basaltic pebbles. Spread over this surface are large blocks, in one case up to 20 m diameter and sticking out 1.8 m above the gravel bed, a block of quartzose chlorite schist of 4.5 x 4.5 m and 1.5 m high, and numerous blocks which range from 0.60 to 1.20 square meters. Darwin (1842 b) mentioned that the top of this sequence is part of a high gravely terrace which reaches 425 m a.s.l. at Condor Cliff, climbing up to 900 m a.s.l. towards the Cordillera, which again does not exceed 2000 m a.s.l. Towards the Atlantic shore this gravel layer, descends slowly down to an elevation of only 245 m a.s.l. This low inclination of the terrace, the absence of mounds and ridges, and the angularity of the boulders lead Darwin (1842b) to postulate a glaciomarine (ice-drifted) origin for this accumulation. One hundred and three years after Darwin's observations, Mercer et al. (1975), in a study of the oldest glaciation in the basins of Lago Argentino and Lago Viedma, drew a new stratigraphic section at Condor Cliff, near the site described by Darwin (1842b). Comparing both sections, it is possible to observe that they only differ in the position of the erratic blocks. According to Mercer et al. (1975) they are located right over the Pliocene basalts (Fig. 3b) while Darwin (1842b), as we have seen, described them as crowning 65 m of gravel. In agreement with Darwin (1842b), Mercer et al. (1975) intercalated the exposures of Pliocene basalt on the northern side of Condor Cliff within two layers of gravel. The 100 m thickness Darwin described for the basalt bed is probably an overestimation caused by basaltic landslides covering part of the northern side of the valley. Mercer et al. (1975) indicated the presence of calcinated gravels at the base of the lava flows corresponding to an alluvial cycle (glaciofluvial) prior to the cycle which accumulated the 100 m of gravel which cover the same flows. By means of radiometric dating they determined a Pliocene age for such lava flows (2.92± 0.07 Ma; 2.79 ± 0.15 Ma; and 2.66 ± 0.06 Ma). According to these authors the basal gravel bed reaches a thickness of 30 m and rests on the Santa Cruz Formation. The 180 m of basal gravel associated with thinner stratified levels described by Darwin (1842b) are probably part of the fluvial facies and tuffaceous levels of the Santa Cruz Formation and the psephitic layer described by Mercer et al. (1975). Our observations (Strelin et al. 1999) agree with Mercer et al.'s (1975) in the sense that the erratic blocks, together with some moraine residue, rest directly on the basaltic flows. The allochthonous blocks found reach up to 3 m in diameter (Fig. 5a) and are mainly of an acidic volcanic nature (rhyodacitic), although some are pelitic and occasionally striated. Autochthonous blocks of basaltic composition are also common. However, we disagree with Darwin (1842b) and Mercer et al. (1975) regarding the stratigraphic relationship between the lava layers and the gravel beds.We have been able to demonstrate that the Condor Cliff basalts are not intercalated between the layers of gravel and that instead they flowed down valleys incised in a terrace level (elevation 500m) which we call La Australasia terrace (Strelin et al. 1999). La Australasia terrace (Figs. 3c and 6) is carved into the Santa Cruz Formation and crowned by gravel. Our conclusion differs from Darwin's proposal (1842b) and Mercer et al. (1975) who considered that the gravel was deposited on the basalts (Fig. 3a and b). The mistake in the interpretation is probably due to the presence of talus deposits formed by gravel which cover the contacts between the lava flows and the base of the Australasia terrace. The basalt channel over a lower terraced level called San Fernando Terrace (Strelin et al. 1999), which at Condor Cliff can reach an elevation between 350 and 400 m a.s.l. This terraced level is also carved into the Santa Cruz Formation and crowned by gravel which appears to be calcinated when covered by basaltic flows. Lava windows are common in this volcanic environment showing intact gravel outcrops with a maximum thickness of 20 metres. In addition to gravel deposits, the sections in the aggraded levels of the San Fernando Terrace show laminated sand beds, tuffaceous and diamictitic levels which include sometimes faceted blocks of up to 0.40 m diameter (Fig. 5b)

 Figure 5: a) Rhyodacitic erratic block located at the top of Condor Cliff; b) Diamictitic level underlying the Condor Cliff lava flows, top of San Fernando Terrace.
Figure 5: a) Rhyodacitic erratic block located at the top of Condor Cliff; b) Diamictitic level underlying the Condor Cliff lava flows, top of San Fernando Terrace.
 Figure 6: Relief model of the Condor Cliff narrow, showing the basaltic flows channeled on the San Fernando Terrace, between the relicts of the Australasian Terrace.
Figure 6: Relief model of the Condor Cliff narrow, showing the basaltic flows channeled on the San Fernando Terrace, between the relicts of the Australasian Terrace.
Site 4.- Upper Río Santa Cruz Valley
Upstream from Condor Cliff, the valley expands into an extensive amphitheatre, which Darwin explained as an ancient estuary of a post Pliocene sea and whose mouth opened towards the west, stressing the marine origin of this section of the valley with the finding of marine shells. The site Darwin (1842b) described is located along a section of the valley located between 127 and 137 miles (235 to 254 km) from the Atlantic coast and 30 - 40 miles (56 - 74 km) from the Cordillera (between sites 4W and 4E, Fig. 1). Both the valley floor (elevation 135 m) and an intermediate terrace (elevation 240 m) are covered with erratic blocks of granitic, sienitic and conglomeratic composition, while no blocks of basaltic composition have been found. In his description, Darwin (1842b) reveals that there is an important difference in the composition of these erratic blocks and those which lie on the upper terrace at Condor Cliff (San Fernando Terrace, Strelin et al. 1999), which he considered evidence of the occurrence of different glaciomarine episodes. Our study (Fig. 4) reveals that the intermediate terrace level that Darwin (1842b) mentions when he described this section of the valley matches the moraine and glacifluvial deposits of the Penultimate Glaciation termed Arroyo Verde (Strelin and Malagnino 1996). The moraine deposits are buried by their own glaciofluvial deposits and the whole set reaches an elevation of 250 m a.s.l., 100 m above the river. The inner slopes of these moraines and glaciofluvial deposits, left behind by ice during the glacial retreat, were modified by an ancient glacial lake (PLA, figure 4). The most prominent lacustrine forms are the paleo-beach levels, bars, spits, lagoons and deltaic deposits. The lithology of the largest erratic blocks which cover the moraine tops is more varied than that of the oldest moraines, and as Darwin (1842b) indicated, blocks of granitic composition appear and basaltic blocks are absent. This particular feature is a result of the level of erosion of the Cordilleran valleys, which reached the granitic substratum (e.g. Fiordo Mayo, Cerro Murallón) in the Cordilleran source area. On the other hand, the decrease in the number of basaltic blocks is related to the decrease of basaltic outcrops upstream. The continental sedimentary sequences of the Santa Cruz Formation crop out along the southern side of the valley, while along the northern side - besides the continental deposits - there are outcrops of marine origin which belong to the Monte León Formation (Patagonian-Patagoniense), with a high content of marine shells, especially oysters and pectinids.
Figure 4: Distribution of the geomorphic units in the Upper Río Santa Cruz Valley region (after Strelin and Malagnino 1996) and Darwin's visited site location.
DISCUSSION AND CONCLUSIONS
Darwin (1842b) incorrectly interpreted the depositional environment of the gravel and erratic blocks which cover the Patagonian plateaus, largely influenced by Lyell's (1830) ideas. Nevertheless, his data allowed the determination -with considerable accuracy- of the extent of the maximum expansion of the glacial advances in the Río Santa Cruz valley. The erratic blocks and moraine remnants deposited to the east and at the foot of Condor Cliff narrow (Sites 2 and 3 respectively, Fig. 4) correspond to these extended glacial advances. Caldenius (1932) and Feruglio (1950) underestimated these observations and did not take into account the location of the erratic blocks mentioned by Darwin (1842b) in Site 2. Therefore, they situated the maximum glacial expansion of the Lago Argentino Basin further up-valley (Caldenius 1932) and at Condor Cliff narrow (Feruglio 1950). Caldenius (1932), for example, considered the Per Dusen Moraines as the oldest (initial glacial) and most expanded system in the Río Santa Cruz valley, with its closure situated at 71º W. Feruglio (1950), on the other hand, referred -in a footnote- to the great erratic blocks at the closure of Cerro Fortaleza (Figs. 4 and 6) as indicators of the maximum glacial expansion in the Río Santa Cruz valley (internal moraines) but, unlike Caldenius, he did not consider them indicators of the oldest glaciation. Likewise, Mercer et al. (1975) thought the glacial advance reached the narrow of Condor Cliff twice. During the first advance of his Most Extensive Glaciation (Mercer et al. 1975) the glacier would have gone beyond the narrow depositing the big erratic blocks which lie on the basaltic cliffs (Figs. 3b and 5a). These blocks would correspond to those mentioned by Darwin (1842b) at the top of his section. During the next advance, the glacier would have deposited the moraines which lie over the western slope of Cerro Fortaleza, at the foot of the basaltic cliffs (Mercer et al. 1975). We agree with Feruglio (1950) in considering that the erratic blocks at Condor Cliff are not the signs of the oldest glaciation in this region (Strelin 1995 and Strelin et al. 1999). Prior to the incision of the valley of the Río Santa Cruz through which the second Patagonian glaciation later channeled (Estancia La Fructuosa Glaciation, Strelin 1995 and Strelin et al. 1999), the Andean glaciers first irrupted in ample lobes along the foothill belt of the Cordilleran front leaving upon their retreat the moraine arches corresponding to the Pampa Alta Glaciation (Strelin 1995) and the glaciofluvial deposits associated and defined as Pampa Alta Proglacial (Strelin 1995). These morphological units make up the top of the plateau at Pampa Alta, located south of the upper valley of the Río Santa Cruz (Fig. 4). Evidence of this first foothill glaciation is also present at other localities in the Patagonian Andes. In the Lago Buenos Aires basin for example, Malagnino (1995) identified the Chipanque Moraines, as a set of three systems made up of 27 belts located on the Guenguel Plateau at an elevation of 900 meters. These were deposited by a glaciation (Chipanque Glaciation, Malagnino 1995) which had not been identified before in the Lago Buenos Aires basin. With this discovery it was possible to raise to six the number of glaciations in this area of the Patagonian Andes and its foothills. After the Pampa Alta Glaciation and before the maximum glacial expansion in the Río Santa Cruz valley (Estancia La Fructuosa Glaciation, Strelin 1995) there was a long interglacial period during which the course of the Río Santa Cruz was formed, producing a deep canyon which was later partially filled with basaltic lava flows during the Late Pliocene. We have been able to demonstrate that the referred basaltic volcanism took place at two different eruptive periods: i.e., before and after the accumulation of the moraines which cover the terrace at Condor Cliff (Strelin et al. 1999). The terrace level on which the lava flows are channeled coincides with the ancient tributary valleys of the ancestral Río Santa Cruz, the valley floors of which formed the current San Fernando Terrace (Strelin et al. 1999). The entrenchment of the tributary valleys, adjusted to the old base level of the Río Santa Cruz, continued until it reached an elevation of 200 -250 m a.s.l. (75 -135 m above the current level of Río Santa Cruz). This can be verified right in front of Condor Cliff at the foot of Cerro Fortaleza, along the southern margin of the narrow, where 12 basaltic lava flows with a total thickness of 100 m lie (through pillow lavas and palagonite tuffs) over the layers of the Santa Cruz Formation. To the west of the Condor Cliff narrow the valley opens into an amphitheatre which Darwin (1842b), as previously mentioned, interpreted as a terraced paleo-estuary (intermediate terrace, elevation 240 m a.s.l.) opening towards the Cordillera. He described it as abundant in erratic blocks with a different composition from those on the upper terrace of Condor Cliff and belonging to a different glaciomarine event from that which deposited the blocks on the upper terrace. Darwin (1842b) thought to confirm the marine environment with the finding of shells on the valley floor (elevation 135 m a.s.l.). This interpretation strongly contrasts with ours at first sight because we consider that Darwin's intermediate terrace is part of the moraine and glaciofluvial deposits formed by the glacial tongue which channeled through the Río Santa Cruz valley during the Penultimate Glaciation (Arroyo Verde Glaciation, Strelin and Malagnino 1996). However, if we analyze the landscape features in detail we will see that Darwin (1842b) did not let his imagination run away. On the one hand, there is the fact -well-established by Darwin- of a different glacial event based on the different composition of the erratic blocks. On the other hand, we can justify his interpretation of the existence of a paleo-estuary (paleo-fjord) in the upper valley of Rio Santa Cruz if we consider the morphological similarities between this valley and other glacial valleys, totally or partially flooded by the sea, visited by Darwin during his voyage to the southern seas (Strait of Magellan, Otway and Skyring Fjords, San Sebastián Bay, Inútil (Useless) Bay, Beagle Channel, etc). The morphological similarity with flooded valleys is even more noticeable if we consider that the section of the valley Darwin (1842b) examined was reshaped by an ancient lake as shown by the lacustrine paleo-forms (raised beaches, bars, spits, lagoons and deltas, in figure 4) hanging at elevations of up to 250 m a.s.l. (100 m above the valley floor). This lake formed as a result of a moraine damming (Arroyo Verde moraines, Strelin and Malagnino 1996) during the glacial retreat which took place in the Paleo-Lago Argentino Interglacial (Strelin and Malagnino 1996). As regards the marine shells Darwin discovered in this section of the valley, Feruglio (1950) suggests that they may have been abandoned there by natives. We consider, however, that it is more likely that Darwin (1842b) found reworked oyster and pectinid shells coming from the coquina exposures of the Monte Leon Formation located near this site. It is worth mentioning that the surfacing of Monte Leon Formation in this part of the valley is a feature which has yet not been clearly explained. Finally, the origin of the erratic blocks (Fig. 2) found in the lower valley of the Río Santa Cruz (Site 1, Fig. 1) has not been elucidated yet. Darwin (1842b) was sensitive to this enigma, which he tried to solve when he suggested that they could have been accumulated after rafting over fluvial ice. At present we consider this feasible and furthermore that it could have been after the catastrophic draining of the ancient Arroyo Verde morainedammed glacier-lake (Strelin and Malagnino 1996).
ACKNOWLEDGMENTS
We are especially grateful to Marcos Mozetic, Eduardo Olivero, and Michael Kaplan for their suggestions and for improving the english of this paper. Also we thank very much Beatriz Aguirre-Urreta for inviting us to participate in this special issue dedicated to Charles Darwin.
WORKS CITED IN THE TEXT
1. Agassiz, L. 1840. Études sur les glaciers. Jent et Gassmann Libraires, 346 p., Neuchâtel.
2. Darwin, C. 1842a. The structure and distribution of coral reefs. Being the first part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 214 p., London.
3. Darwin, C. 1842b. On the distribution of erratic boulders and on the contemporaneous unstratified deposits of South America. Transactions Geological Society London 6: 415-431.
4. Darwin, C. 1844. Geological Observations on The Volcanic Island, visited during the Voyage of H. M. S. Beagle. The Geology of the Voyage of the Beagle, Part 2, Smith Elder and Co. 172 p., London.
5. Darwin, C. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 280 p., London.
6. Feruglio, E. 1950. Descripción Geológica de la Patagonia. Yacimientos Petroliferos Fiscales 3, 431 p., Buenos Aires.
7. Hoff, K.E.A. von 1834. Geschichte der durch Ueberlieferung nachgewiesenen natürlichen Veränderungen der Erdoberfläche, 1-4, Perthes, 2086 p., Gotha.
8. Lyell, C. 1830-33. Principles of Geology. John Murray, Albemarle-Street, (1830) 1, 511 p.; (1832) 2, 330 p.; (1833) 3, 109 p., London.
9. Lyell, C. 1840. On the Boulder Formation, or drift and associated freshwater deposits composing the mud-cliffs of Eastern Norfolk. The London and Edinburgh Philosophical Magazine and Journal of Science, Third Series 16(104): 345-380.
10. Malagnino, E.C. 1995. The discovery of the oldest extra-Andean glaciation in the Lago Buenos Aires Basin, Argentina. A.A. Balkema, Quaternary of South America and Antartic Peninsula 9: 69-83, Rotterdam.
11. Mercer, J.H., Fleck, R.J., Mankinen, E.A. and Sander, W. 1975. Southern Patagonia: Glacial Events Between 4 m.y. and 1 m.y. Ago. In Suggate, R.P. and Creswell, M.M. (eds.) Quaternary Studies, The Royal Society of New Zealand, 223-230, Wellington.
12. Strelin, J.A. 1995. New evidences on the relationships between the oldest estra-andean glaciations in the the Santa Cruz River area. A.A. Balkema, Quaternary of South America and Antarctic Peninsula 9: 105-116, Rotterdam.
13. Strelin, J.A. and Malagnino, E.C. 1996. Glaciaciones Pleistocenas del Lago Argentino y Alto Valle del Río Santa Cruz. 13° Congreso Geológico Argentino, Actas 4: 311-326.
14. Strelin, J., Re, G., Keller, R. and Malagnino E. 1999. New evidences concerning the Plio-Pleistocene landscape evolution of southern Santa Cruz region. Journal of South American Earth Sciences 12: 333-341.
From Buenos Aires to Santa Fe: Darwin's observations and modern knowledge
Martin Iriondo and Daniela Kröhling
CONICET - Universidad Nacional del Litoral, CC 217 (3000) Santa Fe. E-mails rniriond@ceride.gov.ar, dkrohli@fich1.unl.edu.ar
ABSTRACT
During his historical voyage around the world, Darwin raided deeply in the South American interior, travelling over 600 kilometers from Buenos Aires to the north along the Río Paraná. During that journey, he crossed a vast plain characterized by aeolian sediments, something unfamiliar to a European naturalist. However, Darwin's acute observation powers and precise descriptions are noteworthy. After more than 170 years since his visit, modern geological knowledge identifies several sectors in the Buenos Aires-Santa Fe region. One of them (the Tertiary at La Bajada) he described admirably and others such as the Paraná flood plain were brilliantly abstracted in only two sentences. In short, Darwin traversed a first sector (Buenos Aires-Rosario) characterized by aeolian and paludal Early Pleistocene sediments. From Rosario to Santa Fe the plain is formed by Late Pleistocene aeolian and fluvial units. At La Bajada (presently Paraná city) lies exposed the marine Miocene and in SW Entre Ríos is a reconstructed loess-paleosol sequence generated at the Early/Middle Pleistocene transition. The Paraná flood plain and the littoral complex at the mouth (practically not observed by Darwin) underwent rather complex Holocene episodes.
Keywords: Ch. Darwin; Pampa; South America; Quaternary; Tertiary.
RESUMEN: De Buenos Aires a Santa Fe: observaciones de Darwin y conocimiento actual. Durante su histórico viaje alrededor del mundo, Charles Darwin incursionó profundamente en el interior de Sudamérica recorriendo más de 600 kilómetros hacia el norte de Buenos Aires a lo largo del río Paraná. Durante ese viaje, él recorrió una gran planicie caracterizada por sedimentos principalmente eólicos, un caso poco familiar para un naturalista europeo. Sin embargo, sobresale su gran capacidad de observación y precisión en sus descripciones. Después de más de 170 años de la visita de Darwin, el conocimiento geológico moderno ha identificado varios sectores en la región Buenos Aires-Santa Fe, uno de los cuales (el Terciario de La Bajada) él describió admirablemente y otros, como la llanura aluvial del Paraná, reseñó magistralmente en solo dos frases breves. En resumen, Darwin recorrió un primer tramo (Buenos Aires-Rosario) caracterizado por sedimentos eólicos y palustres del Pleistoceno Inferior. Desde Rosario hasta Santa Fe la llanura está formada por unidades eólicas y fluviales de edad pleistocena superior. En La Bajada (hoy ciudad de Paraná) aflora el Mioceno marino y en el sudoeste de Entre Ríos se reconstruyó un sistema loess-paleosuelos generado a lo largo de la transición Pleistoceno Inferior-Pleistoceno Medio. La llanura aluvial del Paraná y el complejo litoral de su desembocadura (prácticamente no observados por Darwin) sufrieron cambios dinámicos bastante complejos durante el Holoceno.
Palabras clave: Ch. Darwin; Pampa; Sudamérica; Cuaternario; Terciario.
INTRODUCTION
Charles Darwin made the journey from Buenos Aires to Santa Fe by cart and horse. After several days of work at La Bajada (presently Paraná city) he returned to Buenos Airs by boat along the Río Paraná. Such a trip allowed him (surely by chance) to cross four major geographic and Quaternary geological environments of southeastern South America (Fig.1). The first one stretches from Buenos Aires city (in fact, it begins further south) to Rosario and reaches several tens of kilometers westwards. It is formed by a slightly elevated block characterized by Early Pleistocene formations and marginal Holocene littoral deposits. The second sector covers the Rosario-Santa Fe road and reaches 300 km west to the hills in Córdoba. This system is a relatively sunken region dominated by Late Quaternary loess bounded at the north by the South American region of Chaco. The third area he visited forms the southwestern sector of Entre Ríos province; it is represented by a hilly landscape carved into Miocene marine sediments and Early Quaternary aeolian and swampy formations. The last environment Darwin recorded during his journey is the Paraná river system, composed of a wide flood plain and a littoral complex at the mouth ("Paraná delta"), both of them Holocene in age.
 Figure 1: Map of the region showing the localities visited by Darwin
Figure 1: Map of the region showing the localities visited by Darwin
In order to visualize Darwin's experiences, this article is organized in the same sequence of geological terrains that he found during his journey (Darwin 1846, 1945). When reading his journal or his geological observations of the region, Darwin's great observation power and exceptional ability for synthesis appear immediately evident, notwithstanding the limitations of geological theory at that time. For instance, the nature and origin of loess (the dominant sediment in the Pampas) was discovered several decades later. It is evident that Darwin did not feel comfortable with such "muds", "clays" and similar. In order to describe and interpret those extensive deposits of the plain, he preferred to hold on to strata and concretions of tosca (caliche), intercalated in the sections. His skill as a paleontologist in comparison with his abilities as a sedimentologist is also clear. Besides, it is interesting to discover in Darwin's writings the existence of qualified observers - i.e., people with some scientific knowledge - among the local population in those early years of Argentina. Some of them were British traders, but others were local inhabitants that were great observers of Nature.
THE GREAT DROUGHT
An interesting environmental issue was described by Darwin in relation to the trip to Santa Fe. It was the "Gran Seca" (the great drought) that particularly affected the north of Buenos Aires and south of Santa Fe from 1827 to 1832. During those five years rain was exceptionally scarce and the plain underwent major changes: vegetation disappeared, minor rivers dried up and the entire country took the aspect of a dusty road, dust clouds continuously dominating the air. The effect on animals was catastrophic; cattle migrated massively to the south and assembled there in a giant flock; droves with thousands of wild horses hurried furiously to the Paraná to drink and later were unable to climb back up the slippery cliff and drowned. Hundreds of thousands of animals floated downstream to the Plata estuary. A few years later Darwin observed ¨true layers of bones¨ covering the bottom of some tributaries. Very probably such great drought was a short late occurrence of the Little Ice Age (LIA) climate, which was characterized by a marked dryness in the Pampas (Parra 1939, Iriondo and Kröhling 1995) and ended around 1800-1810. The Spanish naturalist Azara also described similar events in the late 18th Century. The Little Ice Age provoked advances of glaciers in the Cordillera and related processes in southern South America, but did not produce visible geological effects in the Pampas, probably owing to its relatively short duration and not so severe dryness. On the other hand, a longer dry period occurred between 3.5 and 1.4 ka BP and formed a thin aeolian mantle which covers the Pampas and surrounding regions, i.e., the San Guillermo Formation (Iriondo 1990, Fig. 2). It is an aeolian gray silt that generally tops the sedimentary sequence in the large interfluves. Most of the sediment is originated locally, by deflation of the A-horizon of soils and the subsequent deposition of dust. In Santa Fe province this mantle is formed by coarse silt with scarce proportions of very fine sand and clay, brownish gray in color (10YR 5/1). It is friable, porous, permeable, and moderately structured. Typical thickness varies from 25 to 55 cm; ceramic shards and Indian boleadora balls are included in the upper part of this deposit at some localities.
 Figure 2: Map of the San Guillermo Formation, a late Holocene environmental equivalent of Darwin's big drought (after Iriondo 1990).
Figure 2: Map of the San Guillermo Formation, a late Holocene environmental equivalent of Darwin's big drought (after Iriondo 1990).
BUENOS AIRES/ROSARIO ELEVATED BLOCK
Darwin rode a 250 km long first journey from Buenos Aires to Rosario in a straight northwestern direction. All that time, the road borders the right banks of the Río de la Plata and Río Paraná and runs atop a 10 to 20 m high cliff. This cliff is almost continuous, interrupted in a few places by small Holocene estuaries of minor tributaries (Luján, Areco, Arrecifes and others; Iriondo 2004). The cliff is tectonic in origin and exposes two Quaternary formations described by Darwin as "pale and red Pampean mud". Both units were identified and studied later in the Twentieth Century. González Bonorino (1965) refered the sections outcropping in the Buenos Aires area as the Pampeano Formation. According to its mineralogy, the author differentiated two zones separated by a clear boundary: the upper zone characterized by plagioclase and illite and the lower one dominated by quartz and montmorillonite. Fidalgo et al. (1973) later described the Pampiano Formation and Riggi et al. (1986) discriminated the Ensenada Formation and the Buenos Aires Formation in the same sections. Both units appear along all the distance to Rosario, but have better outcrops on the left bank of the Paraná, in Entre Ríos province. Here they were defined as Punta Gorda Formation (Iriondo and Kröhling 2008) and Hernandarias Formation (Iriondo 1980). Pale Pampean Mud/Punta Gorda Formation is an Early and Middle Pleistocene loess-paleosol sequence. It is composed of the Andean plagioclase-illite association and has an origin similar to the classical Last Glacial Maximum Pampean loess (Kröhling 2001). Both minerals were transported by southwesterly winds from the Andes Cordillera. The Punta Gorda Formation was deposited around 1 Ma BP during the most important Quaternary glaciation occurring in South America (Mercer 1976). The Red Pampean Mud/Hernandarias Formation is a large playa deposit deposited by the Río Uruguay at the Early-Middle Pleistocene transition. It forms the surface of most of Entre Ríos province and below the surface reaches into neighboring regions in Santa Fe and Buenos Aires to the southwest (Iriondo 1980). The typical thickness of this formation varies from 20 to 40 m; it is composed of loam and silty loam with montmorillonite as main clay mineral, and quartz and very fine sand containing gypsum at the base. More than 90 % of the clay minerals, i.e., montmorillonite, beidellite and nontronite, show expansion/contraction properties. The contact between both formations is concordant - a feature correctly observed by Darwin, who stated: "I could clearly distinguish in this fine line of cliffs horizontal lines of variation both in tint and compactness" (Darwin 1846, p. 87). As remarked by Darwin this observation contradicts the insistent assertion of the French naturalist d'Orbigny on the general absence of stratification in the Pampean sediments. The academic point was then whether the Pampean System was diluvial or not: if stratified, not diluvial.
Baradero Section (NE Buenos Aires province)
The section on the Río Baradero north of the city of Baradero (100 km from Buenos Aires), is formed by the Punta Gorda Formation (5 m thick), conformably overlain by the Hernandarias Formation (7 m thick, Iriondo 1980). From bottom to top: 0 - 1 m Punta Gorda Formation. Bed of clayish silt, dull brown in colour, moderately consolidated. It contains numerous Fe-oxide segregations, CaCO3 concretions of different sizes and large rhizoconcretions. It is interpreted as a paludal deposit. 1 - 5 m Punta Gorda Formation. Bed of a silty grain-size composition, pale brown in colour, massive in general and in some places with a weak medium to coarse horizontal stratification. Medium to high consolidation (highly calcareous); carbonate (tosca) concretions. Fine root moulds covered by black films. The deposit has a general aeolian origin, with local rill reworking and krotovinas in some places. Remains of Glyptodon were found in the measured section. At the top, a marked horizontal paleosurface is recognized, locally joined to a residual material of 0.60 - 0.80 m thick, with few local pedogenic features. 5 - 12 m Hernandarias Formation. It is composed of clayish silt to a silty clay, olive gray in general colour and with variations to brown and olive; with abundant large and irregular carbonate concretions. Frequent Fe-Mn sesquioxide segregations appear. It is less consolidated than the underlying unit and is organized in poorly defined coarse strata, and interpreted as a paludal deposit. Numerous dessication cracks appear along all the section. The deposit includes medium to coarse lenticular to tabular strata of pale brown silt, with internal lamination and segregated CaCO3. Such lenses are interpreted as aeolian, locally retransported material. In some sectors appear lenses of retransported botrioidal CaCO3 concretions. The forms of erosion of both formations are very different and distinctive along the river cliffs. They were also affected by several gullies with. 12-15 Tezanos Pinto Formation. Yellowish brown, friable, massive and calcareous silty deposits. It is the typical LGM Pampean loess. Nabel et al. (1993) studied the magnetostratigraphy of two sections at Baradero, finding reverse polarity (older than 0.78 Ma) in the lower section of the cliff. Those authors identified two of the lenticular paleosols and named them El Tala (Brunhes paleomagnetic chron) and Hisisa Geosols (Matuyama chron > 780 ka). Nabel et al. (1993) discriminated eight litoestratigraphic units at Baradero section: units I to V interpreted as loess and paleosols deposits and units VI to VIII associated with limnic environments. The age of the deposit in the region was attributed to the Ensenadense Mammal Age (Tonni et al. 1999). Later, Kemp et al. (2006) reconstructed the sequence of pedosedimentary processes in the same locality based on soil science methods and identified paleosols embedded in swamp and aeolian sediments. Their ages as recorded by luminescence methods (OSL dates range from ca. 114 to 14 ka BP) differ considerably from previous paleomagnetic results of Baradero.
San Pedro Section (NE Buenos Aires province)
In a quarry excavated into the Río Paraná cliff at San Pedro the following section appears from bottom to top: a paleosol developed on top of the Punta Gorda Formation and truncated by erosion (1.20 m thick) and constituted by brown silt, weakly structured in peds. It contains numerous large root moulds generated by the lixiviation of CaCO3 rhizoconcretions. Pedological features are: fine root moulds covered by black films, argillocutans and ferriargillans. This paleosol corresponds to the unit G attributed by Imbellone and Cumba (2003) -Geosol El Tala-Hisisa-. The Hernandarias Formation forms the rest of the exposed section, with general paludal characteristics and interbedded discontinuous soil horizons in the middle sector of the unit. Numerous large krotovinas appear in this formation.
Vuelta de Obligado and Ramallo Sections (NE Buenos Aires province)
The section of the cliff at Vuelta de Obligado is represented by the loessic facies of the Punta Gorda Formation (6.50 - 8 m thick), overlain by a calcrete (2.50 m thick). The unit includes a 1.30 m thick paleosol at the base, composed from top to bottom as follows: 0 - 0.35 m B2t horizon. Brown clayish silt (7.5 YR 5/4), moderately to well structured in very firm, fine to medium prisms, bounded by fine fissures. Common clay cutans and fine root moulds. Very fine to fine Fe-mottles and Fe-Mn oxides segregations frequent. 0.35 - 0.57 m B3 horizon. Brown silt moderately organized in very firm, fine to medium prisms. Scarce argillocutans and fine root moulds. 0.57 - 0.87 m BC horizon. Brown silt, weakly structured in firm and fine blocky peds. Abundant very fine Fe-Mn mottles and fine root moulds. 0.87 - 1.29 m C horizon. Light brown clayish silt with paludal characteristics (very fine resistant blocky peds, frequent Femottles, common CaCO3 irregular and platy concretions). At that locality, a well developed 1.5 m high notch appears at the base of the cliff; that feature indicates erosional action by waves during the Middle Holocene marine ingression. Such a feature also appears in the same position at Fray Bentos, in the Banda Oriental (Uruguay). The Río Paraná cliff at the locality of Ramallo is formed by the Punta Gorda Formation, represented by the loessic strata with frequent CaCO3 vertical platy concretions, 2.50 to 3 m thick. The unit is covered by the paludal facies of the Hernandarias Formation, with frequent CaCO3 concretions.
Arroyo Pavón and Arroyo Seco Sections (Southern Santa Fe province)
Entering the territory of Santa Fe province, Darwin described a cascade about twenty feet high in the Pavón creek, a small tributary of the Paraná. He recorded there two varieties of tosca-rock (limestone). Today the Pavón Cascade is located a few hundred meters from Provincial Highway 11. The cliff downstream the cascade begins at the base with a brown sandy aeolian deposit (1.5 m thick) covered by a loessic unit with segregated CaCO3 (1.5-2 m thick). A paleosol (1 m thick) is recognized on top of it (it very probably correlates with the soil exposed at the bottom of the Vuelta de Obligado section). The section ends with the loessic facies of the Punta Gorda Formation with abundant CaCO3 concretions (2 m thick). Some kilometers to the north, the Punta Gorda Formation forms the banks of the Arroyo Seco (9 m high at the town's camping grounds); locally differentiated is a 3 m thick brown silty strata, paludal in origin.
ROSARIO/SANTA FE BLOCK
From Rosario to Santa Fe and beyond to the north and west, stretches another sector of the Pampas, dominated by Late Pleistocene geological formations. The landscape is there shaped by two widespread aeolian units: Tezanos Pinto For-mation and San Guillermo Formation. However other units originated in fluvial environments have been described. The stratigraphic sequence is the following:
Section of the Paraná cliff between Puerto Gaboto and Rosario (Santa Fe province)
According to Kröhling (1999) the exposed stratigraphic column on the cliffs along the right bank of the Paraná and Coronda rivers, between the cities of Puerto Gaboto and Rosario, begins at the base with the Plio-Quaternary Ituzaingó Formation, deposited by the Río Paraná. The sedimentary record of the unit is represented by mature and well selected sands, typical of channel facies of a fluvial system of high discharge. The conditions that favored the accumulation of the unit changed toward the top of the formation to a fluvial regime of lower energy. The Ituzaingó Formation is separated by an erosional unconformity from the overlying Late Pleistocene Puerto San Martín Formation, formed by the discontinuous accumulation of fine aeolian sediments. The accumulation environment was of semipermanent swamps, intercalated with periods of subaerial conditions (loess). The existence of Ck-horizons of truncated soils near the top of the unit suggests more benign conditions. In some sites, between the two mentioned formations there are intercalated several paleochannels infilled with alluvial and paludal deposits. These were formed by successive avulsions of the last segment of an important tributary in the region - the Río Carcarañá (Timbúes Formation). The landscape developed over the Puerto San Martín Formation was later covered and smoothed during the Last Glacial Maximum by the loess of the Tezanos Pinto Formation. The stratigraphic sequence ends with the Late Holocene San Guillermo Formation, which lies over the buried Holocene Optimum Climaticum soil developed on top of the loess. Darwin described the Paraná cliff in the area of Estancia Grondona, north of Rosario and near the last and more recent segment of the Río Carcarañá. The exposed section was characterized by Darwin as follows, from the base to top: "it consists of a pale yellowish clay, abounding with concretionary cylinders of a ferruginous sandstone". It is interpreted as the Ituzaingó Formation. Darwin continues: "The rest of the cliff at Gorondona, is formed of red Pampean mud, with, in the lower part, many concretions of tosca, some stalactiformed, and with only a few in the upper part… containing mammiferous remains close to its base" (Darwin 1846, p. 87). It corresponds to the general features of the Puerto San Martín Formation. The main characteristics of the mentioned formations exposed in the area visited by Darwin were taken from Kröhling (1998) and are presented below: Ituzaingó Formation (De Alba 1953, Herbst 1971) It is composed of very fine quartz sand, sandy silt to silty clay, grayish yellow in colour (2.5Y 6/2) with variations to olive yellow (5Y 6/3). The sediment has abundant ferric segregations; also the postdepositional rubefication is in general discordant to the stratification. The upper part of the unit has abundant brown Fesesquioxides mottles (7.5 YR 5/8), locally concentrated in centimetric levels. The consistence of the sedimentary mass ranges from friable to poorly consolidated non-calcareous. Segregations of CaCO3 form irregular to botrioidal concretions. Locally, in the lower part there are platty CaCO3 concretions forming an orthogonal net or ferruginous concretions of 30 to 50 mm diameter. The unit is arranged into medium to coarse massive lenticular strata with 7-10 m of lateral extension, with variable concentrations of very fine to medium Fe-mottles (up to 40%) and Mn-segregations and mottles. Abundant fine root moulds with black segregations. In some sectors, the top of the unit is marked by a moderately developed B horizon and by calcareous concretions bellow it. Locally, the truncated paleosol is replaced by medium cuneiform strata composed of fine sand, ochre-yellow in colour, that form a set with diagonal stratification. Upwards they are replaced by medium horizontal strata composed of intraclasts in a sandy matrix or by internally laminated strata. This unit, represented by the fluvial channel sedimentary facies (paludal facies is common too), outcrops at the base of the cliff of the Río Paraná from its junction with the Río Coronda up to the city of Rosario, with variable thickness between 0.5 m and 4.5 m. Locally the formation is covered by the Timbúes Formation separated by an erosional unconformity, but in general the overlying unit is the Puerto San Martín Formation. Main characteristics of the Ituzaingó Formation in its type area (Corrientes province) are presented in Georgieff et al. (2005). Timbúes Formation (Kröhling 1998) It is composed of silty very fine to fine sand, opaque orange in colour (7.5 YR 7/4), with visible micaceous minerals (frequency of 5-20%). It is organized in coarse to very coarse, internally laminated strata, lenticular to tabular in form, with irregular concordant to erosional contacts among them. Deformational structures are common. There are Mnsegregations, root moulds filled by sand and leaf moulds covered by Mn-films. Platty CaCO3 concretions appear at the contact between some strata, locally forming a calcareous net of phreatic origin. Large elliptic krotovinas are frequent. At the Paraná cliff near the locality of Puerto San Martín, a B horizon of a paleosol appears at the erosional contact between the Timbúes Formation and the overlying Tezanos Pinto Formation. It is 0.30 to 0.60 m thick, reddish brown in color and weakly to moderately structured in medium angular prisms. The Timbúes Formation (Late Pleistocene in age) outcrops at the cliff of the Paraná and Coronda rivers, exhibiting a thickness of 5 to 6.5 m and lateral extensions of tens of meters. The sedimentary characteristics indicate a fluvial origin for this unit. It is interpreted according to mineralogical data as a deposit generated by the Río Carcarañá and indicating the position of its previous mouths in the Paraná. Puerto San Martín Formation (Iriondo 1987) The unit is practically continuous along the Río Paraná cliff from the mouth of the Río Coronda up to the city of San Lorenzo. Its thickness varies from 4 up to 10 m. The unit is overlain by the Tezanos Pinto Formation, separated by an unconformity that indicates an irregular paleosurface. It is a yellowish brown silty deposit, organized in coarse to very coarse horizontal strata with poorly defined contacts. At Puerto San Martín - the type locality - the section is as follows from bottom to top: 0-0.60 m: bed composed of brownish olive silt, structured in peds, paludal in origin. 0.60-1.60 m: bed of yellowish brown silt, massive, aeolian in origin. 1.60-2.10 m: bed formed by brownish olive silt, organized in peds and accumulated in a swampy environment. 2.10-3.30 m: yellowish brown loess. 3.30-4.80 m: bed composed by yellowish brown silt, moderately structured, paludal in origin. 4.80-5.10 m: brownish olive silt, weakly structured, paludal in origin. 5.10-6.40 m: stratum of yellowish brown silt, weakly structured in angular peds. Locally a level with CaCO3 rhizoconcretions appears immediately below the erosional unconformity at the top of the unit indicating the existence of a Ck horizon of a truncated soils. At the Río Paraná cliff in front of the city of San Lorenzo, two sectors can be differentiated: the lower one is 4 m thick and formed by olive sandy silt with common ferruginous mottles. The upper part carries calcareous precipitates of phreatic origin. The upper sector is 7 m thick and composed of massive, yellowish brown silt, including a pedogenic horizon marked by CaCO3 concretions. The Puerto San Martín Formation outcrops at sectors of the Río Carcarañá cliff near the town of Oliveros. On the left bank the unit is also differentiated into two sectors separated by a concordant horizontal contact. The lower silty sector, olive in colour, comprises the Ck horizon of a truncated paleosol. The upper sector is represented by a brownish to reddish brown silt with diffuse crossed stratification. It includes a lenticular fine bed formed by CaCO3 pebbles. A moderately structured B horizon of a truncated soil tops this unit. The sedimentary unit is interpreted as a continuous sequence of strata representing a temporary swampy environment alternated with typical aeolian facies that suggest an important accumulation of dust in the region. Taking into account the reference of Darwin about the preponderant number of fresh-water species (Polygastrica and Phytolitharia) scraped from a tooth of one of the mastodons found in the red Pampean mud, we infer that this corresponds to one of the characteristics beds of paludal origin of the Puerto San Martín Formation. Tezanos Pinto Formation (Iriondo 1987) This formation is the typical LGM loess of the northern Pampas that overlies the Puerto San Martín Formation, with thickness ranging between 1 and 4 m. San Guillermo Formation (Iriondo 1987) The brownish gray silty deposit (Late Holocene in age) ends the sedimentary sequence on a truncated soil developed on top of the LGM loess.
Sections of the lower Río Carcarañá (Santa Fe province)
Darwin was right in making his observations along the banks of the lower Río Carcarañá because its basin constitutes a favorable area for research on geomorphology and Quaternary stratigraphy of the northern Pampas plain (Kröhling 1999).The exposed sedimentary column of the lower Carcarañá basin reveals fluctuating environmental conditions, characterized by dry intervals associated with episodes of accumulation and remobilization of aeolian sand or dust accumulation during the stages of Glacial Maximum. These alternate with humid intervals characterized by pedogenesis and the development of fluvial belts. The sedimentary record of the lower basin was reported by Kröhling (1999) and is explained below (Fig. 3).

 Figure 3: General stratigraphic section of the Carcarañá river basin, typical of the Rosario-Santa Fe area (after Kröhling 1999).
Figure 3: General stratigraphic section of the Carcarañá river basin, typical of the Rosario-Santa Fe area (after Kröhling 1999).
Carcarañá Formation (Kröhling 1999) This unit is composed of very fine to fine silty sand, and silt with very fine sand, dull orange in colour (7.5 YR 6/4). It has very thick horizontal strata, not well defined and generally with concordant contacts. The sediment varies in consistency between friable and consolidated and it has a fine to medium blocky structure. In general, it is non-calcareous. It is affected locally by bioturbations, such as krotovinas (up to 0.75 m diameter) and root casts (up to 0.60 m long and 0.10 m diameter). In some places sedimentary structures are visible. These are characterized by an irregular wavy pattern marked by differences in concentrations of colloidal materials, namely dissipation structures. This unit forms the lower sections of cliffs along the Río Carcarañá, forming gentle to subvertical slopes, which reflect its resistance in comparison with the upper units. The maximum outcrop thickness is approximately 5.50 m. It is unconformably overlain by the Tezanos Pinto Formation in the interfluves and by the Lucio Lopez Formation in the main fluvial valleys of the area. A representative section of the unit is exposed along the cliffs of the Río Cañada de Gómez, near the Río Carcarañá. From bottom to top: 0.00-2.00 m: Bed formed by very fine, slightly silty sand, bright reddish brown in colour when moist, with dissipation structures. The sedimentary mass is noncalcareous and contains medium to coarse and hard Fe-Mn sesquioxide nodules. There are rhizoconcretions of CaCO3 and abundant very fine macropores. Concordant upper contact. 2.00-4.50 m: Bed sedimentologically similar to the underlying one, without sedimentary structures, and with less resistance to erosion. Numerous CaCO3 rhizoconcretions appear at the contact between the units. It includes krotovinas up to 0.50 m in diameter. Near the top there is a level formed by a succession of erosional geoforms, 0.50-0.80 m wide and 0.40-0.50 m high. Relicts of a palaeosol appear discontinuously at the bottoms of gullies within the fluvial palaeovalleys of the region. Lateral variations are common in that soil: the structure changes from coarse to very coarse blocky (strong) up to angular very coarse prismatic (strong, defined by coarse fissures, partly filled by CaCO3 precipitates). The sediment is weakly calcareous to calcareous. There are numerous epigenetic carbonate concretions produced by partial dissolution. CaCO3 concretions of phreatic origin occupy coarse fissures between peds. Locally, this level is represented by palaeogullies filled with retransported soil fragments. At the top of the Carcarañá Formation another soil was developed. It is represented by a poorly developed B horizon, 0.40 m thick, dark brown in colour, weakly structured and containing CaCO3 rhizoconcretions. The Carcarañá Formation is Late Pleistocene in age (OIS 3; a TL dating in the middle section indicates an age of 52.31 ± 1.2 ka BP). The mainly aeolian formation is the result of the reworking by erosion of a dune field generated during the OIS 4. Primary structures of the dunes appear in sections located immediately to the south of the area. Locally different paludal and alluvial facies of the Carcarañá Formation are present. According to Darwin, "on the banks of the Carcarañá, a few miles distant (of the Paraná cliff at Grondona; next to the Villa La Ribera -Rosario-Santa Fe road-), the lowest bed visible was pale Pampean mud, with masses of tosca-rock, in one of which I found a much decayed tooth of the Mastodon: above this bed, there was a thin layer almost composed of small concretions of white tosca, out of which I extracted a well preserved, but slightly broken tooth of Toxodon Platensis: above this there was an unusual bed of very soft impure sandstone" (Darwin 1846, p. 88). The section of the Río Carcarañá described by Darwin is interpreted as the Carcarañá Formation. Tezanos Pinto Formation (Iriondo 1980) It is the typical Late Quaternary unit of the Pampas plain, composed of aeolian silts (silt: 71-81%; clay: 13-29%; fine sand: 1-6%). At the base and the top, erosional unconformities connect these deposits with the Carcarañá Formation and with the San Guillermo Formation respectively (Kröhling 1999). A primary loess facies or aeolian facies of the Tezanos Pinto Formation has the greatest areal representation, with a typical outcropping thickness of 2-4 m, and more developed on the interfluves (6-8 m thick). It is a loose deposit, coarse silt with subordinate clay and fine sand, light brown in colour (7.5 YR 6/4). It is a homogeneous, porous and permeable deposit with a coarse granular to mediumcoarse blocky fabric. The sedimentary mass is calcareous; it contains powdery concentrations and hard concretions of CaCO3 (in a variable frequency; varied forms and centimetric sizes). The loess body is crossed by fine rhizoid ramified canalicula. It is stable in steep walls, in parts altered by subcutaneous subfusion and shaped by columnar disjunction. TL datings gave ages of 31.69 ± 1.62 ka BP and 32.0 ka BP in samples of the aeolian facies of the formation in the area, at 4 m below the top of the unit (LGM, OIS 2). Buried soil (Kröhling 1999) The top of Tezanos Pinto Formation is marked by a partially eroded soil, 0.30 -1.10 m thick, typically represented by a Bt horizon. In a minor geomorphological unit (fluvial valleys of the region) that soil forms an accretionary pedocomplex of argillic horizons separated by the accumulation of Andean volcanic ash, mainly concentrated by alluvial processes. At the interfluves of the region, the Bt-horizon is formed by a dark brown (7.5 YR 3/4) clayish silt, with intense illuviation and formation of cutans. It has a strongly to moderately developed structure in medium angular prisms, strong consistence, with very fine and fine cracks between peds. It lacks nodules or mottles, and the silt fraction is non-calcareous. There are abundant root moulds. The lower boundary is wavy in form and well marked. The Bt horizon is composed of a dull brown (7.5 YR 5/4) slightly clayish coarse silt. This horizon shows a weakly to moderately well-developed structure, formed by medium blocks, very firm, limited by fine and very fine fissures. Nodules or mottles are absent, but fine macropores are present. The silt fraction is non-calcareous. The C horizon shows characteristics transitional to the underlying loess. The buried soil was generated during the Holocene Optimum Climaticum and it is a distinct pedostratigraphic marker in different areas of the Pampas plain. San Guillermo Formation (Iriondo 1987) A younger loessic formation overlies the buried soil on top of the Tezanos Pinto Formation, separated from it by an erosional unconformity. It is composed of a brownish grey (10 YR 5/1) coarse silt with scarce proportions of very fine sand and clay. The unit is massive, friable in general, porous, permeable, and moderately structured in very coarse firm prisms bounded by very fine fissures. This deposit lacks nodules or mottles, but contains numerous root moulds, abundant macropores, very fine and fine canalicula and tubes generated by bioturbation. The sediment mass is non-calcareous. The unit is partly the product of deflation of the A horizon of the underlying soil and the subsequent deposition of dust. The typical thickness is 0.30 m, with a maximum of 0.55 m. The San Guillermo Formation generally forms the top of the sedimentary sequence in the natural sections of the Pampas. It was deposited during a Late Holocene dry period that occurred between 3.5 and 1.4 ka BP (Iriondo 1990). Lucio Lopez Formation (Kröhling 1996) It constitutes a complex sequence differentiated into three sectors. It includes a clayish silt with scarce very fine sand (silt: 58-77%; clay: 19-37%; sand: 3-6%), organized in fine strata with variations in colour from olive to grey; pedogenic horizons are intercalated in the middle section (pedocomplex). Locally, it exhibits high proportions of biogenic material (mainly gastropods, ostracods, diatoms and abundant plant remains). The Lucio Lopez Formation outcrops along the cliffs of the main rivers of the northern Pampas region, with most typical characteristics and the greatest thickness on the Río Carcarañá cliffs. The thickness varies from 0.50 to 5.00 m. The formation unconformably overlies the Carcarañá Formation in sections located inside the fluvial valleys of the region. The contact indicates a palaeotopography of channels and gullies. In general the formation is covered by contemporary or recent deposits of fluvial origin; locally the unit forms the top of the section. The formation, with a clear paludal origin, constitutes the infilling of erosional landforms located in a recent belt of the main rivers and at the bottom of the major fluvial palaeovalleys of the region. A TL date in the upper section of the formation (a cineritic stratum) indicates an age of 1.32 ± 0.12 ka BP. The soil complex of the middle section was generated during the Holocene Optimum Climaticum. The formation was generated during the Holocene and possibly even the Late Pleistocene too. This unit correlates with the Luján Formation defined by Fidalgo et al. (1973), representing the Late Quaternary continental infilling of the fluvial valleys of NE Buenos Aires province. Toledo (2005) presented a sequence stratigraphic model in the Lujan type section of this unit, based on detailed stratigraphic observations and C14 datings on mollusks. The author refered ages from > 40 ka BP to >11 ka for the Guerrero Member (fluvial-paludal deposits or Lujanense) and ages from 10.7 to 3.3 ka BP for the Río Salado Member of the Luján Formation (grayish to whitish silty facies or Platense). Prieto et al. (2004) gave ages ranging from 11 to 3.5 ka BP for the Luján Formation in the Río Luján.
LA BAJADA SECTION
The town La Bajada is located across the Río Paraná from Santa Fe. At present, that locality (nowadays Paraná) is the capital city of Entre Ríos province. Such area is crossed by an old transcurrent fracture, the Tostado-Gualeguaychú fault, which elevated its southern block some 40 m in the Middle Pleistocene (Iriondo 1989), thus exposing Tertiary marine strata. Darwin payed special attention to those strata, collected an interesting fossil assemblage and made an excellent stratigraphic description of the section. Darwin quoted that "In Entre Ríos, the cliffs, estimated at between sixty and seventy feet in height, expose an interesting section: the lower half consists of Tertiary strata with marine shells, and the upper half of the Pampean formation. The lowest bed is obliquely laminated, blackish, indurated mud, with distinct traces of vegetable remains. Above this there is a thick bed of yellowish sandy clay, with much crystallized gypsum and many shells of Ostreae, Pectens, and Arcae: above this generally comes an arenaceous crystalline limestone, but there is sometimes interposed a bed, about twelve feet thick, of dark green, soapy clay, weathering into small angular fragments. The limestone, where purest, is white, highly crystalline, and full of cavities: it includes small pebbles of quartz, broken shells, teeth of sharks, and sometimes, as I was informed, large bones: it often contains so much sand as to pass into a calcareous sandstone, and in such parts the great Ostrea patagonica chiefly abounds. In the upper part, the limestone alternates with layers of fine white sand. The shells included in these beds have been named for me by M. d'Orbigny (a list of fossils follows) - M. d'Orbigny has given a detailed description of this section, but as he does not mention this lowest bed, it may have been concealed when he was there by the river. There is a considerable discrepancy between his description and mine, which I can only account for by the beds themselves varying considerably in short distances" (Darwin 1846, p. 88-89). In fact, the major discrepancies between both naturalists appeared because Darwin worked in the elevated block of the fault and d'Orbigny researched in the downthrown block, where the marine beds appear only during low waters. Besides, as Darwin correctly stated, the strata vary in short distances. However, Darwin's description fits considerably well with the type section of the Paraná Formation, formally defined there 150 years later by Iriondo (1973). The marine beds of Bajada were renamed as Paraná Formation in the 20th Century. This formation represents the last widespread marine transgression occurring in the interior of South America (Yrigoyen 1969, Herbst 1971, Iriondo 1973, Marengo 2005) and the Bajada-Paraná area is considered the type locality. Darwin's section, indeed, can clearly be recognized in the type section of the formation (Iriondo 1973, Fig. 4):
- Both sections are composed of five beds and have similar thicknesses; a comparison shows the following: - The lowest bed - Darwin (profile B of Fig. 4): Obliquely laminated, blackish, indurated mud, with distinct traces of plant remains. Type section: 3 m. Green quartz sand in 30-50 cm thick strata, including a large proportion of clay intraclasts, most of them are platy angular fragments without rounding. Major axes of plates are parallel to stratification planes. Mean grain size around 0.2 mm, finer towards the bottom. Transitional upper contact. - Second bed - Darwin: Yellowish sandy clay, with much arenaceous crystalline gypsum and Ostreae. Type section: 3 m. Massive green sand, without internal bedding, containing up to 10 cm long elliptic clay intraclasts. 20% of the sediment mass is composed of chaotically distributed small platy intraclasts. Grain size around 0.25 mm with variable dispersion, which grows upwards. Lightly cemented with CaCO3. The detachment and fall of intraclasts produces numerous small hollows in outcrops. Neat upward contact. - Third bed - Darwin: 4 m. Dark green, soapy clay weathering into small angular fragments. Type section: 6 m. Sequence of interestratified sand and clayish silt. The sand bodies are short lenses with internal diagonal lamination, up to 1.60 m thick. Loose, quartzose and yellow in colour. Sandy bodies are generally composed of 5-15 cm thick internal units with diagonal stratification. The fine sediments are plastic, gray clayey silts, forming continuous strata 1 to 30 cm thick and at least 50-60 m long; internal laminae of sand are common. In parts with contorted internal lamination. Silt strata conformably cover the underlying sand lenses. Grain size of the fines is 30 microns at the bottom, diminishing upwards to clay-sizes. The upper section of this bed changes to white sandstone with dune structures containing well preserved bivalves. Further on, the bed passes to a fine conglomerate. - Fourth bed - Darwin: Sandy crystalline limestone. Where purest, it is white, highly crystalline, and full of cavities: it includes small pebbles of quartz, broken shells, teeth of sharks and sometimes large bones: it often contains so much sand as to pass into a calcareous sandstone, and in such parts the great Ostrea patagonica abounds (Darwin 1846, p. 89). Type section: 1.5 m. White calcareous sandstone with dune structures and undulose strata. Lateral and upward enrichment with shells of bivalves and scarce oysters. Bivalve shells underwent an advanced degree of dissolution and re-precipitation, remaining often only as moulds. Oysters are well preserved. - Fifth (upper) bed - Darwin: The limestone alternates with layers of fine white sand. Type section: 5.5 m. Most of the outcrop is covered by debris. The lower sector is characterized by white sand, incipiently cemented by the carbonate provided by the local dissolution of shells; the upper section is composed of fine quartz sand. Green in colour.
 Figure 4: The Paraná Formation outcropping in La Bajada. A: Type profile (Iriondo 1973). B: Darwin's profile.
Figure 4: The Paraná Formation outcropping in La Bajada. A: Type profile (Iriondo 1973). B: Darwin's profile.
The general scenario suggested by the section in both approaches (Darwin's and modern) is one of a tidal environment, with subtidal sands and gravels, intertidal muds, changing tidal channels and coastal currents. Two different sedimentary mechanisms can be deduced: a) Transport and accumulation of sand by tractive currents, probably from nearby beaches if one considers the negative skewness of the sediment. b) Flocculation of fines in calm environments. The fossil assemblage also points to a littoral/neritic position in a warm climate. A general setting of the Paraná Formation is as follows: The Paraná Formation covers an important sector of a major geological event occurring in the Neogene of South America. Around the Middle Miocene occurred a generalized marine transgression in large regions of the continent. The Paraná Formation comprises the portion included in the Chaco-Paranense Basin, which extends from the central lowlands of Argentina at 36° lat. S northwards to the Bolivian and Paraguayan Chaco. The Miocene sea was a shallow, platform branch of the Atlantic Ocean. Mean thickness of the formation is about 100 m; the sediment is dominantly sandy in the type area (Paraná; La Bajada in Darwin times) and characterizes the upper half of the section in all the basin. The lower half appears as composed of green sand. A research borehole drilled in the center of the basin in the northern Pampas (San Guillermo locality; 30°15'S, 61°50'W) recorded 40.5 m of the Paraná Formation covered by 60.3 m of Quaternary loessic formations (Kröhling and Iriondo 2003). Following Darwin, "The upper half of the cliff in La Bajada, to a thickness of about thirty feet, consists of Pampean mud, of which the lower part is pale-coloured, and the upper part of a brighter red… Close above the marine limestone, there is a thin stratum with a concretionary outline of white hard tosca-rock or marl…" Three Quaternary formations can be recognized in that reference: The upper one is Tezanos Pinto Formation, the main loessic deposit of the Last Glacial Maximum. The "lower mud" is the Hernandarias Formation, an Early Pleistocene playa deposit of the Río Uruguay. The tosca-rock is a well developed level, which thickens southwards up to 9 m and is known as Puerto Alvear Formation.
SOUTHWESTERN ENTRE RÍOS
A key area of the region is located in the southwestern part of Entre Ríos province and outcrops along the eastern cliff of the Paraná flood plain. Darwin did not study such outcrops, but navigated along them in his journey back to Buenos Aires. However, he made a few comments on "the Punta Gorda in Entre Ríos" (different from the other Punta Gorda in the Banda Oriental), which is an interesting sedimentary cycle that occurred around one million years before present. That locality is placed 60 km south of Santa Fe and has the best stratigraphic section of the area (Fig. 5) in Entre Ríos.

 Figure 5: Punta Gorda section, representative of SW Entre Ríos province (after Iriondo and Kröhling 2008).
Figure 5: Punta Gorda section, representative of SW Entre Ríos province (after Iriondo and Kröhling 2008).
Punta Gorda Section
The Quaternary geologic column of the region (Kröhling 2001) records two well defined aeolian sedimentation cycles. In both cases, the main source of sediment has been the Andean region, that produced fine materials originated by nival processes and volcanic eruptions. The older cycle, defined as the Punta Gorda Group, is composed of three units accumulated in paludal and subaereal environments during the Early Pleistocene. The second sedimentary cycle occurred during the Late Pleistocene and Holocene (basically at the Last Glacial Maximum) and is the Pampean Aeolian System traversed by Darwin from Buenos Aires to Santa Fe. It comprises two formations accumulated in subaereal environments. A short period of aeolian remobilization occurred during the Late Holocene. Several episodes of pedogenesis and local erosional unconformities were recorded in both major sedimentation periods, particularly in the older one. The long sedimentation hiatus, covering all the Middle Pleistocene and a half of the Late Pleistocene constitutes a remarkable regional feature. Important similarities were found between both sedimentary cycles: i) the same source of sediments; ii) the same long distance transport agent (wind); iii) striking similarity in grain-size distributions; iv) scarce contribution of materials from the Brazilian shield and associated areas in the north. The age of the older cycle can be located between the Brunhes - Matuyama magnetic polarity change (the Upper Matuyama Chron > 0.78 Ma.) and the Jaramillo Subchron (1 Ma). This conclusion coincides with recent datings in the "Great Patagonic Glaciation" (Ton-That et al. 1999), which rendered ages from 1 Ma to 1.17 Ma. The following sedimentary units outcrop at Punta Gorda: Punta Gorda Group (Iriondo 1980): Sedimentary cycle 1 Puerto Alvear Formation (Iriondo 1980) This is a sedimentary body accumulated as infilling of an abandoned belt of the Río Paraná. The unit was a non-permanent swamp at the beginning of the Pleistocene. The formation lies unconformably on the Paraná Formation (marine Miocene), with a maximum thickness of 9 m. The most visible field feature is a closed net formed by CaCO3 plates of phreatic origin, 0.5 to 4 cm thick with a dominant horizontal development. Numerous vertical large and botrioidal concretions indicate a post-depositional remobilization of the carbonate. The clastic component of the unit is a silty clay to sandy silt with diffuse lamination, light reddish brown in colour with olive patches. Abundant Mn- and Fe segregations and nodules are conspicuous. This formation is divided into two members; the lower one is characterized by thick continuous partition walls, with a wavy trace. The upper member includes a similar horizontal carbonate net, although formed by thinner and more irregular platy precipitates. An internal unconformity separates both members; in some places the top of the lower member are the Bw-and C horizons of a weakly developed paleosol. That pedogenic level is non-calcareous. La Juanita Formation (Iriondo 1998) This unit was formed in a paludal environment during the Early Pleistocene. It lays unconformably on the Puerto Alvear Formation, with a typical thickness of 2 to 3 m. The La Juanita Formation is composed of light brown to olive silt with scarce fine sand with horizontal diffuse bedding. Strata are 15 to 30 cm thick, with internal lamination. Fe- and Mn segregations and root moulds are locally abundant. The sediment is slightly structured in firm blocks. Concretions, which form more than 10% of the sedimentary mass, are rough vertical infillings of roots, 2 to 5 cm thick and up to 20 cm long. The general tendency of the CaCO3 precipitates indicates infiltration in a non-permanent swamp environment with a non-saturated bottom. There are also infillings of horizontal fissures. Frequent Mn films cover the surface of concretions, indicating a younger age for the mobilization of such oxides. Punta Gorda Formation (Iriondo and Kröhling 2008) This is a brown loess characterized by pedogenesis in several levels and frequent local internal unconformities. According to palaeomagnetic analyses, the age of the unit is Early Pleistocene. Carbonatic cement is widespread in the formation. Local second order variations are the rule in most outcrops. Three sections were described in the unit at the type locality (Punta Gorda, Entre Ríos province). The lower one was unconformably deposited over La Juanita Formation and begins with a paludal deposit composed of clayish silt, light brown in colour, with Fe- and Mn segregations. This is conformably overlain by a petrocalcic horizon formed by vertical cylindrical welded concretions 50 cm high. The central section is formed by 4 m of homogeneuos light brown loess, with a vertical slope. A lattice pattern of CaCO3 precipitates appears in the exposures. Three weak to moderate paleosols (Band Ck horizons) were recorded at the typical site. These soils are laterally discontinuous as a result of uneven erosional contact with the overlying loess. They are re-calcified from this material too. The upper section begins with a Ckm horizon of another eroded paleosol, covered by a one meter thick paludal deposit. Tezanos Pinto Formation (Iriondo 1980): Sedimentary cycle 2 The unit constitutes the Peripheral Loess Belt of the Pampean Aeolian System, which was deposited during the OIS 2 (36-8 ka BP, Iriondo and Kröhling 1995). The aeolian dust was transported from Andean sources by S and SW winds. In SW Entre Ríos province the unit is yellowish brown loess, 2 to 3 m thick, composed of powderish sandy silt, massive and friable. OSL ages of 32 and 24 ka BP were obtained for the loess in this region. The loess forms typical vertical slopes, with columnar disjunction. It contains small CaCO3 concretions and frequent rhizoconcretions (15.48 ± 0.19 C14 ka BP).The unit received contributions of fine sediments from the Paraná headwaters, located to the northeast. The clay fraction of the loess is a complex mixture of illite, interstratified illitesmectites and kaolinite. Illite is a pampean mineral whereas smectites and kaolinite are derived from Brazil. A partially eroded soil on top of the loess is represented by well developed Bt-horizon and C horizons. The Bt horizon is 35 to 55 cm thick, clayey silt dark brown in colour, structured in very firm prisms; argillocutans are frequent. The buried soil was developed under humid subtropical conditions during the Holocene Optimum Climaticum. San Guillermo Formation (Iriondo 1980): Sedimentary cycle 2 The sedimentary sequence of Punta Gorda ends with an aeolian unit unconformably accumulated over the eroded soil. It is a brownish gray loess, 20 to 35 cm thick, loamy to silty-loamy. This unit is the result of deflation of the A-horizon of the soil on top of the Tezanos Pinto Loess and the subsequent deposition of dust. Such an event occurred from 3.5 to 1.4 ka BP.
THE PARANÁ FLOOD PLAIN
Darwin travelled from Santa Fe back to Buenos Aires by boat along the Paraná and made some remarks about the river, its banks and fauna. Particularly, he noted the large number of islands that are permanently eroded and reconstructed by the currents. In fact, according to modern geomorphology the river flows there within a large flood plain, approximately 600 km long, stretching from the Paraguay-Paraná junction (27°25'S) to south of Rosario (33°S). Downstream it passes transitionally to a well-developed Holocene littoral complex. The general direction of the flood plain is north-south, crossing several tectonic blocks in a perpendicular or diagonal manner. This relationship has generated local segments within the plain, with slightly different directions and slopes. The width of the floodplain varies from 13 to 45 km, with typical values between 25 and 35 km. It is composed of several internal geomorphological units, the most modern of them is the "bar plain", that is developing at present (Iriondo 2007). The bar plain is a belt of large elliptic bars composed of very fine sand, that encloses the main channel along the whole flood plain. This belt is permanently modified by erosion and sedimentation and has a width of 2-7 km; it is formed by islands inside the main (navigation) channel and sand bars juxtaposed to the margins. The large bars are crossed by minor active channels, most of them with intense lateral migration, resulting in minor meander belts. The powerful braided mainstream - with a mean discharge between 16,000 and 20,000 m3/sec - continuously reshapes islands and banks. This is the environment described by Darwin. Other important geomorphological units of the flood plain are the hindered drainage plain and the deltas of tributaries. The hindered drainage plain was described by Darwin as a labyrinth of small branches separated by low islands covered by forest, he observed this landscape when crossing the flood plain from Santa Fe to La Bajada. This unit is characterized by a morphology of flat areas with numerous shallow ponds, swamps and small adventitious channels. Such channels are tortuous, active only at the beginning and end of floods, conveying water into ponds during rising and back to the main channel at low levels. The number of ponds and shallow lakes is huge - about 5,000 according to Paira and Drago (2007). This system was generated under a hydric regime different than the present one. The deltas of tributaries are areas of sedimentation formed inside the flood plain during a recent dry climate period (from 3.5 to 1.4 ka BP, according to the latest datings). Water discharges were lower than today in all the system and tributaries did not reach the collector, depositing the sediment loads in lateral positions inside the flood plain; the Paraná River itself was also smaller then than it is today. That resulted in relatively short and wide delta-like sedimentary bodies. Most of such deltas (Los Amores, del Rey, Corrientes) are located north of Santa Fe, a region not visited by Darwin owing to health problems. Terraces also appear in the north, in Chaco and Corrientes provinces. The sediments of the flood plain are very homogeneous. Practically all the sedimentary mass is composed of only three sedimentary facies: a) Channel facies, which are composed of fine and very fine quartz sand, generally in 10-40 cm thick horizontal strata with diagonal internal structures. Such sand was originated in Mesozoic sandstones of the upper basin and comprises the major part of the buried sediments. b) Levee facies, which are characterized by silty-clayish very fine sand with intermediate plasticity; ochre to greenish-gray in color, mottled, with numerous pores, tubes and root moulds. Gley processes are frequent. c) Lake-and-swamp facies, composed of dark gray silt with abundant organic matter in different degrees of decomposition. It is compact, with low porosity; sand and clay are scarce. A really modest proportion of the sediment volume is represented by a fourth facies, i.e., aeolian dunes composed of very fine sand, which appear in isolated spots in the northern half of the flood plain.
THE LITTORAL COMPLEX AT THE RÍO PARANÁ MOUTH
Darwin noted that "Some leagues downstream Rosario begins, at the western margin of the Paraná, a line of perpendicular cliffs that extends beneath San Nicolás…The banks of the river are formed by very soft earths; in consequence the waters are muddy…". The author has entered the Paraná Delta, which in fact is a large littoral complex of Holocene age formed by several geomorphological and sedimentary units sequentially generated. Four main phases have been identified there by Iriondo (2004, Fig. 6): 1) A fluvial period represented by river flood deposits; 2) A marine ingression with development of a sand barrier, a lagoon, minor tributary deltas and estuaries and well-developed regression deposits; 3) An estuarine phase characterized by extensive tidal deposits in the central area; and 4) The present fluvial period, with channel deposits and a deltaic unit advancing into the Río de la Plata. The present dynamics of the deltaic complex is dominated by the Paraná floods, the floods of the Uruguay and Gualeguay rivers, Atlantic tides, and floods produced by the southeasterly winds. The southeasterly wind (Sudestada), indeed, is a major factor in the water dynamics and navigation in that area and downstream in the Río de la Plata. The wind can be strong and blow sometimes during several days, provoking a rise of the water level. Records of up to 2.5 m above the terrain have been reported at several places. During such periods, navigation in boats and small vessels stops; surely, Darwin underwent such a meteorological phenomenon: "At night, the wind is scantly favorable and we stop; the next day blows a strong wind…"
Figure 6: Map of the Holocene littoral complex at the Paraná mouth (after Iriondo 2004).
COMMENTS AND CONCLUSIONS
Undoubtedly, the short incursion of the great naturalist to the interior of the Argentine plains was a very modest portion of his famous trip around the world, with plenty of discoveries, experiences and amazing landscapes such as Tahiti or Tierra del Fuego. The attention paid by Darwin to these flat plains in his subsequent studies was necessarily far more modest than those devoted to the Galapagos Islands or the Andes Cordillera. However, the few pages dedicated to the Buenos Aires-Santa Fe journey reveal the extraordinary abilities of Darwin for keen observation and logical thinking, in spite of the limitations of geological theory at that time. No real errors can be noted in his geological observations recorded during this journey, rather good science and acute synthetic descriptions. In order to make a comparison between the geological contributions of Darwin and the present knowledge on the region, the authors of this contribution necessarily follow the general structure of Darwin's books, a traveler's diary enriched by digressions on Science, Politics and History.
WORKS CITED IN THE TEXT
1. De Alba, E. 1953. Geología del Alto Paraná - Relación con los trabajos de derrocamiento entre Ituzaingó y Posadas. Revista de la Asociación Geológica Argentina 8:129-161.
2. Darwin, C. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 280 p., London.
3. Darwin, C. 1945. Viaje de un naturalista alrededor del Mundo. Librería El Ateneo, 118 p. (Spanish translation of the second English edition), Buenos Aires.
4. Fidalgo, F., De Francesco F. and Colado, U. (1973). Geología superficial de las Hojas Castelli, J.M. Cobo y Monasterio (provincia de Buenos Aires). 5° Congreso Geológico Argentino (Córdoba), Actas 4: 27-39.
5. Georgieff, S., Anis, K., Orfeo, O., and Rizo, G. 2005. Architecture of Ituzaingo Formation (Pliocene), a comparison with deposits of Paraná River, Argentina. University of Technology, 8th. International Conference on Fluvial Sedimentology, Abstracts: 107, Delft.
6. González Bonorino, F. 1965. Mineralogía de las fracciones arcilla y limo del Pampeano en el área de la ciudad de Buenos Aires y su significación estratigráfica y sedimentológica. Revista de la Asociación Geológica Argentina 20: 67-148.
7. Herbst, R. 1971. Esquema estratigráfico de la provincia de Corrientes. Revista de la Asociación Geológica Argentina 26: 221-243.
8. Imbellone, P.E. and Cumba, A. 2003. Una sucesión con paleosuelos superpuestos del Pleistoceno medio tardío-Holoceno, zona sur de la Plata, provincia de Buenos Aires. Revista de la Asociación Argentina de Sedimentología 10: 3-22, La Plata.
9. Iriondo, M. 1973. Análisis ambiental de la Formación Paraná en su área tipo. Boletín de la Asociación Geológica de Córdoba 2(1-2): 19-23.
10. Iriondo, M. 1980. El Cuaternario de Entre Ríos. Revista de la Asociación de Ciencias Naturales del Litoral 11: 125-141, Santo Tomé.
11. Iriondo, M. l987. Geomorfología y Cuaternario de la Provincia de Santa Fe. D'Orbignyana 4: 1-54, Corrientes.
12. Iriondo, M. 1989. Major fractures of the Chaco-Pampa plain. Bulletin of the INQUA Neotectonic Commission 12: 42-46, Stockholm.
13. Iriondo, M. 1990. A Late Holocene dry period in the Argentine plains. Quaternary of South America and Antarctic Peninsula 7: 197-218.
14. Iriondo, M. 1998. Excursion Guide Nº3: Province of Entre Rios. International Joint Field Meeting: Loess in Argentina: Temperate and Tropical. INQUA - PASH - CLIP - UNER - CECOAL 3: 1-19, Paraná.
15. Iriondo, M. 2004. The littoral complex at the Paraná mouth. Quaternary International 114: 143-154.
16. Iriondo, M. 2007. Geomorphology and Quaternary of the Flood-Plain. In Iriondo, M., Paggi, J. and Parma J. (eds.) Limnology of the Middle Paraná River: A Subtropical Wetland, Springer Verlag, 382 p., Berlin, Heidelberg, London.
17. Iriondo, M. and Kröhling, D. 1995. El Sistema Eólico Pampeano. Comunicaciones del Museo Provincial de Ciencias Naturales "Florentino Ameghino" (NS) 5(1): 1-68, Santa Fe.
18. Iriondo, M. and Kröhling, D. 2008. Cambios ambientales en la cuenca del Uruguay (desde el Presente hasta dos millones de años atrás). Colección Ciencia y Técnica, Ediciones UNL 358 p., Santa Fe.
19. Kemp, R., Zárate, M, Toms, P. King, M., Sanabria, J. and Arguello, G. 2006. Late Quaternary paleosols, stratigraphy and landscape evolution in the Northern Pampa, Argentina. Quaternary Research 66: 119-132.
20. Kröhling, D. 1996. La Formación Lucio Lopez (Holoceno), noreste de la llanura pampeana. 13° Congreso Geológico Argentino (Buenos Aires), Actas 4: 69.
21. Kröhling, D. 1998. Geomorfología y Geología del Cuaternario de la cuenca del río Carcarañá, desde la confluencia de los ríos Tercero y Cuarto, provincias de Santa Fe y Córdoba. Doctoral Thesis, Universidad Nacional de Córdoba (unpublished), 224 p., Córdoba.
22. Kröhling, D. 1999. Upper Quaternary of the Lower Carcarañá Basin, North Pampa, Argentina. In Partridge, T., Kershaw, P. and Iriondo, M. (eds.) Paleoclimates of the Southern Hemisphere, Quaternary International 57/58: 135-148, Oxford.
23. Kröhling, D. 2001. Quaternary paleosol: loessic sequences of southwestern Entre Ríos Province, Northern Pampa, Argentina. 6° International Symposium and Field Workshop on Paleopedology in Mexico City (INQUA - ISSS), Proceedings: 27-28, México.
24. Kröhling, D. and Iriondo, M. 2003. El loess de la Pampa Norte en el Bloque de San Guillermo. Revista de la Asociación Argentina de Sedimentología 10(2): 137-150, La Plata.
25. Marengo, H. 2005. Micropaleontología y Estratigrafía del Mioceno Marino de la Argentina: las transgresiones de Laguna Paiva y del ¨Entrerriense- Paranense¨. Doctoral Thesis, Universidad de Buenos Aires (unpublished), 150 p., Buenos Aires.
26. Mercer, J. 1976. Glacial history of Southernmost South America. Quaternary Research 6: 125- 166.
27. Nabel, P.E., Camilión, M.C., Machado, G.A., Spiegelman, A. and Mormeneo, L. 1993. Magneto y litoestrafigrafía de los sedimentos pampeanos en los alrededores de la ciudad de Baradero, Provincia de Buenos Aires. Revista de la Asociación Geológica Argentina 48: 193-206.
28. Paira, A. and Drago, E. 2007. Origin, evolution and types of flood plain water bodies. In Iriondo, M., Paggi, J. and Parma, M. (eds.) The Middle Paraná: Limnology of a Subtropical Wetland, Springer 3: 53-82, Heidelberg.
29. Prieto, A., Blassi, A., De Francesco, C. and Fernández, C. 2004. Environmental history since 11,000 C14 yr. BP of the northeastern Pampas, Argentina, from alluvial sequences of the Luján River. Quaternary Research 62: 146-161.
30. Parra, P. 1939. Diario y derrotero de sus viajes. Editorial Solar, 251 p., Buenos Aires.
31. Riggi, J., Fidalgo, F., Martinez, O. and Porro, N. 1986. Geología de los "Sedimentos Pampeanos" en el Partido de La Plata. Revista de la Asociación Geológica Argentina 41: 316-333.
32. Toledo, M. 2005. Secuencias pleistocenas "lujanenses" en su sección tipo: Primeras dataciones C14 e implicancias estratigráficas, arqueológicas e históricas, Luján-Jáuregui, provincia de Buenos Aires. Revista de la Asociación Geológica Argentina, Comunicaciones 60(2): 417-424.
33. Tonni, E.P., Nabel, P., Cione, A.L., Etchichury, M., Tófalo, R., Scillato Yané, G., San Cristóbal, J., Carlini, A. and Vargas, D. 1999. The Ensenada and Buenos Aires formations (Pleistocene) in a quarry near La Plata, Argentina. Journal of South American Earth Sciences 12: 273-291.
34. Ton-That, T., Singer, B., Mörner, N. and Rabassa, J. 1999. Datación de lavas basálticas por Ar/ Ar y geología glacial de la región del lago Buenos Aires, prov. de Santa Cruz, Argentina. Revista de la Asociación Geológica Argentina 54: 333-352.
35. Yrigoyen, M. 1969. Problemas estratigráficos del Terciario de Argentina. Ameghiniana 6: 315- 328.
On the formations of the Pampas in the footsteps of Darwin: south of the Salado
Marcelo Zárate1 and Alicia Folguera2
1 CONICET- Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Santa Rosa, La Pampa. E-mail: marcelozarate55@yahoo.com.ar 2 Servicio Geológico Minero Argentino (SEGEMAR), Buenos Aires. Email: alifolguera@yahoo.com
ABSTRACT
In 1833 during his journey across the Buenos Aires Pampas, Charles Darwin made observations that reflected his thoughts on two major landscape units, Pampa interserrana and Pampa deprimida, later identified by other authors. Darwin grouped the Pampean sediments into a single unit, the Pampean Formation, based upon the lithological homogeneity and the large extension of the deposits; the unit was thought to be of estuarine-marine origin and attributed to the Recent Epoch considering the paleontological content (vertebrates and mollusks). At present, the Pampean sedimentary succession, which accumulated approximately during the last 11-12 Ma, is interpreted as a pedosedimentary sequence due to the ubiquity of pedogenetic features throughout the deposits. Four main subcycles of sedimentation are identified related to reactivations of the Pampean landscape. At a regional scale, the outcrop distribution of Pampean sediments of different ages suggests the dominance of more stable conditions since the late Miocene-Pliocene in a vast area of Pampa interserrana, documented by the formation of calcretes. However, sedimentation during the late Pliocene-Pleistocene was active within the domain of the Salado tectonic basin and Sierras de Tandil. The regional disparity shown by the Pampean stratigraphic record reveals the major morphostructural differences of its basement.
Keywords: Charles Darwin; Pampean sediments; Calcrete; Geomorphology; Buenos Aires.
RESUMEN: Sobre las formaciones de las Pampas en los pasos de Darwin: al sur del Salado. Durante su viaje por la Pampa bonaerense en 1833 Charles Darwin efectuó observaciones que reflejaban las dos grandes unidades de paisaje posteriormente reconocidas en la región, la Pampa interserrana y la Pampa deprimida. La homogeneidad litológica y la vasta extensión de los depósitos fueron los criterios básicos empleados para agruparlos en una única unidad, la Formación Pampeano; basado en criterios paleontológicos le atribuyó origen estuárico-marino y la asignó a la época Reciente. Si bien han existido otras propuestas estratigráficas, Formación Pampeano o simplemente Pampeano, son denominaciones empleadas informalmente y con un significado algo vago. Actualmente, sobre la base de la ubicuidad de los rasgos pedológicos de la sucesión sedimentaria se la interpreta como una secuencia pedosedimentaria acumulada durante los últimos 11-12 millones de años, se han reconocido 4 ciclos de sedimentación principales relacionados con reactivaciones del paisaje pampeano. Regionalmente, la distribución areal de niveles aflorantes de distintas edades sugiere una mayor estabilidad relativa en el sur, marcada por la formación de calcretos en la Pampa interserrana a partir del Mioceno tardío-Plioceno y mayor continuidad del proceso sedimentario en el ámbito de la cuenca tectónica del Salado y de las sierras de Tandil. Las variaciones regionales exhibidas por el registro estratigráfico del Cenozoico tardío constituyen una manifestación del comportamiento diferencial de las unidades morfoestructurales que integran el basamento de la sucesión sedimentaria.
Palabras clave: Charles Darwin; Sedimentos pampeanos; Tosca; Geomorfología; Buenos Aires.
INTRODUCTION
In the course of the winter of 1833, Charles Darwin travelled across the Buenos Aires Pampas. His observations, as well as those by Alcide d'Orbigny (1842), the French naturalist who explored the region some years before Darwin, were part of the initial expansion stage of geology as a science. The explorations of both naturalists are framed within the creation of a universal scheme of the Earth history in which all the regions of the world would be incorporated (Podgorny et al. 2008). In early August of 1833, Darwin disembarked from the Beagle at Patagones, a town by the Rio Negro, in northern Patagonia and began his journey heading north towards Bahía Blanca. From there he travelled across the southern Pampas to the Rio Salado area and Buenos Aires, continuing later to Santa Fe. Logistics for the trip was provided by Juan Manuel de Rosas - the Argentine administrator at the time - in the form of a passport to travel, horses, and a group of soldiers and gauchos to escort him. In his diary, Darwin took notes on the Pampean scenery, referring to the dominant vegetation, the available water sources, the topographic features and the sediments and rocks he found along the journey. He also provided a historical picture of Argentina with descriptions and his own viewpoints on the people and the ongoing war with the aboriginal groups to expand the frontier of the southern territories westward. The information gathered during the field trip was the raw material to write On the Formations of the Pampas, the fourth chapter of his Geological Observations on South America (1846). The vast extent, the disputed origin and the abundance of fossil mammal remains were the three main aspects underlined by Charles Darwin when he summarized the characteristics of the Pampean sediments. What was the meaning of the Pampean Formation as used by Darwin?; what is the meaning of this term at present?; what is our present understanding of the late Cenozoic Pampean geology?. This paper is particularly focused on the Buenos Aires Pampas, south of the Rio Salado, the region that Darwin explored during the winter of 1833; it includes comments on specific observations he made elsewhere to understand better his interpretations and inferences. Consequently, the main goal is to analyze and discuss his observations on the landscape and the sediments within the framework of our present understanding of the late Cenozoic geology of the southern Pampean plain. With this purpose in mind, the present contribution brings in Darwin 's comments and interpretations on the landscape along with the nature, composition and areal distribution of the Pampean sediments (terms, descriptions, and special comments are quoted in italics and referred to his diary and the chapter on the Formations of the Pampas). These particular issues are then analyzed from our present geological perspective, providing an updated geological review.
DARWIN'S ITINERARY AND HIS PERCEPTION OF THE PAMPEAN LANDSCAPE
Charles Darwin travelled around 650 km (400 geographical miles) from Rio Colorado to Buenos Aires following the road that connected the military outposts (Postas) set up along the western frontier in 1833 during the ongoing war with the aboriginal groups (Fig. 1). On the way to Bahía Blanca, once he crossed the Río Colorado, Darwin noticed a major change between the Patagonian and the Pampean environments "...The country had a different appearance from that South of the Colorado: there were many different plants and grasses and not nearly so many spiny bushes and these gradually became less frequent; until a little to the North there is not a bush..."(Darwin's Beagle Diary 1831-1836). In southern Buenos Aires province he examined the neighboring areas of Bahia Blanca and Punta Alta where he spent around a week. On August 27th Darwin continued to the next military outpost, "Sauce Posta", situated by the Rio Sauce Grande, "…the distance was about 6 leagues from Sierra de la Ventana…" (Darwin's Beagle Diary 1831-1836). It is likely that this Posta was situated in the vicinity of the present village of Saldungaray.
 Figure 1: Darwin's itinerary across the Pampean plain of Buenos Aires province. S: Sauce Posta, T: Tapalqué, G: Guardia del Monte; 1833 frontier adapted from Rolleri et al. (2005)
Figure 1: Darwin's itinerary across the Pampean plain of Buenos Aires province. S: Sauce Posta, T: Tapalqué, G: Guardia del Monte; 1833 frontier adapted from Rolleri et al. (2005)
From the Sauce Posta, Darwin headed to Sierra de la Ventana (Ventania range). There he stayed for around 3 days exploring the ranges which were described as "…a desert mountain of pure quartz rock.Its height is between 3 and 4000 feet …" (Darwin's Beagle Diary (1831-1836). From the notes reported in his diary he probably examined the rock outcrops and the landscape of the upper basin of Rio Sauce Grande and its surroundings, reaching the area of the highest peaks (Cerro Tres Picos). His observations include remarks on the vast extent of the plain surrounding Sierra de la Ventana "…the plain was like the ocean…", (Darwin's Beagle Diary 1831-1836); "...the high plain round this range sinks quite insensibly to the eye on all sides..."(Darwin 1846, p. 79). After surveying this sector of Sierra de la Ventana, he returned to the Sauce Posta. Several days later he continued northward to Sierras de Tandil (Tandilia range). Proceeding to the 3rd Posta, the landscape was perceived as "…a dry grassy plain and on our left hand at a greater or less distance were low hills.." (Darwin's Beagle Diary 1831-1836); this description seems to depict the plain sector northward of Ventania (Fig. 2). From there he travelled around 160 km between the two mountain systems. Heading to the 4th Posta the landscape is reported as "…a low swampy country which extends for nearly 80 miles to the Sierra Tapalken…" (Darwin's Beagle Diary 1831-1836). After crossing this large swampy area Darwin arrived to the 7th Posta in the proximity of Sierra Tapalken (spelled Tapalqué nowadays) depicted as "…a low broken ridge of quartz rock, 2 or 300 feet high extending to the east to Cape Corrientes…" (Darwin's Beagle Diary 1831-1836).
 Figure 2: Geographical location of the Laprida depression, fluvial streams and localities mentioned in the text. 1: Río Quequén Grande, 2: Río Quequén Salado, 3: Río Sauce Grande, T: Tapalqué, S: San Miguel del Monte.
Figure 2: Geographical location of the Laprida depression, fluvial streams and localities mentioned in the text. 1: Río Quequén Grande, 2: Río Quequén Salado, 3: Río Sauce Grande, T: Tapalqué, S: San Miguel del Monte.
From this range Darwin headed to Buenos Aires along the basin of Rio Tapalken (at present arroyo Tapalqué); the 8th and 9th Postas were situated along this stream. On the way to the Rio Salado he described the plain as "…partly swamp and partly good to the east of rio Tapalguen…" (Darwin's Beagle Diary, 1831-1836). He arrived at Guardia del Monte (the present town of San Miguel del Monte) on September 19th, 34 days after disembarking in Patagones, Rio Negro.
THE SOUTHERN PAMPEAN LANDSCAPE
Swampy or dry areas, scarcity or more frequency of tosca outcrops and rounded or flat topped hills were some of the fundamental geographical attributes used by Darwin to describe the landscape of the Buenos Aires Pampas. Southward of Tandilia, he perceived the region as a dominantly dry environment; northward, in the Rio Salado basin, Darwin referred to more fertile lands, the dominance of swampy environments and a general paucity of tosca exposures. The attributes of the landscape and the environmental features reported by Darwin essentially reflect the major geomorphological differences between the so-called Pampa deprimida (depressed Pampa) and Pampa interserrana (intermontane Pampa) (Frenguelli 1950), a still current subdivision of the Buenos Aires Pampean plain (Fig.1). The Tandilia and Ventania mountain systems together with the Pampa interserrana, situated in between the two ranges, constitute the geomorphological expression of a geologically complex structural block named Positivo Bonaerense by Yrigoyen (1975). This positive structural unit separates the Salado and the Colorado tectonic basins (Fig. 3).
 Figure 3: Morphostructural units of Buenos Aires province, modified after Yrigoyen (1975).
Figure 3: Morphostructural units of Buenos Aires province, modified after Yrigoyen (1975).
The Pampa interserrana, characterized by Darwin as a dry plain of tosca higher than the country rock (Darwin 1846, p. 79) is a large-scale plain extending across most of southern Buenos Aires province and continuing southwestward into La Pampa Province, where it has been interpreted as a structural plain (Calmels 1996). The plain is dissected by fluvial systems that drain the central part of Pampa interserrana and the neighboring ranges of Tandilia and Ventania. Fluvial terraces are present along the main river valleys generating a stepped topography. In the upper basins of some of the main rivers (Quequén Grande, Quequén Salado, Sauce Grande), as well as other streams draining the Ventania range, the erosional retreat of the plain surface during the excavation of river valleys gave rise to a hilly and moderately undulating landscape composed of flat-topped and smooth-shaped isolated hills (Fig.4a).

 Figure 4: Plain capped by the tosca rock (calcrete). a) Isolated hill formed by fluvial erosion of the Río Sauce Grande, south of Saldungaray; Sierra de la Ventana at the background; b) Calcrete crust and resulting tableland aspect of the plain by the Río Sauce Chico valley, west of Bahia Blanca.
Figure 4: Plain capped by the tosca rock (calcrete). a) Isolated hill formed by fluvial erosion of the Río Sauce Grande, south of Saldungaray; Sierra de la Ventana at the background; b) Calcrete crust and resulting tableland aspect of the plain by the Río Sauce Chico valley, west of Bahia Blanca.
The plain is capped by a calcareous duricrust (Fig. 4b) or calcrete (Darwin's tosca rock, see below), in turn covered by a large apron of eolian sandy silts and silty sands, around 1.5- 2 meters thick, making up the parent material of the presently cultivated Pampean soils. North-northwestward the landscape grades into a low -gradient area characterized by very poor drainage conditions with numerous shallow lakes and flooded environments, known as Laprida depression (Fig. 2), an endorheic sector (Dangavs 2005) still poorly known. The environmental features of this area fit Darwin's description of a large swampy environment before he reached Sierras de Guitru Gueyú (the present group of low ranges at the northwestern part of Tandilia). Since Darwin's observations on some general morphological characteristics of the southern ranges of Buenos Aires, the geomorphology of Tandilia and Ventania has been the focus of only a few studies, although these have documented significant stages of landscape development, some not yet fully understood. In Sierra de la Ventana, Keidel (1916) pointed out the presence of old erosion surfaces at different altitudes in the area of the upper basin of Rio Sauce Grande. In turn, Teruggi and Kilmurray (1980) mentioned the presence of monadnocks and old peneplain surfaces in the Sierras de Tandil. Recently, a stepped topography composed of erosion surfaces at different elevations was recognized and geomorphologically correlated at both mountain ranges (Demoulin et al. 2005). A model of landscape evolution has been proposed hypothesizing that the highest erosion surfaces of both ranges formed prior to the break-up of Gondwana and the rifting process which led to the opening of the South Atlantic Ocean. This long-lasting history of denudation included intervals of landscape stability and reactivation which are thought to be related to the evolution of the Colorado and Salado tectonic basins (Demoulin et al. 2005). In this context, one of Darwin's observations in Sierra de la Ventana, becomes particularly revealing. He reported a rock outcrop, "…a few small patches of conglomerate and breccia firmly cemented by ferruginous matter at a height of 300 to 400 m…" (Darwin 1846, p. 79). This description is in agreement with the geomorphic setting and the general lithological characteristics of Brecha Cerro Colorado (Andreis et al. 1971) or Conglomerado Rojo (Harrington 1936), a deposit associated with one of the regional erosion surfaces identified in Sierra de la Ventana (Keidel, 1916, Demoulin et al. 2005). Northward of Tandilia range, in the domain of the Salado tectonic basin, the landscape can be subdivided into two main geomorphic settings. Along the Sierras de Tandil, Tricart (1973) pointed out the occurrence of extensive areas of fluvial discharge interpreted as alluvial fans that reached the Río Salado. Later, Fidalgo (1983) described a 20-km wide piedmont region and also identified alluvial fan-like landforms exhibiting a distributary drainage pattern. The area of arroyo Tapalqué visited by Darwin is located in this piedmont setting, characterized by a progressive altitudinal decrease northward, reflecting a gradual change to imperfect drainage conditions. A very flat topography with gradients ranging from 0.1% to 0.01% typifies the central part of the Salado tectonic basin; numerous shallow lakes (lagunas) and swampy environments are present. The Río Salado floodplain, several kilometers wide in some areas, and including shallow lakes interconnected during flooding events, was crossed by Darwin who pointed out: "…12th to 13th to 14th posta: we had to ride for a long distance in water above the horse knees..." (Darwin's Beagle Diary, 1831-1836). After passing the Río Salado and before arriving to Guardia del Monte, he stayed at a large estancia owned by Juan Manuel de Rosas. Several isolated hills (Los Cerrillos), occur in this sector. These features are around 10 meters above the surrounding plain and are located on the eastern side of shallow lakes (Fig. 6). The hills represent lunettes generated by eolian deflation of the topographic depressions, presently occupied by shallow water bodies, under drier conditions (Tricart 1973, Dangavs 2005).

 Figure 6: The Río Salado Valley, south of San Miguel del Monte (Guardia del Monte). a) Hill on the northeastern-eastern side of the Río Salado floodplain; b) Quarry excavated at the hill showing a 3 m thick loess section; present soil developed on top.
Figure 6: The Río Salado Valley, south of San Miguel del Monte (Guardia del Monte). a) Hill on the northeastern-eastern side of the Río Salado floodplain; b) Quarry excavated at the hill showing a 3 m thick loess section; present soil developed on top.
Stratigraphy and lithology of Pampean sediments: the meaning of the Pampean Formation
Darwin typified the sediments of the Pampean Formation as "...a dull reddish, slightly indurated argillaceous earth or mud, often but not always, including in horizontal lines concretions of marl, and frequently passing into a compact marly rock..." (Darwin 1846, p. 89). The reddish argillaceous earth was named "Pampean mud". Darwin emphasized the great extent of the Pampean Formation as one of its most striking features. According to his own observations and the reports from d'Orbigny and other travelers he considered that the unit extended from the vicinity of Rio Colorado in the south to beyond Santa Fe in the north "…M. d'Orbigny traced it for 250 miles further north…"(Darwin 1846, p. 97); and from Banda Oriental (Uruguay) in the east to Cordoba to perhaps Mendoza in the west. Finally he stated "… the area of the Pampean Formation, as remarked by d'Orbigny is at least equal to that of France and perhaps twice or three as great…" (Darwin 1846, p. 97). Thus Darwin correctly perceived what we now know to be the great geographic extent of the Pampean Formation. Darwin also stressed the lithological homogeneity of the sediments that he grouped into a single unit, the Pampean Formation. The concept of Formation (gebirgsformation), as a geological unit, was coined by Abraham Gottlob Werner (1749-1817), a German geologist from the School of Mines at Freiberg, with the purpose of defining a historical entity that included rock bodies formed during the same time interval; at a higher hierarchical rank, the term terrain grouped several formations. Both terms, formation and terrain, although representing units of different stratigraphic hierarchy, were later used as synonyms by other authors. Nevertheless, during the decade of the 1840s, the term formation took root in the Anglo-Saxon community, while terrain, as a synonym of formation, was adopted in the French geology literature (Podgorny et al. 2008). Since Darwin's visit, several other researchers have stressed the apparent lithological homogeneity of the Pampean sediments. At a regional scale of analysis the deposits look rather alike, dominantly composed of sandy silt and silts of light brown to reddish brown colors, common presence of calcareous accumulations of various morphologies and a general massive appearance. This perception of lithological homogeneity was accentuated by the nature of the exposures, scattered and discontinuous outcrops of relatively reduced thickness and mostly situated along river banks, which constrain the detailed analysis of the spatial and temporal relationships of the sediments. Exceptions were the exposures along the Río Paraná and the Monte Hermoso seacliff, where Darwin was able to examine the deposits and their paleontological content in better exposures. To examine the Pampean subsurface he described the lithology of some wells in the vicinity of Buenos Aires and south of Bahia Blanca:"… in a well at the depth of seventy feet, according to Ignatio Nunez, much tosca-rock was met with, and at several points, at 100 feet deep, beds of sand have been found..." (Darwin 1846, p. 77). Also, 50 km SE Buenos Aires, along the coast of Rio de la Plata at Ensenada (Fig. 2), Darwin examined the exposures of a quarry "…specimens from near Ensenada, given me by Sir W. Parish, where the rock is quarried just beneath the surface of the plain..." (Darwin 1846, p. 78). Since the mid-XIX century, many contributions have been focused on the stratigraphy and several different classifications were proposed. The purpose herein is not to make a review of the different stratigraphic classifications, but to stress the concepts and ideas on which the subdivision of the Pampean sediments is based. Up to the present, however, Pampean stratigraphy has remained a controversial and debatable issue; various different schemes are still used simultaneously (Table 1).
TABLE 1: Some of the best known stratigraphic schemes of the Pampean sediments. Those by Fidalgo et al. (1973) and Riggi et al. (1986) are currently used. 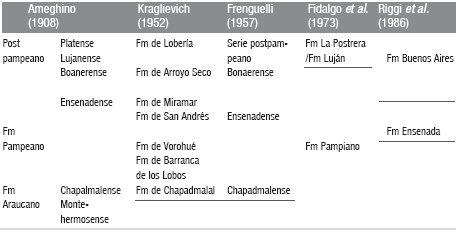
The homogeneity of the deposits together with the discontinuity of the exposures forced the search of a criteria other than the lithology to separate the Pampean sediments. Eventually this led to the utilization of the fossil vertebrate content to separate the deposits into different stratigraphic units. Therefore, those localities with exposures laterally continuous for several km as the Mar del Plata sea-cliffs, the Río Paraná bank exposures and the sea-cliff at Monte Hermoso were subdivided on the basis of their paleontological content (among others Ameghino 1908, Frenguelli 1957, Kraglievich 1952). At the end of the 19th century, the stratigraphic analysis of the Pampean deposits was benefited significantly by deep excavations during the construction of the harbours at Buenos Aires and La Plata. These temporary exposures provided sections several meters thick that became type localities, not later available, to characterize the uppermost interval of the Pampean deposits (Ameghino 1908). The resulting schemes became the groundwork that guided subsequent stratigraphic studies. Much later, the identification of units on the basis of the vertebrate fossil content evolved into the concept of land-mammal ages (Pascual et al. 1965), a fundamental stratigraphic tool still widely accepted and used. In recent years, these biostratigraphic units have been redefined and updated (Cione et al. 2007 and references therein). The relative degree of evolution inferred from the fossil assemblages was used in the stratigraphic arrangement of the Pampean deposits. Accordingly, and considering their paleontological content, several units were identified, and classified as lithostratigraphic units (formations) receiving their names after the locality where they were described (among others, Epecuén Formation, Saldungaray Formation, Irene Formation, Arroyo Chasicó Formation; Fidalgo et al. 1975 and references therein). As time passed, in spite of the numerous schemes proposed, Pampean Formation or simply Pampeano developed into terms that became deeply rooted in the Argentine geological literature; both became synonyms to refer to the Pampean deposits. At present, the term Pampeano is still used informally with a rather loose significance. However, general agreement exists to separate the surficial Late Pleistocene-Holocene sedimentary cover from the underlying Pampean deposits. Following this line of reasoning, the prelate Pleistocene deposits exposed in the Pampa interserrana and the Salado Basin were grouped again into a single unit, the Formación Pampeano (Pampean Formation) identified exclusively on the basis of lithological attributes (Fidalgo et al. 1973). This proposal is somewhat close to the original meaning given by Darwin to the term Pampean Formation. Regardless of its vertebrate fossil content, the unit defined is nevertheless useful for mapping purposes and well descriptions. More recent schemes tend to divide the upper part of the Pampean deposits into two units (Ensenada and Buenos Aires Formations) distinguished on the basis of their paleontological content (Table 1). In turn, the uppermost sedimentary mantle was subdivided into several lithostratigraphic units that represent lithofacies of different geomorphic processes and environmental settings such as eolian, fluvial and littoral deposits (among others Fidalgo et al. 1973 a,b).
The Pampean sediments today: current ideas and interpretations
At present the Pampean sediments are lithologically characterized as light brown to reddish brown sandy silts or silty sands, moderately indurated with a general massive appearance. Darwin's interpretation was that the "...deposits were accumulated in the former estuary of Rio de la Plata and in the sea adjoining it…" (Darwin 1846, p. 99); the source area of the Pampean mud was thought to come from the weathering of igneous and metamorphic rocks of Brazil and Uruguay (Darwin 1846, p. 100). The mineral composition of the Pampean sediments reflects their derivation from volcaniclastic material in turn mostly derived from Andean rocks, with minor contributions (Precambrian granitic and metamorphic rocks and Paleozoic quartzites) from the Tandilia and Ventania ranges in the southern part. Northward, the Pampean ranges of Córdoba and San Luis and the Brazilian shield were secondary source areas (Teruggi 1957, González Bonorino 1965, Zárate 2003 and references therein). A detailed analysis of the deposits reveals grain size changes and variations in the density, color, and mineralogical composition according to the sediment age and the geographic setting considered within the Pampean plain. The massive appearance and the rather uniform lithology, among other attributes, were usually taken as evidence of an eolian origin and the sediments have been interpreted as loess. Nervertheless, Teruggi (1957) pointed out that the distribution of primary loess was quite limited, a statement reconfirmed by more recent studies (Zárate 2003 and references therein). Across the Pampean environment, primary loess facies have a low preservation potential. The deposits are locally reworked either by aqueous transport agents (loess-like or loessoid deposits) or modified by pedogenesis (weathered loess). Loess-like deposits exhibit a weak and poorly expressed sedimentary structure consisting either of horizontal bedding or cross bedding. Along the piedmont areas, channelized lithofacies representing fluvial paleoenvironments include rock fragments from bedrock of the Tandilia and Ventania ranges. At distal settings from the ranges, the coarse fluvial facies are made up of bones and tosca fragments embedded in a sandy matrix. Paleosols are a very common feature with pedogenic traits (pedogenic structure, textural changes resulting from eluviation, carbonate leaching, evidences of bioturbation features from soil microfauna and vegetation) distributed throughout the Pampean sediments. Welded paleosols resulting from the superposition of pedogenetic process have been reported in both Pliocene and Pleistocene stratigraphic sections of the Pampean succession. Micromorphological studies also point to the occurrence of intervals when sediments accumulated progressively on the surface while soil formation was active, resulting in accretionary soil surfaces. From a sedimentological perspective, the occurrence of numerous paleosol levels throughout the deposits documents an episodic sedimentation process. Due to the ubiquity of pedogenic features, the Pampean succession is interpreted as a pedosedimentary sequence (i.e. Kemp and Zárate 2000, Zárate et al. 2002). The presence of welded paleosols and intervals of accretionary pedogenesis suggest a variable balance of pedogenesis and sedimentation through time. At some sections, the occurrence of discrete paleosol levels, stratigraphically superposed, allow inferring some sort of cyclicity in soil formation that has been related to climatic changes (among others Nabel et al. 2000, Kemp and Zárate 2000). Yet not assessed, bioturbation structures are an outstanding feature of the Pampean sediments. Not only are there traces of invertebrates in the sediments, particularly those beds more intensely modified by pedogenesis, but also burrows, caves and galleries (krotovinas) excavated by small rodents and larger mammals, mainly xenarthrans (Vizcaino et al. 2001). The resulting structures are filled in by diamicton-like deposits, including fragments of tosca, paleosol aggregates and bones redeposited by water. Also, some burrows and caves are filled with eolian and/or fine grained laminated sediments. In the Pampean sediments, minor erosive episodes are recorded by both the partial truncation of paleosol surfaces and the local incision of fluvial paleochannels (Ruocco 1989, Zárate and Fasano 1989). Three major episodes of landscape reactivation, inferred from stratigraphic analysis and gemorphological evidence, are documented by cycles of fluvial incision. These generated erosional unconformities in the sedimentary record as well as terrace formations and different aggradational surfaces in the Sierra de la Ventana piedmont (Zárate 2005).
Carbonate accumulations of the Pampean sediments: the Tosca-rock
Darwin gave special attention to the calcareous accumulations in his general characterization of the Pampean Formation. The duricrust (calcrete) that caps the sediments in the Pampa interserrana area, and constitutes massive accumulations in some other locations, was described as a marly rock and named tosca-rock, after the popular term given to the carbonate deposits by the local Pampean inhabitants (Darwin 1846). He reported the morphology (nodular and stalactiform concretions, compact stratified tosca rock), the relation with the Pampean mud, as well as the geographical distribution and relative occurrence (more frequent outcrops of tosca in the area of the ranges and the Pampa interserrana, general scarcity of tosca exposures in the Rio Salado basin, almost no tosca-rock in northeastern Buenos Aires province:"… At Arroyo del Medio the bright red Pampean mud contains scarcely any tosca-rock…", "… At Rosario there is but little tosca-rock..." (Darwin 1846, p. 87). He also made comparisons of tosca samples collected throughout the Pampas, referring to their similar characteristics (the absolute identity in his own words) of specimens collected in the northern Pampas, Tandilia (Tapalqué) and south of Bahia Blanca. In addition, thin sections of tosca were petrologically analyzed by a colleague "...Dr Carpenter has kindly examined under the microscope, sliced and polished specimens of these concretions and of the solid marl-rock collected in various places between the Colorado and Sta Fe Bajada…." (Darwin 1846, p. 77). Despite the economic importance of calcareous crusts as a source of road building material and in agricultural development across most of the southern Buenos Aires Pampean plain, no detailed and systematic studies of these materials have been carried out. In the Positivo Bonaerense area, the morphology and thickness of calcretes vary according to the relative age of the geomorphic surfaces that they cover. The oldest crust capping late Miocene-Pliocene deposits is up to 1.5-2 m thick. The general morphology of calcretes suggests a high degree of relative development close to morphological stages 5 and 6 as defined by Machette (1995). Thus, a typical calcrete section is usually composed of three main parts from top to bottom. The uppermost part is made up of platy calcrete consisting of 5-10 cm thick indurated layers, showing an irregular horizontal fracturing (Fig 5B). Lamination, brecciation, and dissolution are very common features suggesting a very complex history of multiple episodes of carbonate precipitation and weathering on the surface. The middle part consists of powdery and massive carbonate grading downward into the host Pampean sediments or showing a very sharp lower contact (erosional surface) on those deposits. Carbonate nodules, concretions and massive accumulation of tosca are also very frequently found. Studies on specific accumulations suggest different mechanisms of formation, such as precipitation from phreatic waters, capillary rise, and pedogenic carbonate leaching (Bk horizons) (Tricart 1973, Imbellone and Teruggi 1986).

 Figure 5: Plain edge in the area of Bahía Blanca. a) Plain surface with a thin eolian cover and fragments of calcrete, Atlantic Ocean in the background; b) Calcrete section at a quarry showing typical platy structure in upper part, grading downward into Pliocene sediments. Note hammer for scale.
Figure 5: Plain edge in the area of Bahía Blanca. a) Plain surface with a thin eolian cover and fragments of calcrete, Atlantic Ocean in the background; b) Calcrete section at a quarry showing typical platy structure in upper part, grading downward into Pliocene sediments. Note hammer for scale.
Age and origin of the Pampean sediments
On the basis of its lithological homogeneity (similar composition) and "...the apparent absolute specific identity of some of its mammiferous remains..." (Darwin 1846, p. 100), Darwin inferred that all over its large area the Pampean Formation belonged to the same geological epoch. This age interpretation followed Charles Lyell's ideas, where the percentages of living and extinct mollusks was a fundamental tool for assigning relative ages to the Tertiary strata. He collected some of the mollusk shells from late Pleistocene-Holocene marine deposits exposed along the Pampean coastal fringe. At Punta Alta Darwin considered that the proportion and specimens of shells found in the sediments was comparable to those species living today in the bay. Therefore, taking into account the paleontological content including living species of mollusks that he considered contemporaneous with the mammiferous remains found in the sediments, Darwin attributed the Pampean Formation to the Recent Period or what is now called the Holocene. From a stratigraphic viewpoint, Darwin recognized stages in the Pampean Formation on the basis of both the different altitudes shown by the outcrops of Monte Hermoso, Punta Alta and Sierra de la Ventana, and an observation now regarded as a taphonomic attribute of a fossil remnant: "...From the rolled fragment of black bone and from the plain of Punta Alta being lower than that of Monte Hermoso, I conclude that the course sub-littoral deposits of Punta Alta are of subsequent origin to the Pampean mud of Monte Hermoso; and the beds at this latter place… are probably of subsequent origin to the high tosca-plain round the Sierra Ventana..." (Darwin 1846, p. 87). The Pampean succession is now interpreted as the late Cenozoic (approximately the last 12 Ma) continental record of central Argentina. Sediments of the Pampean succession accumulated after a Miocene regression of the Paranense sea (Folguera and Zárate, submitted). Major differences are evident with regard to the regional distribution of the deposits. Fidalgo et al. (1975) mapped the exposures bearing Late Miocene and Pliocene vertebrate fossil assemblages, showing a distribution restricted to the Positivo Bonaerense area. Darwin examined an outcrop on the bank of the Río Sauce Grande noting that "...there is an imperfect section about 200 feet high displaying in the upper part tosca rock and in the lower part red pampean mud...". This likely corresponds to late Miocene?-early Pliocene sandy siltstones. Several other outcrops are discontinuously exposed along other neighboring river systems as well as some railroad cuts. Northward, in the Río Salado valley, the relatively oldest exposures of Pampean sediments are much younger (Pleistocene sensu lato, see below). These deposits consist of compacted clayish silts and sandy silts, showing evidence of hydromorphic conditions (mottles, greenish colors, manganese nodules, massive carbonate accumulations). Considering the stratigraphic relations, numerical ages and relative ages inferred from the vertebrate fossil assemblages, the Pampean succession of sedimentation has been recently subdivided into four stratigraphic intervals, interpreted as subcycles of sedimentation (Zárate 2005) (Table 2).
TABLE 2: Age, geographic distribution and fossil mammal remains of the subcycles of sedimentation identified in the Pampean deposits (after Zárate 2005). 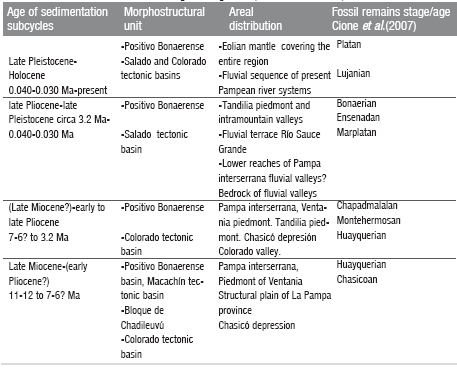
The oldest sedimentary subcycle, is exposed in the Positivo Bonaerense and the northern part of the Colorado tectonic basin. It is composed of Late Miocene deposits (11-12 Ma to circa ?7-6 Ma) bearing Chasicoan and Huayquerian fossil remains. These deposits have an average thickness of 150-200 m, and overlie the Precambrian and Paleozoic bedrock of Pampa interserrana. The geographical distribution of this unit is broad, extending southwestward into La Pampa province (Folguera and Zárate, submitted). Geomorphologically, these late Miocene deposits, capped by the thickest calcrete crust form the very extensive plain within which the present river valleys were excavated. The second sedimentary subcycle, represented by late Miocene?-early to late Pliocene deposits (?7-6-3.2 Ma) is discontinuously exposed along the river banks of the drainage system. The deposits include Huayquerian, Montehermosan and Chapadmalalan fossil remains. The outcrops are located in the Positivo Bonaerense and distributed throughout the Sierra de la Ventana piedmont, the valleys of Quequén Salado, Quequén Grande as well as of minor streams of the southern part of Pampa interserrana, and along the lower section of the Mar del Plata-Chapadmalal sea-cliffs. The Monte Hermoso stratigraphic section (Quattrocchio et al. 2009) is included in this cycle of sedimentation. Farther southward, deposits of comparable age crop out in the Colorado tectonic basin. Exposures up to 15 meter thick and 200-300 m long can be found at various quarries in the surroundings of Bahia Blanca. The sediments consist of fluvial facies, including rock fragments from Sierra de la Ventana, capped by a 1-1-5 meter thick calcrete (Folguera and Zárate, submitted). Darwin explored the very top part of the exposures (Fig. 5a) "…At the settlement of Bahia Blanca, the uppermost plain is composed of very compact, stratified tosca rock containing rounded grains of quartz distinguishable by the naked eye...". Further eastward, deposits of this cycle still poorly known, crop out along the lower reaches of the Quequén Salado and the Quequén Grande rivers; Huayquerian, Montehermosan and Chapadmalalan fossil remains have been exhumed (among others Verzi et al. 2008). Along the Mar del Plata-Chapadmalal sea cliffs, this stratigraphic interval is composed of early to late Pliocene (3.2 to 4.5-5 Ma), massive siltstones with well developed paleosols bearing Chapadmalalan fossil remains. The third subcycle of sedimentation (late Pliocene-late Pleistocene circa 3.2 Ma- 0.040/0.030 Ma) includes deposits bearing Marplatan, Ensenadan and Bonaerian fossil remains. They are distributed in the Salado tectonic basin, forming the bedrock in which fluvial valleys are excavated. The lower sections of deep quarries sometimes penetrate levels containing Ensenadan fossil mammal remains. Rabassa (1973) reported sediments bearing Ensenadan fossil remains in the central part of Tandilia range. These deposits are also discontinuously exposed along the SW piedmont of Tandilia, along the Atlantic coast between Mar del Plata and Miramar. The late Pliocene-late Pleistocene subcycle unconformably overlies Pliocene siltstones along the Mar del Plata sea cliff. Included in this subcycle are the Sauce Grande fluvial terrace sediments with Bonaerian fossil remains reported by Deschamps (2005). Along the Atlantic coast, between Miramar and west of Necochea, the sea cliffs are composed of fine sandy facies including diamicton layers with a high content of calcareous nodules and capped by a platy calcrete. Inland, at a distance of several kilometers from the coastline, the uppermost 2 meters of these deposits are exposed in numerous quarries and roadcuts, suggesting that they form an extensive plain. The stratigraphic and geomorphologic relationship with older deposits is still unknown. The lithofacies are similar to the Pleistocene exposures of Mar del Plata and might be part of the late Pliocene-late Pleistocene subcycle of sedimentation which should be confirmed in future studies. The most recent subcycle of sedimentation (40-30 ka-to the present) is composed of a large eolian apron along with fluvial deposits accumulated during the last glacial cycle. Recent numerical ages obtained in fluvial deposits of northern Buenos Aires point to an older age for the beginning of this cycle of sedimentation (Blasi et al. 2008). The eolian lithofacies constituting the parent material of the present cultivated soils are composed of sandy deposits towards the west, grading into sandy silts and silt east and northeastward. In the Pampa interserrana area the eolian mantle consists of loessial sands, sand mantles, and sandy loess (Zárate and Blasi 1993, Bidart 1996), representing proximal eolian facies derived by deflation from the Río Colorado flood-plain. In the area of the Río Salado valley, the eolian facies are made up of silty deposits (loess) named La Postrera Formation by Fidalgo et al. (1973) (Fig. 6b).
FINAL REMARKS
Darwin as well as many other researchers after him emphasized the relatively uniform lithological composition, the vast extent and the rich vertebrate fossil content of the Pampean sediments. His perception of the Pampean environment illustrated not only the major landscape differences but also some key lithological features of the deposits. To date, the geographical distribution of exposures of different ages suggests significant variations of the sedimentary dynamics throughout the region, and four main subcycles of sedimentation have been identified. The outcrops of the first two cycles recording the late Miocene and most of the Pliocene sedimentation are restricted to the Positivo Bonaerense (Sierra de la Ventana piedmont, part of Pampa interserrana and Sierras de Tandil). A remarkable geomorphological characteristic of this southern area is the occurrence of several intervals of landscape stability documented by the development of calcrete crusts (tosca-rock) on topographic surfaces of different ages. During the very late Pliocene-Pleistocene, sedimentation continued in the piedmont of Sierras de Tandil and the Salado tectonic basin. In the meantime, the Positivo Bonaerense was geomorphologically much more stable with episodes of fluvial incision along the main river systems. Yet, no information is available on the Laprida depression, the swampy environment crossed by Darwin, and the Atlantic fringe where possible Pleistocene deposits are exposed. In both the Positivo Bonaerense and the Salado tectonic basin, the last subcycle of sedimentation is related to incision of the present fluvial valleys sometime circa 40 ka and the accumulation of an eolian mantle during the last glacial cycle. What was the primary control of sediment accumulation at a regional scale?. Why did sedimentation cease in the late Miocene over a vast area of Positivo Bonaerense?. Which factor triggered the subcycles of sedimentation now identified?. At present, research in progress is addressing these main questions. The studies under way are focused on the potential role that might have been played by both Andean tectonism and tectonic dynamics of the Colorado and Salado basins. With this purpose, future stages of current research will be mainly directed towards establishing improved chronological control of the deposits along with detailed mapping and sedimentological analysis of the areas, still poorly known (the northern area of Pampa interserrana, the Laprida depression and the Atlantic sea cliff west of Mar del Plata. Last but not least, the Geological Observations on South America was read by one of us (MZ) in the early 1980s. From the present perspective, after almost three decades of exploring the geology of this vast environment, Darwin's careful and detailed descriptions and his interpretations, capture the attention of those of us who have devoted time and patience to understanding the nature of the Pampas.
ACKNOWLEDGEMENTS
We thank Beatriz Aguirre-Urreta for inviting us to participate in this special volume. The critical and helpful comments and suggestions on the text by the reviewers Daniel Muhs, Edward Derbyshire and Cecilia Deschamps, improved and smoothed the manuscript. We are grateful to Alejandra Pupio and Irina Podgorny for helping us with literature and information used in this paper. Vivi Martinez helped with the illustrations. This paper was financially supported by Universidad Nacional de la Pampa (Research P.186-06), and is a contribution to PIPCONICET 5627.
WORKS CITED IN THE TEXT
1. Andreis, R., Spalletti, L. and Mazzoni, M. 1971. Sedimentología de la Brecha Cerro Colorado, Sierras Australes de la provincia de Buenos Aires. Reunión sobre Geología Sierras Australes Bonaerenses, Provincia de Buenos Aires (La Plata): 65-96.
2. Ameghino, F. 1908. Las formaciones sedimentarias de la región litoral de Mar del Plata y Chapadamalan. Anales del Museo Nacional de Buenos Aires 3(10): 343-428.
3. Bidart, S. 1996. Sedimentological study of aeolian soil parent materials in the Río Sauce Grande basin, Buenos Aires province, Argentina. Catena 27: 191-207.
4. Blasi, A., Prieto, A., Castiñeira, C., Fucks, E., del Puerto, L., De Francesco, C., Figini, A., Carbonari, J., Huarte R., and Hanson, P. 2008. Variaciones ambientales y climáticas durante el Pleistoceno tardío en la cuenca media del río Luján, Argentina. 12° Reunión Argentina de Sedimentología, Resúmenes: 43, Buenos Aires.
5. Calmels, P. 1996. Bosquejo geomorfológico de la provincia de la Pampa. Universidad Nacional de la Pampa, Facultad de Ciencias Exactas y Naturales, 110 p., Santa Rosa.
6. Cione, A.L., Tonni, E.P., Bargo, M.S., Bond, M., Candela, A.M., Carlini, A.A., Deschamps, C. M., Dozo, M.T., Esteban, G., Goin, F.J., Montalvo, C.I., Nasif, N., Noriega, J.I., Ortiz-Jaureguizar, E., Pascual, R., Prado, J.L., Reguero, M.A., Scillato-Yané, G.J., Soibelzon, L., Verzi, D.H., Vieytes, E.C., Vizcaino, S.F. and Vucetich, M.G. 2007. Mamíferos continentales del Mioceno tardío a la actualidad en la Argentina: los últimos cincuenta años de investigaciones. Ameghiniana 50º aniversario, Publicación especial 11: 257-278.
7. Dangavs, N.V. 2005. Los ambientes acuáticos de la provincia de Buenos Aires. In de Barrio, R., Etcheverry, R., Caballé M. and Llambías, E. (eds.) Geología y Recursos Minerales de la Provincia de Buenos Aires, Relatorio 16° Congreso Geológico Argentino: 219-236, La Plata.
8. Darwin's Beagle Diary 1831-1836. In The Complete Work of Charles Darwin. www.darwinonline. org.uk,University of Cambridge, Cambridge.
9. Darwin, C. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 280 p., London.
10. Demoulin, A., Zárate, M. and Rabassa, J. 2005. Long-term landscape development and denudation rates: the Buenos Aires pampean (Argentina) perspective. Journal of South American Earth Sciences 19: 193-204.
11. Deschamps, C.M. 2005. Late Cenozoic mammal bio-chronostratigraphy in southwestern Buenos Aires Province, Argentina. Ameghiniana 42: 733-750.
12. d' Orbigny, A. 1842. Voyage dans l'Amérique méridionale... exécuté pendant les années 1826, 1827, 1828, 1829, 1830, 1831, 1832 et 1833. Tome troisième. 3e partie, Géologie. Bertrand, 290 p., Paris.
13. Fidalgo, F. 1983. Algunas características de los sedimentos superficiales en la cuenca del río Salado y en la Pampa ondulada. In Coloquio Internacional sobre Hidrología de Grandes Llanuras, 11-20 abril 1983, Olavarria, Argentina. Comité Nacional para el Programa Hidrológico Internacional: 1045-1067, Buenos Aires.
14. Fidalgo, F, De Francesco, F. and Colado, U. 1973a. Geología superficial en las Hojas Castelli, J.M. Cobo y Monasterio (Pcia. de Buenos Aires). 5° Congreso Geológico Argentino, Actas 4: 27-39.
15. Fidalgo, F, Colado, U. and De Francesco, F. 1973 b. Sobre ingresiones marinas cuaternarias en los partidos de Castelli, Chascomús y Magdalena (Provincia de Buenos Aires). 5° Congreso Geológico Argentino, Actas 4: 225-240.
16. Fidalgo, F., De Francesco, F. and Pascual, R. 1975. Geología superficial de la llanura bonaerense. In Relatorio Geología de la Provincia de Buenos Aires, 6° Congreso Geológico Argentino: 103-138.
17. Folguera, A. and Zárate, M.(submitted). La sedimentación neógena continental en el sector extraandino, Argentina central. Revista de la Asociación Geológica Argentina.
18. Frenguelli, J. 1950. Rasgos generales de la morfología y la geología de la provincia de Buenos Aires. LEMIT, serie 2, 33: 1-72, La Plata.
19. Frenguelli, J. 1957. Neozoico. Sociedad Argentina de Estudios geográficos GAEA 2, 3, 218 p., Buenos Aires.
20. González Bonorino, F. 1965. Mineralogía de las fracciones arcilla y limo del Pampeano en el área de la ciudad de Buenos Aires y su significado estratigráfico y sedimentológico. Revista de la Asociación Geológica Argentina 20: 67-148.
21. Harrington, H.J. 1936. El Conglomerado Rojo de las sierras australes de la provincia de Buenos Aires y sus relaciones con el relieve de montaña. Obra del Cincuentenario, Museo de la Plata 2: 145-184, La Plata.
22. Imbellone, P. and Teruggi, M.E. 1987. Morfología y micromorfología de toscas en paleosuelos de los alrededores de la Plata. Ciencia del Suelo 4: 209-215.
23. Keidel, J. 1916. La geología de las sierras de la provincia de Buenos Aires y sus relaciones con las montañas del Cabo y los Andes. Anales Dirección Nacional Geología Minería 9(3): 5-78, Buenos Aires.
24. Kemp, R. and Zárate, M. 2000. Pliocene pedosedimentary cycles in the southern Pampas, Argentina. Sedimentology 47: 3-14.
25. Kraglievich, L. 1952. El perfil geológico de Chapadmalal y Miramar, provincia de Buenos Aires. Resumen preliminar. Revista Museo Municipal de Ciencias Naturales y Tradicional de Mar del Plata 1(1): 8-37.
26. Machette, M.N. 1995. Calcitic soils of southwestern United States. In Weide, D.L. (ed) Soils and Quaternary Geology of the southwestern United States. Special Paper, Geological Society of America 203: 1-21.
27. Nabel, P.E., Cione, A. and Tonni, E. 2000. Environmental changes in the pampean area of Argentina at the Matuyama-Brunhes boundary (C1r-C1n) Chrons boundary. Palaeogeography, Palaeoclimatology, Palaeoecology 162: 403-412.
28. Pascual, R., Ortega Hinojosa, J., Gondar, D., Tonni, E.P. 1965. Las edades del cenozoico mamalífero de la Argentina, con especial atención a aquellas del territorio bonaerense. Anales Comisión de Investigaciones Científicas de la Provincia de Buenos Aires 6: 165-193.
29. Podgorny, I. Ballestero, D., Farro, M., García S., Pegoraro, A., Pupio, A., Reguero, M. and Zárate, M. 2008. Las formaciones geológicas sudamericanas en los viajes de Charles Darwin y Alcide d'Orbigny.Mapas geológicos, fósiles e itinerarios Registros 5. Revista Anual de Investigación del Centro de Estudios Históricos Arquitectónico-Urbanos. Facultad de Arquitectura, Urbanismo y Diseño, UNMDP: 136-157.
30. Quattrocchio, M.E., Deschamps, C.M., Zavala, C.A., Grill, C.S. and Borromei, A.M. 2009. Geology of the area of Bahía Blanca, Darwin's view and the present knowledge: a story of 10 million years. Revista de la Asociación Geológica Argentina 64(1): 137-146.
31. Rabassa, J. 1973. Geología superficial en la hoja "Sierras de Tandil", provincia de Buenos Aires. LEMIT Anales 3, Serie 2, 240: 115-160.
32. Riggi, J.C., Fidalgo, F., Martinez, O., and Porro, N. 1986. Geología de los sedimentos pampeanos en el partido de la Plata. Revista de la Asociación Geológica Argentina 44: 316-333.
33. Rolleri, E., Caballé, M. and Tessone, M. 2005. Breve reseña histórica y apuntes sobre los avances del conocimiento geológico del territorio bonaerense. In de Barrio, R., Etcheverry, R., Caballé, M. and Llambías, E. (eds.) Geología y Recursos Minerales de la Provincia de Buenos Aires, Relatorio 16° Congreso Geológico Argentino: 1-20, La Plata.
34. Ruocco, M. 1989. A 3 Ma paleomagnetic record of coastal continental deposits in Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology 72:105-113.
35. Teruggi, M.E. 1957. The nature and origin of Argentine loess. Journal of Sedimentary Petrology 27: 322-332.
36. Teruggi, M. and Kilmurray, J. 1980. Sierras Septentrionales de la Provincia de Buenos Aires. In Turner, J.C.M. (ed.) Segundo Simposio de Geología Regional Argentina 2: 918-965, Academia Nacional de Ciencias, Córdoba.
37. Tricart, J.L. 1973. Geomorfología de la Pampa Deprimida. Base para los estudios edafológicos y agronómicos. INTA, 12º Colección Científica, 202 p., Buenos Aires.
38. Verzi, D., Montalvo, C. and Deschamps, C. 2008. Biostratigraphy and biochronology of the late Miocene of central Argentina: evidence from rodents and taphonomy. Geobios 41: 145-155.
39. Vizcaino, S.F., Zárate, M.A., Bargo, M.S. and Dondas, A. 2001. Pleistocene burrows in the Mar del Plata area (Buenos Aires province, Argentina) and their probable builders. Acta Paleontologica Polonica 46: 289-301.
40. Yrigoyen, M. 1975. Geología del subsuelo y la plataforma continental. In Relatorio Geología de la Provincia de Buenos Aires, 6to Congreso Geológico Argentino: 139-168, Buenos Aires.
41. Zárate, M. 2003. The Loess record of Southern South America. Quaternary Science Reviews 22: 1987-2006.
42. Zárate, M. 2005. El Cenozoico tardío continental de la provincia de Buenos Aires. In de Barrio, R., Etcheverry, R., Caballé, M. and Llambías, E. (eds.) Geología y Recursos Minerales de la Provincia de Buenos Aires. Relatorio 16° Congreso Geológico Argentino: 139-158, La Plata
43. Zárate, M. and Blasi, A. 1993. Late Pleistocene-Holocene eolian deposits of the southern Buenos Aires Province, Argentina: a preliminary model. Quaternary International 17: 15-20.
44. Zárate, M. and Fasano, J. 1989. The Plio-Pleistocene record of the central eastern Pampas, Buenos Aires province, Argentina: the Chapadmalal case study. Palaeogeography, Palaeoclimatology, Palaeoecology 72: 27-52.
45. Zárate, M., Kemp, R.A, and Blasi, A. 2002. Identification and differentiation of Pleistocene paleosols in the northern pampas of Buenos Aires, Argentina. Journal of South American Earth Sciences 15: 303-313.
Mirta E. Quattrocchio1-2, Cecilia M. Deschamps3, Carlos A. Zavala1-2, Silvia C. Grill1 and Ana M. Borrome1-2
1 Departamento de Geología, Universidad Nacional del Sur. 2 CONICET 3 Museo de La Plata, CIC
Emails: mquattro@criba.edu.ar; ceci@fcnym.unlp.edu.ar; czavala@criba.edu.ar; sgrill2criba.edu.ar; borromei@criba.edu.ar
ABSTRACT
The aim of this paper is to give an updated outlook of the scenery described by Charles Darwin when he visited Bahía Blanca and surrounding areas, following the itinerary during his voyage on board HMS Beagle. Such an outlook is a state of the art of the current understanding of the Late Miocene-Holocene history in the southwestern Pampas (Argentina). Multidisciplinary results were integrated in a chronosequence chart synthesizing the suggested space-time correlation of the recognized events. Some of the studied localities covering the whole time interval represented in the area were arranged in this chart in a hypothetical E-W line crossing the Río Sauce Grande basin and the surrounding highlands. This line is also approximately the one followed in part by Darwin when riding from Bahía Blanca to Tapalqué (Tapalguen) as he crossed the region toward the Río Sauce. Paleoenviromental and paleoclimatic inferences for the last 10m.y. are also given. Paleontological studies included vertebrates, ostracods and palynomorphs. Many of the results of these investigations are the answers to Darwin's question when he first visited the area.
Keywords: Pampas; Tapalqué; Pampean Formation; Punta Alta; Red earth cliffs.
RESUMEN: Geología del área de Bahía Blanca, los comentarios de Darwin y el conocimiento actual: una historia de 10 millones de años. El objetivo de este trabajo es brindar una actualización del escenario descrito por Charles Darwin cuando visitó Bahía Blanca y sus alrededores, siguiendo el itinerario de su viaje alrededor del mundo a bordo del Beagle. Se trata de una puesta al día del conocimiento de la historia del sudoeste de la región pampeana (Argentina) desde el Mioceno tardío hasta el Holoceno y tiempos históricos. Las investigaciones multidisciplinarias llevadas a cabo por el grupo de investigación del Laboratorio de Palinología, UNS, Bahía Blanca, se integraron en una carta cronoestratigráfica secuencial, en la cual se sintetizaron las correlaciones espacio-temporales de los eventos reconocidos. En esta carta, algunas de las localidades estudiadas que cubren todo el lapso representado en el área, se ordenaron en una línea hipotética E-O que corta la cuenca del río Sauce Grande y sus divisorias. En parte, esta línea también sigue aproximadamente el primer tramo de la cabalgata de Darwin desde Bahía Blanca hacia Tapalqué (Tapalguén), cuando cruzó la región hacia el Río Sauce. Asimismo, se brinda información paleoambiental y paleoclimática de los últimos 10 ma para esta región. Los estudios paleontológicos incluyeron vertebrados, ostrácodos y palinomorfos. Muchos de los resultados de estas investigaciones son respuestas a comentarios que Darwin hizo en su visita.
Palabras clave: Pampas; Tapalqué; Formación Pampeano; Punta Alta; Acantilados de tierras rojas.
INTRODUCTION
The aim of this paper is to give an updated outlook of the scenery described by Charles Darwin when he visited Bahía Blanca and surrounding areas, following the itinerary during his voyage on board the Beagle (Fig. 1). Most of the information is taken from Quattrocchio et al. (2008) - and references therein - who gave a state of the art account of the understanding of the late Miocene-Holocene history in the southwest Pampas (Argentina) (Fig. 2). This information was assembled by researchers at the Palynology Laboratory of the UNS (Bahía Blanca) over almost 20 years of mapping, stratigraphic studies and analyses of the fossil record. Paleontological studies included vertebrates, ostracods and palynomorphs. Many of the results of these investigations are the answers to Darwin's comments when he first visited the area. Darwin spent 67 days in this area, from September 7 to October 20, 1832, and from August 17 to September 8, 1833. The amount of geological and biological information provided in his notes and further publications, and the acute observations concerning the genesis of the deposits, the past environments and organisms are overwhelming even today, more than one and a half century later. Only those issues that have been revisited by the authors are here discussed.
 Figure 1: Itinerary of Charles Darwin, and below right, the Fortaleza Protectora Argentina. Taken from Dozo and Ginobili (1999
Figure 1: Itinerary of Charles Darwin, and below right, the Fortaleza Protectora Argentina. Taken from Dozo and Ginobili (1999
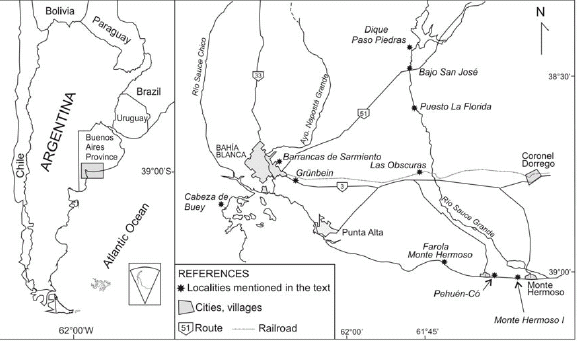 Figure 2: Map of the studied area with the localities mentioned in the tex
Figure 2: Map of the studied area with the localities mentioned in the tex
Charles Darwin mentioned Bahía Blanca, and described this area several times in his reports. The writer Eduardo Mallea (Bahía Blanca, 1903-Buenos Aires, 1982) already underlined this visit when he wrote in his novel "Todo verdor perecerá" (Mallea 1941): "Cuántas civilizaciones humanas habían evolucionado y perecido cuando este pedazo de desierto emergía de su sopor! En torno al fortín, valla opuesta al indio predatorio, comenzó a crecer, hacia los ochocientos veintiocho, la población militar, y cuatro años más tarde Rosas y Darwin se paraban ante aquellos salitrales que después de los secos calores extendían en la bahía su ardiente sábana blanca." ["All greenness will perish": "¡How many civilizations had evolved and perished by the time this piece of desert was emerging from its drowsiness! Surrounding the fortress, boundary for the predatory aborigines, there started to grow, near 1828, the military population, and four years later, Rosas and Darwin stood before those saltpeter beds that, after dry heat waves, spread their fiery white sheet along the bay"].
THE ARRIVAL
Darwin first saw Bahía Blanca when the Beagle anchored there on September 6, 1832. "Baia (sic) Blanca has only been settled within the last six years: previous to which even the existence of the bay was not known. It is designed as a frontier fort against the Indians & thus to connect Buenos Ayres to Rio Negro" (Keynes 2001). He stayed in the area until October 19, visiting Punta Alta and Monte Hermoso. Then the Beagle went back to Montevideo. On November 28 they sailed again south to Patagonia, and when returning to Buenos Aires, he left the Beagle at Patagones and rode to Buenos Aires. On August 17, 1833 he wrote:" Bahia Blanca scarcely deserves the name of a village. A few houses and the barracks for the troops are enclosed by a deep ditch and fortified wall" (Darwin 1845, p. 76). Bahía Blanca is located in the southeastern part of the province of Buenos Aires, Argentina, on the Atlantic Ocean. It has a population of 325,000 inhabitants according to the 2005 census. The city was founded as a fort on 11th. April 1828 by Colonel Ramón Estomba (Fig. 3) under the orders of Brigadier-General and subsequent Governor of Buenos Aires, Juan Manuel de Rosas. The original founding was named Fortaleza Protectora Argentina (Argentine Protective Fortress, Fig. 1), intended to protect settlers and their cattle from raids of native aborigines, and also to protect the coast from the Brazilian navy, which had landed in the area the previous year. The fort was attacked by malones (incursions of nomad aborigines on horseback) several times, most notably in 1859 by 3,000 Calfucurá warriors.
 Figure 3: Map of Bahía Blanca city.
Figure 3: Map of Bahía Blanca city.
About the area, he wrote: "I have several times alluded to the surface of the ground being incrusted with salt. This phenomenon is quite different from that of the salinas, and more extraordinary. In many parts of South America, wherever the climate is moderately dry, these incrustations occur; but I have nowhere seen them so abundant as near Bahia Blanca. The salt here, and in other parts of Patagonia, consists chiefly of sulphate of soda with some common salt. As long as the ground remains moist in the salitrales (as the Spaniards improperly call them, mistaking this substance for saltpeter), nothing is to be seen but an extensive plain composed of a black, muddy soil, supporting scattered tufts of succulent plants. On returning through one of these tracts, after a week's hot weather, one is surprised to see square miles of the plain white, as if from a slight fall of snow..." (Darwin 1845, p. 78). Bahía Blanca means White Bay. The name is due to the typical colour of the salt covering the soils surrounding its shores. "The wide expanse of water is choked up by numerous great mud-banks, which the inhabitants call Cangrejales, or crabberies, from the number of small crabs..." (Darwin 1845, p. 80). The bay (which is presently considered an estuary, Perillo et al. 2004) was seen by Ferdinand Magellan in 1520 during the first circumnavigation around the world on the orders of Charles I of Spain, as he searched for a pass connecting the Atlantic and Pacific oceans. Darwin's observations of the area are arranged according to the age of the sediments he described, from the oldest to the youngest.
THE GREAT PAMPEAN FORMATION
"The Beagle arrived here on the 24th of August, and a week afterwards sailed for the Plata. With Captain Fitz Roy's consent I was left behind, to travel by land to Buenos Ayres. I will here add some observations, which were made during this visit and on a previous occasion, when the Beagle was employed in surveying the harbour. The plain, at the distance of a few miles from the coast, belongs to the great Pampean formation, which consists in part of a reddish clay, and in part of a highly calcareous marly rock" (Darwin 1845, p. 81). This sedimentation, including both loess (sediments with a high content of volcanic derived particles) and loessoid sediments (reworked loess) (Zárate 2003), was related to a phase of late Miocene (ca. 10 Ma) orogeny in the Andes, which acted as a barrier to moisture-laden Pacific winds. This initiated ''the desertification of Patagonia caused by the rain shadow while precocious Pampa environment probably came into prominence at about this time'' (Patterson and Pascual 1972). Traverse (1982) refers to this 10-millionyear period as the "Ultimogene". Its inception is based on the presence of practically 100% extant plant genera. By this time, also began the establishment of steppe vegetation, dominated by grasses and by shrubby composites and chenopods in America and Eurasia. In the southern Pampas (Argentina), at least the last 9-3 Ma record is described as sequences composed of alternating loess and palaeosol units known in general as Pampean sediments (Fidalgo et al. 1975) and locally as the Saldungaray, La Toma (Furque 1967), and Monte Hermoso (Zavala 1993) formations. These sequences, stretch across South America from 23ºS to 41ºS (Teruggi 1957, Bargo and Deschamps 1996). They indicate climatic fluctuations, alternating between arid and cold (loess deposition) and warm and humid (palaeosol development) intervals. Further interpretation of these sediments and the related tosca beds are subject of another article in this volume (Zárate and Folguera 2009). The stratigraphic correlation among exposures of the Pampean sediments is quite difficult because of the lithologic uniformity, but they bear faunas of different ages (from the late Miocene to the Middle Pleistocene) which greatly help in this correlation. The oldest exposures of these Cenozoic sequences in the area are those of Grünbein and Barrancas de Sarmiento near Bahía Blanca city (Fig. 2), including loessoid sediments devoid of pollen, but yielding vertebrate remains. The fauna-especially the octodontoid rodents of the genera Xenodontomys and Phtoramys-suggests a late Miocene age (Verzi and Deschamps 1996, Deschamps et al. 1998, Verzi et al. 2008). Other loessoid sediments exposed at Las Obscuras (Fig. 2), in the middle basin of Río Sauce Grande, were also sterile in pollen, but the mammal remains - especially the octodontoid rodent Actenomys priscus - suggest an early Pliocene age. On this basis, Las Obscuras Formation may be correlated with the base of the Monte Hermoso Formation - Montehermosan Stage/ Age - exposed at Farola Monte Hermoso (Fig. 2) on the Atlantic coast.
MONTE HERMOSO: THE RED EARTH CLIFFS NEAR PUNTA ALTA
On October 19, 1832 he visited Monte Hermoso for the first time: "The Captain landed for half an hour at Monte Hermoso, (or Starvation point as we call it) to take observations" (Keynes 2001). "At the distance of about thirty miles from Punta Alta, in a cliff of red earth, I found several fragments of bones, some of large size. Among them were the teeth of a gnawer, equalling in size and closely resembling those of the Capybara (now Phugatherium cataclisticum = Chapalmatherium perturbidum, see Vucetich et al. 2005), whose habits have been described; and therefore, probably, an aquatic animal. There was also part of the head of a Ctenomys; the species being different from the Tucutuco, but with a close general resemblance (now Actenomys priscus, Deschamps 2005). The red earth, like that of the Pampas, in which these remains were embedded, ...." (Darwin 1845, p. 82). (see also Fernicola et al. 2009). These red earth cliffs, today known as Farola Monte Hermoso (Fig. 4) to distinguish it from Monte Hermoso Beach, were described by Darwin with more detail in 1846: "At Monte Hermoso there is a good section, about one hundred feet in height, of four distinct strata, appearing to the eye horizontal, but thickening a little towards the N.W. The uppermost bed, about twenty feet in thickness, consists of obliquely laminated, soft sandstone, including many pebbles of quartz, and falling at the surface into loose sand. The second bed, only six inches thick, is a hard, dark-coloured sandstone. The third bed is pale-coloured Pampean mud; and the fourth is of the same nature, but darker coloured, including in its lower part horizontal layers and lines of concretions of not very compact pinkish tosca-rock." Zavala (1993) and Zavala and Navarro (1993), based on facies analysis, gave a new interpretation of the stratigraphy and paleoenvironments of this locality, worldwide known by its fossil content since the visit of Ameghino (1887), being the basis for the Montehermosan Age (see Pascual et al. 1966). They formally recognized three litostratigraphic units from bottom to top, the Monte Hermoso, Puerto Belgrano and Punta Tejada formations (the first unit including the two lower strata described by Darwin as of Pampean mud). Darwin argued about the origin of these strata at Monte Hermoso: "...were the strata at Monte Hermoso depositing at the bottom of a great open sea, between 800 and 1000 feet in depth? I much doubt this; for if so, the almost perfect carcasses of the several small rodents, the remains of which are so very numerous in so limited a space, must have been drifted to this spot from the distance of many hundred miles. It appears to me far more probable, that during the Pampean period this whole area had commenced slowly rising ... As the land continued to rise, it appears that this source of sediment was cut off; and in its place sand and pebbles were borne down by stronger currents, and conformably deposited over the Pampean strata".

Figure 4: The red earth cliffs near Punta Alta: Farola Monte Hermoso.
Zavala and Navarro (1993) interpreted that the Monte Hermoso Formation (including the Hermosense típico, the Limolitas Estratificadas Member and Limolitas Claras Member of Bonaparte 1960) was deposited through a fluvial dynamics of high sinuousity muddy finegrained rivers, with associated floodplain and meandering channel facies. The discontinuity observed between the lower section (Hermosense típico) and the upper section (Limolitas Estratificadas and Limolitas Claras members) is here interpreted as an internal erosional surface developed during the migration of a highly sinuous fluvial channel over flood plain deposits. The Puerto Belgrano Formation (arenas estratificadas of Bonaparte 1960) is interpreted as accumulated in an aeolian environment with facies of low relief dunes, and associated dry interdunes. The cliff is topped by the Punta Tejada Formation, formed by sands and conglomerates with quartzite pebbles accumulated in wadi and fluvial environments. Several points of this interpretation agree with that of Darwin: "It appears to me far more probable, that during the Pampean period this whole area had commenced slowly rising ..., and that tracts of land had thus been formed of Pampean sediment round the Ventana and the other primary ranges, on which the several rodents and other quadrupeds lived, and that a stream (in which perhaps the extinct aquatic Hydrochoerus lived)... As the land continued to rise, it appears that this source of sediment was cut off; and in its place sand and pebbles were borne down by stronger currents, and conformably deposited over the Pampean strata" (Darwin 1846, p. 82). The record of extinct aquatic capybaras as evidence for the presence of water-related sediments was analyzed in Deschamps et al. (2007). By this time grasslands were dominant at high latitudes. They developed during a global cooling event and marine transgression (Janis 1993), this latter probably locally responsible for the cliffs of Grünbein and Barrancas de Sarmiento (Zavala and Quattrocchio 2001). For the period between 5 and 3 Ma, Lambeck et al. (2002) proposed global warm conditions in view of low values of δ18O in foraminifera of marine sediments, and low range sea level oscillations. The environments might have been similar to the modern Chaqueña Phytogeographic Province with open xerophytic woodlands, but more humid, with seasonal differences in rainfall (Pascual and Ortiz Jaureguizar 1990).
THE LATE PLIOCENEEARLY PLEISTOCENE
"Even the view was insignificant;-a plain like the sea, but without its beautiful colour and defined outline." Sea level changes produced different geomorphic features in these homogeneous sediments, including erosion, differences in the equilibrium river profiles, regressive and transgressive events, and building of shell terraces during the regressive phases. During the late Pliocene-Early Pleistocene interval this area was probably affected by erosion processes (and no deposition; see Fig. 9). In the study area, a single late Pliocene event has been identified in the conglomerates of the La Delta Sequence within the valley of Río Sauce Grande at Dique Paso Piedras (Deschamps 2005). The complex terraces in the Río Sauce Grande (see below) were interpreted within an evolutionary model of valleys in arid-semiarid regions (Zavala and Quattrocchio 2001). In this model, valleys were active only sporadically and most of the time behaved as geomorphologically depressed zones, hosting locally sourced gravitational and aeolian deposits. Geologic and sedimentological data suggest that La Delta Sequence could represent the second stage of initial filling within a transport zone, or zone 2 of the evolution of fluvial valleys in arid and semiarid zones. This process is related to the beginning of a transgressive cycle of an interglacial period. Semidesertic environments that began their development during the global climatic deterioration of the late Miocene were dominant during the Pliocene in western Argentina. The final rise of the Sierras Pampeanas acted as a wind shadow, increasing the desertification of the western areas (Pascual 1984). The caviomorph rodent fauna in the southeastern coast of the Buenos Aires Province, suggests a strong arid pulse at the end of the late Pliocene that could be coeval with the global climatic deterioration at the Gauss-Matuyama boundary (Verzi 2001, Verzi and Quintana 2005).
THE LATE PLEISTOCENE: PUNTA ALTA AND THE MONSTERS OF EXTINCT RACES
The Pleistocene-Holocene is recorded in the coastal area as well as in the valleys of the main rivers of the area. One of the most renowned Pleistocene sites visited by Darwin is Punta Alta. He spent here several days looking for fossils in September and October, 1832 and August 1833: "We passed the night in Punta Alta, and I employed myself in searching for fossil bones; this point being a perfect catacomb for monsters of extinct races." (Darwin 1845, p. 80). Vertebrate fossils are referred to in another chapter of this volume (Fernicola et al. 2009). This area is today buried beneath the naval base of Puerto Belgrano and could not be revisited; he reported the description and a drawing of the profile (Darwin 1846, p. 82-83): "Punta Alta is situated about thirty miles higher up on the northern side of this same bay: it consists of a small plain, between twenty and thirty feet in height, cut off on the shore by a line of low cliffs about a mile in length, represented in the diagram with its vertical scale necessarily exaggerated (Fig. 5). He described four beds (AD). "The lower bed (A) ... stratified gravel or conglomerate ... curvilinear, owing to the action of currents, and dip in different directions; they include an extraordinary number of bones of gigantic mammifers and many shells... The second bed (B) is about fifteen feet in thickness ... of red, tough clayey mud, with minute linear cavities... The bed (C) is of stratified gravel, like the lowest one... These three lower beds are covered by an unconformable mantle (D) of stratified sandy earth, including many pebbles of quartz, pumice and phonolite, land and seashells."
 Figure 5: The original drawing of Punta Alta section.
Figure 5: The original drawing of Punta Alta section.
The mammal bones found in this section "... were associated with twenty-three species of shells, of which thirteen are recent and four others very closely related to recent forms; whether the remaining ones are extinct or simply unknown, must be doubtful, as few collections of shells have been made on this coast. As, however, the recent species were embedded in nearly the same proportional numbers with those now living in the bay, I think there can be little doubt, that this accumulation belongs to a very late tertiary period" (Darwin 1845, p. 83). The 23 mentioned species are in fact 20 mollusk species, the other three are barnacles (Balanus), anthozoan corals (Astraea) and bryozoans (Flustra), all of them pertaining to the middle Holocene transgression recorded in the Bahía Blanca estuary (Farinati personal communication). Darwin made extensive observations on one pennatulaceous (Coelenterata, Anthozoa) which he named Virgularia patagonica, but known today as Stylatula darwini (Farinati 1989): At low water hundreds of these zoophytes might be seen, projecting like stubble, with the truncate end upwards, a few inches above the surface of the muddy sand" (Darwin 1845, p. 99). Especially noteworthy in this area, although not seen by Darwin, is the record of fossil trackways discovered by researchers of the UNS some kilometres east of Punta Alta (Fig. 2), in the paleoichnological site of Pehuén-Có (Fig. 6a). These trackways are quite rare in the fossil record, particularly in such amounts, and have been visited by researches of many foreign countries. They belong to birds, xenarthrans, camelids, macraucheniids, and were dated in 12,000 + 100 years 14C BP. They have been extensively studied by Drs. Manera and Aramayo (Aramayo and Manera de Bianco 1996, Quattrocchio and Borromei 1998). (Fig. 6a) Another outstanding record in this area is that of early aborigines' footprints (Fig. 6b) which were found at the Monte Hermoso1 site and were dated in 7,000 years BP (Bayón and Politis 1996, Zavala et al. 1992).

 Figure 6: a) Trackway of Megatherium on Pleistocene sediments of the Pehuén-Có ichnological site; b) Human footprint on Holocene sediments of the Monte Her-moso I archaeological site.
Figure 6: a) Trackway of Megatherium on Pleistocene sediments of the Pehuén-Có ichnological site; b) Human footprint on Holocene sediments of the Monte Her-moso I archaeological site.
THE RIO SAUCE
The Late Pleistocene-Holocene is also recorded in the valleys of the main rivers which flow across Tertiary units. The Río Sauce Grande is the main river in the southwestern Pampas, and Darwin crossed it ridding to Buenos Aires on September 8th. 1833: "I hired a Gaucho to accompany me on my ride to Buenos Ayres... The distance to Buenos Ayres is about four hundred miles, and nearly the whole way through an uninhabited country. We started early in the morning; ascending a few hundred feet from the basin of green turf on which Bahia Blanca stands, we entered on a wide desolate plain... After a long gallop, having changed horses twice, we reached the Rio Sauce: it is a deep, rapid, little stream, not above twenty-five feet wide. The second posta on the road to Buenos Ayres stands on its banks; ..." (Darwin 1845, p. 106; Figs. 7 and 8).
 Figure 7: The Río Sauce Grande and Sierra de la Ventana, with aborigines as drawn by E. Lassalle in 1827 during the voyage with the French naturalist Alcide d'Orbigny.
Figure 7: The Río Sauce Grande and Sierra de la Ventana, with aborigines as drawn by E. Lassalle in 1827 during the voyage with the French naturalist Alcide d'Orbigny.
 Figure 8: The Río Sauce Grande today.
Figure 8: The Río Sauce Grande today.
Remains of three levels of ancient terraces assigned to La Delta (see above), San José and Agua Blanca sequences (dated as Late Pliocene, Middle Pleistocene and Late Pleistocene-Holocene respectively; Zavala and Quattrocchio 2001, Deschamps 2005), document different episodes of incision and valley infilling. The Middle Pleistocene was particularly studied at Bajo San José, represented by the Bajo San José Sequence, which was deposited by a braided river, typical of arid to semiarid environments. The characteristic longitudinal bars and channels infilling would have provided varied niches for the rich fauna found in these deposits (Deschamps and Borromei 1992, Deschamps 2003, 2005). No pollen was found, but mammals, especially the octodontoid rodent Ctenomys kraglievichi as well as murid rodents and Tayassuidae, suggest the strongest warm pulse so far recognized in the Middle Pleistocene (Bonaerian age) of southern South America (Verzi et al. 2004).
THE VEGETATION AT THAT PERIOD
Darwin wondered about the vegetation coexisting with the large Late Pleistocene mammals of Punta Alta: "What, it may naturally be asked, was the character of the vegetation at that period; was the country as wretchedly sterile as it now is? As so many of the coembedded shells are the same with those now living in the bay, I was at first inclined to think that the former vegetation was probably similar to the existing one; but this would have been an erroneous inference, for some of these same shells live on the luxuriant coast of Brazil... Nevertheless, from the following considerations, I do not believe that the simple fact of many gigantic quadrupeds having lived on the plains round Bahia Blanca, is any sure guide that they formerly were clothed with a luxuriant vegetation: I have no doubt that the sterile country a little southward, near the Rio Negro, with its scattered thorny trees, would support many and large quadrupeds" (Darwin 1845, p. 84-85). This latter issue is developed in another article of this volume (Vizcaíno et al. 2009), but pollen and ostracod analyses performed at several localities of the Río Sauce Grande (e.g. Puesto La Florida, Fig. 2), and Arroyo Napostá Grande, including one site at the outlet near Punta Alta (Bertels and Martínez 1990, Borromei 1992, 1995, 1998, Grill 1993, 1995, 1997, Martínez 2002) provided information about the vegetation and climate. These analyses suggest arid conditions associated with more continental environments and related to lower sea level in the Atlantic coast of Buenos Aires province (Quattrocchio et al. 1995, 1998), quite different from "the luxuriant vegetation" and close to a "sterile country" of Darwin's comments. The pollen assemblage is dominated by Chenopodiaceae-Amaranthaceae, Poaceae and Asteraceae. Pollen grains that reflect the shrub and thorn scrub plant communities include among others the families Rhamnaceae (Condalia microphylla, or piquillín), Papilionaceae (Geoffroeae decorticans, or chañar) and Mimosaceae (Prosopis or algarrobo). The pollen assemblage represents the modern halophytic steppe and psammophytic herbaceous steppe vegetation, along with shrubby woodland vegetation. Sporadic high abundances of Cruciferae probably reflect habitats subject to natural causes of disturbance such as intense eolian action under an arid climate (from Quattrocchio and Borromei 1998).
THE RECENT PAST
The Late Pleistocene/Holocene transition is characterized by development of palaeosols. During the early Holocene sea level was still lower than today; pollen records reflect the development of a vegetation community characteristic of coastal dunes. Temperature and humidity reached its maximum during the mid-Holocene when the high diversity and abundance of marine dinocysts and acritarchs indicate a transgression. This event is associated with the gramineous steppe in the continent reflecting more temperate or local humid conditions. The relative sea level rise led to the flooding of the riverbeds, producing deposition of grey muddy facies. Approximately at 3000 years BP the marine influence ended in the area. After 2610 years BP, the development of a psammophytic herbaceous steppe suggests arid to semiarid conditions, followed by an interval of higher humidity inferred at approximately 2000 years BP, through the development of gramineous steppe communities. A relative rise of temperature may be inferred by the southward expansion of the Brazilian mammal fauna. These multidisciplinary research results are presented in a chronosequence chart synthesizing the suggested space-time correlation of the events recognized (Fig. 9). In this chart, some of the studied localities-covering the whole time interval represented in the area- were arranged in a hypothetical E-W line crossing the Río Sauce Grande basin and the highlands. In part, this line is also approximately the one followed by Darwin when riding from Bahía Blanca to Tapalqué (Tapalguen) as he crossed the region toward the Río Sauce and stopped at the second posta.
 Figure 9: Chronostratigraphic chart showing space-time correlation of the geologic events in the area. ABS. Agua Blanca Sequence; ChLBFm., Chacra La Blanqueada Formation; LDS, La Delta Sequence; MH-S-LT fms., Monte Hermoso-Saldungaray-La Toma formations; MSFm., Matadero Saldungaray Formation; SJS, San José Sequence. Thick black lines indicate the levels exposed at each locality.
Figure 9: Chronostratigraphic chart showing space-time correlation of the geologic events in the area. ABS. Agua Blanca Sequence; ChLBFm., Chacra La Blanqueada Formation; LDS, La Delta Sequence; MH-S-LT fms., Monte Hermoso-Saldungaray-La Toma formations; MSFm., Matadero Saldungaray Formation; SJS, San José Sequence. Thick black lines indicate the levels exposed at each locality.
ACKNOWLEDGEMENTS
The authors thank Dr. Beatriz Aguirre-Urreta and the Asociación Geológica Argentina for the invitation to participate in this volume, and the reviewers Drs. M.G. Vucetich and M. Toledo for their fruitful comments which certainly improved the manuscript. Dr. E. Farinati kindly provided information about Pleistocene shells.
WORKS CITED IN THE TEXT
1. Ameghino, F. 1887. Monte Hermoso. Diario "La Nación", March 10: 1-10, Buenos Aires.
2. Aramayo, S.A. and Manera de Bianco, T. 1996. Edad y nuevos hallazgos de icnitas de mamíferos y aves en el yacimiento paleoicnológico de Pehuén-Có (Pleistoceno tardío), provincia de Buenos Aires, Argentina. 1º Reunión Argentina de Icnología, Asociación Paleontológica Argentina, Publicación Especial 4: 47-57.
3. Bargo, M.S. and Deschamps, C.M. 1996. El registro de Mylodon Owen, 1840 (Mammalia, Tardigrada) en el Pleistoceno del sur de la provincia de Buenos Aires. Comentarios sobre la distribución de los "sedimentos Pampeanos". Ameghiniana 33: 343-348.
4. Bayón, C. and Politis, G. 1996. Estado actual de las investigaciones en el sitio Monte Hermoso 1 (Prov. de Buenos Aires). Instituto de Ciencias Antropológicas, Facultad de Filosofia y Letras, Universidad de Buenos Aires, Arqueología 6: 83-116.
5. Bertels, A. and Martínez, D. 1990. Fresh water and transitional environments ostracodes of the Buenos Aires Province, Argentina. European Ostracodologists Meeting (EOM 1989) 123: 141-159.
6. Bonaparte, J. 1960. La sucesión estratigráfica de Monte Hermoso (Provincia de Buenos Aires). Acta Geológica Lilloana 3: 272-290.
7. Borromei, A.M. 1992. Geología y palinología de los depósitos cuaternarios en el valle del río Sauce Grande, provincia de Buenos Aires, Argentina. Unpublished PhD Dissertation, Universidad Nacional del Sur, Bahía Blanca. 200 p.
8. Borromei, A.M. 1995. Palinología, estratigrafía y paleoambientes del Pleistoceno tardío-Holoceno en el valle del río Sauce Grande, provincia de Buenos Aires, Argentina. Polen 7: 19-31.
9. Borromei, A.M. 1998. Vegetación y clima del Cuaternario tardío en el valle superior del Río Sauce Grande, Provincia de Buenos Aires, Argentina. Polen 9: 5-15.
10. Darwin, C. 1845. Journal of researches into the natural history and geology of countries visited during the voyage of HMS Beagle round the world, under the command of Capt. Fitz Roy, R.N. Second Edition, corrected, with additions. John Murray, 519 p., London.
11. Darwin, C. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co. 280 p., London.
12. Deschamps, C.M. 2003. Estratigrafía y paleoambientes del Cenozoico en el sur de la Provincia de Buenos Aires. El aporte de los vertebrados. Unpublished PhD Dissertation, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata. 317 p.
13. Deschamps, C.M. 2005. Late Cenozoic mammal bio-chronostratigraphy in southwestern Buenos Aires Province, Argentina. Ameghiniana 42: 733-750.
14. Deschamps, C.M. and Borromei, A.M. 1992. La fauna de vertebrados pleistocénicos del Bajo San José (provincia de Buenos Aires, Argentina). Aspectos paleoambientales. Ameghiniana 29: 177-183.
15. Deschamps, C.M., Tonni, E.P., Verzi, D.H., Scillato Yané, G.J., Zavala, C.A., Carlini, A.A., and Di Martino, V. 1998. Bioestratigrafía del Cenozoico superior continental en el área de Bahía Blanca, provincia de Buenos Aires. 5° Jornadas Geológicas y Geofísicas Bonaerenses, Actas 1: 49-57.
16. Deschamps, C.M., Olivares, A.I., Vieytes, E.C. and Vucetich, M.G. 2007. Ontogeny and diversity of the oldest capybaras (Rodentia: Hydrochoeridae; late Miocene of Argentina). Journal of Vertebrate Paleontology 27: 683-692.
17. Dozo, A.L. and Ginóbili, M.E. 1999. Bahía Blanca en imágenes 1828-1929. Edi UNs (Editorial de la Universidad Nacional del Sur, 326 p., Bahía Blanca.
18. Farinati E. 1989. Pennatuláceos (Coelenterata, Anthozoa) en sedimentos marinos holocenos de Bahía Blanca, Argentina. Ameghiniana 26: 229-231.
19. Fernicola, J.C., Vizcaíno, S.F. and De Iuliis, G., 2009. Historical review of the fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. Revista de la Asociación Geológica Argentina 64(1): 147-159.
20. Fidalgo, F., De Francesco, F. and Pascual, R. 1975. Geología superficial de la llanura bonaerense (Argentina). In Geología de la Provincia de Buenos Aires. 6° Congreso Geológico Argentino, Relatorio: 103-138, Buenos Aires.
21. Furque, G. 1967. Estratigrafía de la Región de Pillahuincó. Acta Geológica Lilloana 9: 79-114.
22. Grill, S.C. 1993. Estratigrafía y paleoambientes del Cuaternario en base a palinomorfos en la cuenca del arroyo Napostá Grande, provincia de Buenos Aires. PhD Dissertation, Universidad Nacional del Sur, (unpublished), 145 p., Bahía Blanca.
23. Grill, S.C. 1995. Análisis palinológico de un perfil cuaternario en la cuenca del arroyo Napostá Grande, Localidad García del Río, provincia de Buenos Aires. 4° Jornadas Geológicas Bonaerenses (Junín), Actas 1: 99-107.
25. Grill, S.C. 1997. Análisis palinológico de un perfil cuaternario en la cuenca media del arroyo Napostá Grande, Localidad García del Río, provincia de Buenos Aires, Argentina. Implicancias paleoambientales. Polen 8: 23-40.
26. Janis, C.M. 1993. Tertiary mammal evolution in the context of changing climates, vegetation, and tectonic events. Annual Review of Ecology and Systematics 24: 467-500.
27. Keynes, R.D. ed. 2001. Charles Darwin's Beagle Diary. Cambridge University Press. 464 p., Cambridge.
28. Lambeck, K., Esat, T.M.and Potter, E.-K. 2002. Links between climate and sea levels for the past three million years. Nature 419: 199-206.
29. Mallea, E. 1941. Todo verdor perecerá. Espasa Calpe Argentina, 222p., Buenos Aires.
30. Martínez, D. 2002. Microfauna (Ostracoda, Crustacea) del Cuaternario de las cercanías de Bahía Blanca, República Argentina. PhD Dissertation, Universidad Nacional del Sur, (unpublished), 378 p., Bahía Blanca.
31. Pascual, R. 1984. Late Tertiary mammals of southern South America as indicators of climatic deterioration. Quaternary of South America and Antarctic Peninsula 2: 1-30.
32. Pascual, R. and Ortiz Jaureguizar, E. 1990. Evolving climates and mammal faunas in Cenozoic South America. Journal of Human Evolution 19: 23-60.
33. Pascual, R., Ortega Hinojosa, E.J., Gondar, D. and Tonni, E.P. 1966. Paleontografía Bonaerense. Fascículo IV. Vertebrata. Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, 202 p., La Plata.
34. Patterson, B. and Pascual, R. 1972. The Fossil Mammal Fauna of South America. In Keast, A., Erk, F.C. and Glass, B. (eds.) Evolution, Mammals, and Southern Continents, State University of New York Press, 247-309, New York.
35. Perillo, G.M.E., Piccolo, M.C., Palma, E., Pierini, J.O. and Pérez, D.E. 2004. Oceanografía Física del estuario de Bahía Blanca. In Piccolo, M.C. and Hoffmeyer, M. (eds.) El Ecosistema del Estuario de Bahía Blanca. Instituto Argentino de Oceanografía, 61-67, Bahía Blanca.
36. Quattrocchio, M.E. and Borromei, A.M. 1998. Paleovegetational and paleoclimatic changes during the Late Quaternary in southwestern Buenos Aires province and southern Tierra del Fuego (Argentina). Palynology 22: 67-82.
37. Quattrocchio, M.E., Borromei, A.M. and Grill, S.C. 1995. Cambios vegetacionales y fluctuaciones paleoclimáticas durante el Pleistoceno Tardío-Holoceno en el sudoeste de la provincia de Buenos Aires (Argentina). 6° Congreso Argentino de Paleontología y Bioestratigrafía (Trelew), Actas: 221-229.
38. Quattrocchio, M.E., Grill, S.C. and Zavala, C.A. 1998. Chronostratigraphic and palynozone chronosequences charts of Napostá Grande Creek, southwestern Buenos Aires Province, Argentina. Quaternary of South America and Antarctic Peninsula 11: 111-133.
39. Quattrocchio, M.E., Borromei, A.M, Deschanps, C.M., Grill, S.C., and Zavala, C.A. 2008. Landscape evolution and climate changes in the Late Pleistocene - Holocene, southern Pampa (Argentina): Evidence from palynology, mammals and sedimentology. Quaternary International 181: 123-138.
40. Teruggi, M. 1957. The nature and origin of Argentine loess. Journal of Sedimentary Petrology 27: 322-332.
41. Traverse, A. 1982. Response of world vegetation to Neogene tectonic and climatic events. Alcheringa 6: 197-209.
42. Traverse, A. 2007. Paleopalynology. In Landman, N.H. and Jones, D.S. (eds.) Topics in Geobiology, Springer (Second Edition), 28 , 813 p., Dordrecht.
43. Verzi, D.H. 2001. Phylogenetic position of Abalosia and the evolution of the extant Octodontinae (Rodentia, Caviomorpha, Octodontidae). Acta Theriologica 46: 243-268.
44. Verzi, D.H. and Deschamps, C.M. 1996. Presencia de roedores ctenominos del género Xenodontomys en Loma Sarmiento, Provincia de Buenos Aires. 3º Jornadas Geológicas y Geofísicas Bonaerenses (1995), Actas 1: 47-54.
45. Verzi, D.H. and Quintana, C.A. 2005. The Caviomorph rodents from the San Andrés Formation, east-central Argentina, and global Late Pliocene climatic change. Palaeogeography, Palaeoclimatology, Palaeoecology 219: 303-320.
46. Verzi, D.H., Deschamps, C.M. and Tonni, E.P. 2004. Biostratigraphic and paleoclimatic meaning of the Middle Pleistocene South American rodent Ctenomys kraglievichi (Caviomorpha, Octodontidae). Palaeogeography, Palaeoclimatology, Palaeoecology, 212: 315-329.
47. Verzi, D.H., Montalvo, C.I. and Deschamps, C.M. 2008. Biostratigraphy and biochronology of the Late Miocene of central Argentina: evidence from rodents and taphonomy. Geobios 44: 145-155.
48. Vizcaíno, S.F., Fernicola, J.C. and Fariña, R.A., 2009. Young Darwin and the ecology of South American fossil mammals. Revista de la Asociación Geológica Argentina 64(1): 160-169.
49. Vucetich, M.G., Deschamps, C.M., Olivares, A.I. and Dozo, M.T. 2005. Capybaras, shape, size and time: a model kit. Acta Paleontologica Polonica 50: 259-272.
50. Zárate, M.A. 2003. Loess of southern South America. Quaternary Science Reviews 22: 1987-2006.
51. Zárate, M.A. and Folguera, A. 2009. On the formations of the Pampas in the footsteps of Darwin: south of the Salado. Revista de la Asociación Geológica Argentina 64: 124-136.
52. Zavala, C.A. 1993. Estratigrafía de la localidad de Farola Monte Hermoso (Plioceno-Reciente). Provincia de Buenos Aires. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza), Actas 2: 228-235.
53. Zavala, C.A. and Navarro, E. 1993. Depósitos fluviales en la Formación Monte Hermoso (Plioceno inferior-medio), provincia de Buenos Aires. 12° Congreso Geológico Argentino y 2° Congreso de Exploración de Hidrocarburos (Mendoza), Actas 2: 236-244.
54. Zavala, C.A. and Quattrocchio, M.E. 2001. Estratigrafía y evolución geológica del río Sauce Grande (Cuaternario), provincia de Buenos Aires. Revista de la Asociación Geológica Argentina 56: 25-37.
55. Zavala, C.A., Grill, S.C., Martínez, D., Ortíz, H.O. and González, R. 1992. Análisis paleoambiental de depósitos cuaternarios, Sitio paleoicnológico Monte Hermoso I, provincia de Buenos Aires. 3° Jornadas Geológicas Bonaerenses (La Plata), Actas: 31-37.
Juan Carlos Fernicola1, Sergio F. Vizcaíno2 and Gerardo De Iuliis3
1 Sección Paleontología de Vertebrados, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia". Buenos Aires, Argentina. Email: jctano@macn.gov.ar 2 División Paleontología Vertebrados, Museo de La Plata, La Plata, Argentina. CONICET. Email: vizcaino@fcnym.unlp.edu.ar 3 School of Nursing, George Brown College of Applied Arts and Technology, Toronto; and Department of Ecology and Evolutionary Biology, University of Toronto, Toronto, Canada. Email: gerry.deiuliis@utoronto.ca
ABSTRACT
During the first two years of his voyage aboard HMS Beagle, Charles Darwin collected a considerable number of fossil mammals from various localities in Argentina and Uruguay. Among these remains are those of large mammals that Darwin informally assigned to Megatherium and Mastodon, the only large taxa then known for South America, and of small and mediumsized mammals that Darwin recognized as representing at least two rodents and a horse. The study of Darwin's collection was entrusted to Richard Owen, who described eleven taxa between 1837 and 1845, including the six following ones: Toxodon platensis, Macrauchenia patachonica, Equus curvidens, Scelidotherium leptocephalum, Mylodon darwini and Glossotherium sp. This contribution provides a synthesis of Darwin's preliminary assignments and evaluates the reasons that led him to recognize only megatheres and mastodonts for the large fossil remains. Also, it discusses the current taxonomic status of the taxa described or erected by Owen between 1837 and 1845 and the influence that Owen's taxonomic and phylogenetic conclusions had on the development of Darwin's ideas on evolution.
Keywords: Darwin; Taxonomy; South America; Fossil mammals.
RESUMEN: Los mamíferos fósiles colectados por Charles Darwin en América del Sur durante su viaje a bordo del HMS Beagle. Durante los dos primeros años de su viaje a bordo del HMS Beagle, Charles Darwin colectó en distintas localidades de Argentina y Uruguay un considerable número de mamíferos fósiles. Entre estos se cuentan los grandes mamíferos que informalmente Darwin asignó a Megatherium y Mastodon, únicos grandes taxones conocidos hasta ese momento para América del sur y entre los pequeños y medianos mamíferos reconoció la presencia de al menos dos tipos de roedores y un caballo. El estudio posterior de todos los ejemplares colectados por Darwin fue llevado a cabo Richard Owen, quien entre 1837 y 1845 describió once taxones, entre los cuales, seis eran nuevos taxones: Toxodon platensis, Macrauchenia patachonica, Equus curvidens, Scelidotherium leptocephalum, Mylodon darwini y Glossotherium sp. En esta contribución se brinda una síntesis de las asignaciones preliminares efectuadas por Charles Darwin y se evalúan los motivos que lo habrían llevado a reconocer sólo megaterios y mastodontes entre los grandes mamíferos fósiles por él colectados. También tiempo, se discute el estatus taxonómico de los taxones descritos o fundados por Richard Owen entre 1837 y 1845 y la influencia que habrían tenido las conclusiones taxonómicas y filogenéticas de Owen en el desarrollo de las ideas evolutivas de Darwin.
Palabras clave: Darwin; Taxonomía; América del Sur; Mamíferos fósiles.
INTRODUCTION
Charles Darwin collected fossil mammals from various South American localities during his voyage aboard HMS Beagle. He recovered his first fossil at Punta Alta (Buenos Aires Province, Argentina) on September 23, 1832, and continued collecting intermittently at this locality until October 16. Three days later he collected several specimens from Monte Hermoso (Buenos Aires Province, Argentina). Darwin returned to Punta Alta between August 29 - 31, 1833. Several weeks later, around September 19, he collected in Guardia del Monte (Buenos Aires Province); the Rio Carcarañá or Tercero (Santa Fe Province, Argentina) on October 1; and the Bajada Santa Fe (Paraná, Entre Ríos Province, Argentina) on October 10. Moving to Uruguay, he collected fossils during November 25 - 26 from the Arroyo Sarandí near the city of Mercedes (Soriano Department). He returned to Argentina and collected his last specimens at Puerto San Julián (Santa Cruz Province) during the first several months of 1834 (see Fig. 1). All the specimens, sent to his mentor John Stevens Henslow and later deposited in the Royal College of Surgeons (London, England), were studied by Richard Owen beginning in 1836.
 Figure 1: Map of the localities where Charles Darwin collected fossil mammals.
Figure 1: Map of the localities where Charles Darwin collected fossil mammals.
Regrettably, the intense bombardment suffered by this institution during April 10 and 11, 1941, destroyed a large part of its paleontological treasures. Cave's (1942) catalogue of the surviving specimens revealed that some 95% of the collection had been lost - of 5,000 specimens, only 175 remained. Among these, fortunately, are various of the specimens collected by Charles Darwin and described by Richard Owen (1837-1845) as new taxa, including Equus curvidens, Glossotherium sp; Macrauchenia patachonica, Mylodon darwini, Scelidotherium leptocephalum and Toxodon platensis (Cave 1942). Beginning in 1946, nearly all of Darwin's collection was transferred to the Natural History Museum (London), where it is still housed (see appendix 1). The Royal College of Surgeons retained only a single specimen, the remains of the Megatherium americanum specimen that apparently allowed Owen (1840) to recognize the presence of a fifth upper molariform in this taxon (see appendix 1). The aim of this contribution is to provide an update on the taxonomic status of the various taxa recognized or established by Owen (1837-45), and to consider the influence the specimens and their assignment had on the development of Darwin's ideas.
HISTORICAL BACKGROUND
The first notices of South American "fossils" appear in the reports of early Spanish explorers to South America and in the earliest histories of America (e.g. Cieza de León 1553, Falkner 1774). These objects were interpreted as the remains of an ancient race of giant humans that inhabited this part of the world before being smitten by divine force. Cieza de León (1553, p. 183) provided the following vivid tale about huge bones found in Santa Elena, Ecuador: "Y así, dicen que, estando todos juntos envueltos en su maldita sodomía, vino fuego del cielo temeroso y muy espantable, haciendo gran ruido, del medio del cual salió un ángel resplandeciente, con una espada tajante y muy refulgente, con la cual de un solo golpe los mató a todos y el fuego los consumió, que no quedó sino algunos huesos y calaveras, que para memoria del castigo quiso Dios que quedasen sin ser consumidas por el fuego. Esto dicen de los gigantes; lo cual creemos que pasó, porque en esta parte que dicen se han hallado y se hallan huesos grandísimos. Y yo he oído a españoles que han visto pedazo de muela que juzgaban que a estar entera pesara más de media libra carnicera, y también que habían visto otro pedazo del hueso de una canilla, que es cosa admirable contar cuán grande era, lo cual hace testigo haber pasado…". This sort of general belief persisted, in South America at least, until the end of the 18th century, even though some of the remains were recognized as nonhuman, as indicated in the following passages by the English Jesuit Thomas Falkner (1774, p. 54-55) "On the banks of the River Carcarania, or Tercero, about three or four leagues before it enters into the Parana, are found great numbers of bones, of an extraordinary bigness, which seem human. There are some greater and some less, as if they were of persons of different ages. I have seen thigh-bones, ribs, breast-bones, and pieces of skulls. I have also seen teeth, and particularly some grinders which were three inches in diameter at the base. These bones (as I have been informed) are likewise found on the banks of the Rivers Parana and Paraguay, as likewise in Peru. The Indian Historian, Garcilasso de la Vega Inga, makes mention of these bones in Peru, and tells us that the Indians have a tradition, that giants formerly inhabited those countries, and were destroyed by God for the crime of sodomy. I myself found the shell of an animal, composed of little hexagonal bones, each bone an inch in diameter at least; and the shell was near three yards over. It seemed in all respects, except it's [sic] size, to be the upper part of the shell of the armadillo; which, in these times, is not above a span in breadth." It was during this period that the illustrious French naturalist George Cuvier (1796) published the first scientific work on a South American fossil, which he described and named Megatherium americanum based on the specimen recovered by Fray Manuel Torres in 1787 from Luján in Buenos Aires Province, Argentina and sent the following year by the Marquis of Loreto, Viceroy of Río de la Plata, to the Real Gabinete de Historia Natural in Madrid, Spain. Mones (1986) considered the erection of this species as the first Linnean nomenclatorial act for a South American fossil species. This specimen had, however, previously been described and figured by Juan Bautista Bru de Ramón, an artist and first taxidermist of the Real Gabinete de Historia Natural. In this regard, Garriga (1796, as quoted in López Piñeiro and Glick 1993, p. 126) noted that "… restaba superar la dificultad de que permitiese Bru que se diese a luz una obra que él había tenido en otro tiempo intento de publicar, y por varias circunstancias imprevistas sepultó en el olvido." López Piñeiro and Glick (1993, p. 66) maintained that the preparation of the monograph "… no fue entonces editada, pero su preparación debía estar tan adelantada que un tal Roume, representante del gobierno de francés en Santo Domingo, consiguió un juego de pruebas de las planchas a su paso por Madrid en 1793. Roume envió las pruebas de las planchas al Institut de France, del que era miembro correspondiente, acompañadas de una "corta descripción" del esqueleto. La sección de ciencias del Institut encargó a George Cuvier un informe sobre el tema, que apareció publicado en 1796…" Cuvier (1806) also studied fossil proboscidean remains found by Dombey in Peru and by Humboldt in various localities in Bolivia, Chile, Colombia, and Ecuador, among which he recognized three morphotypes, designated informally as "mastodonte a dents étroites", "mastodonte Cordillierès" and "mastodonte humboldien". Cuvier (1823) later formally named them Mastodon angustidens, Mastodon andium and Mastodon humboldti, respectively (see Cabrera 1929). In this paper, Cuvier (1823, p. 179) included a handwritten communication by "don Dámaso Antonio Larrañaga, a priest of Montevideo, to Mr. Augusto de Saint-Hilaire" that briefly described a group of fossil specimens, including a partial exoskeleton (or carapace), under the name "Dasypus (Megatherium Cuv)." Cuvier (1823) thus acknowledged that this carapace probably belonged to Megatherium. Whatever the reasons that prompted Larrañaga's subgeneric assignment (that is, in recognizing the existence of armored megatheres), the fact that his opinion appeared in a work by Cuvier ensured its credibility, so that several incomplete remains later sent to Europe were similarly recognized (e.g., as by Weiss 1830, Clift 1835). It was not until later that Owen (1839a) demonstrated that megatheres were not armored and that the remains belonged instead to the group of mammals he termed glyptodonts, a group closely related to armadillos. Knowledge of South American fossil mammals remained at this elementary level for much of the early part of the 19th century, as is made clear from Owen's (1838, p. 13) opening paragraph on his work on the fossil mammals in Darwin's The Zoology of the Voyage of H.M.S. Beagle: "It may be expected that the description of the osseous remains of extinct Mammalia, which rank amongst the most interesting results of Mr. Darwin's researches in South America, should be preceded by some account of the fossil mammiferous animals which have been previously discovered in that Continent. The results of such a retrospect are, however, necessarily comprised in a very brief statement; for the South American relics of extinct Mammalia, hitherto described, are limited, so far as I know, to three species of Mastodon, and the gigantic Megatherium." It was against this systematic framework that Darwin began his efforts in collection and preliminary taxonomic assignments that Owen (1837-1845) later modified. Several of his assignments were considered taxonomic errors or misinterpretations (e.g., Sulloway 1982, Keynes 2001), even though his inferences were in accordance with what was then known.
DARWIN'S FOSSIL MAMMALS
The following presents a synthesis of Owen's initial taxonomic assignments (1837-45) of the remains collected by Darwin in South America, including the preliminary assignments made by Darwin. The latter's assignments were gleaned from field notes (Darwin 2002-2008), personal correspondence (Darwin 1985) and Charles Darwin's Beagle Diary (in Keynes 2001), all material written by Darwin before 1836, the year of his return to England. The suprageneric classification used here follows McKenna and Bell (1997).
SYSTEMATIC PALEONTOLOGY
Grandorder Ungulata Linnaeus, 1776 Mirorder Meridiungulata McKenna, 1975 Order Notoungulata Roth, 1903 Suborder Toxodontia Owen, 1853 Family Toxodontidae Owen, 1845 Subfamily Toxodontinae Owen, 1845 Genus Toxodon Owen, 1837
Toxodon platensis Owen, 1837 Fig. 2
 Figure 2: Skull of Toxodon platensis in ventral view (from Owen 1838). Scale bar = 10 cm.
Figure 2: Skull of Toxodon platensis in ventral view (from Owen 1838). Scale bar = 10 cm.
The specimens assigned to this taxon by Owen (1837, 1838) were recovered by Darwin from various South American localities. On October 1 and 10, 1833, he collected at Río Carcaraña o Tercero (Santa Fe Province) and at Bajada Santa Fe (Paraná, Entre Ríos Province). His field notes (Darwin 2002-2008) describe a fossil specimen as "…a large rotten tooth & in the layer large cutting tooth", a description that agrees with those included in his Beagle Diary ("… a curious & large cutting tooth"; in Keynes 2001, p. 193) and written to John Stevens Henslow ("In ye R. Carcarana I got a tooth, which puzzles even my conjectures, it looks like an enormous gnawing one"; Letter 229 in Darwin 1985). Darwin's (2002-2008) field notes for November 26, 1833, state: "Started went round by a house to see large head & bones, washed out of Barranca & found after a flood. - pieces here also of Casca - Barranca" while his Beagle Diary (in Keynes 2001) records: "Began my return in a direct line to M. Video; went by an Estancia where there was a part, very perfect, of the head of a Megatherium. I purchased it for a few shillings". An alveolus of this specimen, recovered from Arroyo Sarandí, Departament of Soriano, Uruguay, easily accommodated a molariform tooth found by Darwin in Carcaraña (Owen 1838).
Toxodon sp.
The mandible and isolated molariforms assigned by Owen (1837, 1838) to this genus were collected between September 23 - October 16, 1832, at Punta Alta (Buenos Aires Province, Argentina). On the molariforms, Darwin (1985, Letter 192) noted that "some large molar teeth, which in some respects would seem to belong to an enormous Rodentia". Burmeister (1866) based the species Toxodon darwini on the mandible described by Owen (1838) Bond (1999) acknowledged the possible synonymy of Toxodon darwini with Toxodon platensis, although he recognized that a revision of the genus was required. Owen (1837, 1838, p. 16) recognized Toxodon as "A gigantic extinct mammiferous animal, referrible to the Order Pachydermata, but with affinities to the Rodentia, Edentata, and Herbivorous Cetacea". Darwin (1839, p. 180) summarized the morphological basis leading to this idea of multiple affinities: "Mr. Owen says, judging from the portion of the skeleton preserved, the Toxodon, as far as dental characters have weight, must be referred to the rodent order. But from that order it deviates in the relative position of its supernumerary incisors, in the number and direction of the curvature of its molars, and in some other respects. It again deviates, in several parts of its structure which Mr. Owen enumerated, both from the Rodentia, and the existing Pachydermata, and it manifests an affinity to the Dinotherium and the Cetaceous order. Mr. Owen, however, observed, that the development of the nasal cavity and the presence of frontal sinuses, renders it extremely improbable that the habits of he Toxodon were so exclusively aquatic as would result from the total absence of hinder extremities; and concludes, therefore, that it was a quadruped, and not a Cetacean; and that it manifested an additional step in the gradation of mammiferous forms leading from the Rodentia, through the Pachydermata to the Cetacea; a gradation of which the water-hog of South America (Hydrochærus capybara) already indicates the commencement amongst existing Rodentia, of which order it is interesting to observe this species is the largest, while at the same time it is peculiar to the continent in which the remains of the gigantic Toxodon were discovered". The genus Toxodon is now considered among the more derived native notoungulates of South America. The Notoungulata, a clade of the Mirorder Meridiungulata, shares an ancestry with North American condylarths (Cifelli 1993) In other words, Toxodon has no close phylogenetic relationships with the groups mentioned by Owen (1838).
Grandorder Ungulata Linnaeus, 1776 Mirorder Meridiungulata McKenna, 1975 Order Liptopterna Ameghino, 1889 Superfamily Macrauchenioidea Gervais, 1855 Family Macraucheniidae Gervais, 1855 Subfamily Macraucheniinae Gervais, 1855 Genus Macrauchenia Owen, 1838
Macrauchenia patachonica Owen, 1838 Fig. 3a
 Figure 3: a) Cervical vertebra of Macrauchenia patachonica in lateral and ventral views (from Owen 1838). Scale bar = 5 cm; b) Tooth of Equus sp. in occlusal and lateral views (from Owen 1840). Scale bar = 1 cm.
Figure 3: a) Cervical vertebra of Macrauchenia patachonica in lateral and ventral views (from Owen 1838). Scale bar = 5 cm; b) Tooth of Equus sp. in occlusal and lateral views (from Owen 1840). Scale bar = 1 cm.
In a letter from March, 1834, to John Stevens Henslow, Darwin (1985, Letter 238) recounted that "At Port St Julian I found some very perfect bones of some large animal, I fancy a Mastodon". On this specimen, the only one found by Darwin in Puerto San Julián, Owen (1838, p. 35) erected Macrauchenia patachonica, which he considered "A large extinct Mammiferous Animal, referrible to the Order Pachydermata; but with affinities to the Ruminantia, and especially to the Camelidae". The evolutionary significance of this purported kinship impressed Darwin (1839, p. 210): "The most important result of this discovery, is the confirmation of the law that existing animals have a close relation in form with extinct species. As the guanaco [Camelidae] is the characteristic quadruped of Patagonia, and the vicuna of the snow-clad summits of the Cordillera, so in bygone days, this gigantic species [Macrauchenia patachonica] of the same family must have been conspicuous on the southern plains". The genus Macrauchenia is now considered as among the more derived native South American litopterns. As with the Notoungulata, the Litopterna comprises a clade of the Mirorder Meridiungulata and is closely related to North American condylarthrans, rather than camelids, as erroneously proposed by Owen (1838).
Grandorder Ungulata Linnaeus, 1776 Mirorder Altungulata Protero and Schoch, 1989 Order Perissodactyla Owen, 1848 Suborder Hippomorpha Word, 1937 Family Equidae Gray, 1821 Genus Equus Linnaeus, 1758
Equus sp. Fig. 3b
The remains assigned to this genus represent the first fossil horses found in South America. The two molars from Argentina were recovered from Punta Alta (Buenos Aires Province) and Bajada Santa Fe (Entre Rios Province). The specimen from Bajada Santa Fe was referred to by Darwin as a horse (in Keynes 2001). We have not been able to find any mention of the other specimen in his field notes (Darwin 2002-2008), Beagle Diary (in Keynes 2001), or personal correspondence (Darwin 1985). Although Owen (1840) did not attempt specific assignation of these molars, he did note that the only difference between them and those of Equus caballus was the smaller size of the former. Owen (1845) erected Equus curvidens based on Darwin's specimens, a species which is considered synonymous with Equus (Amerhippus) neogeus Lund by Prado and Alberdi (1994).
Grandorder Ungulata Linnaeus, 1776 Order Uranotheria McKenna and Bell, 1987 Suborder Tethytheria McKenna, 1975 Infraorder Behemota McKenna and Bell, 1987 Parvorder Proboscidea Illiger, 1811 Superfamily Elaphantoidea Gray, 1821 Family Gomphotheriidae Hay, 1922 Subfamily Rhynchoteriinae Hay, 1922 Tribe Cuvieroniini Cabrera, 1929 Genus Mastodon Rafinesque, 1814
Mastodon sp.
The specimens assigned to this taxon, collected by Darwin at Bajada Santa Fe (Entre Ríos Province) and Río Carcarañá or Tercero (Santa Fe Province), were noted by Owen (1840, p. 180) as "…the fragments of the teeth and portions of the skeleton which reached England, are not sufficient to lead to a determination of the species; but sufficiently prove it to have been nearly allied, if not identical, with the Mastodon angustidens of Cuvier, and unquestionably distinct from the Mastodon giganteum of the United States". Currently Stegomastodon platensis is the only proboscidean species recognized from the areas yielding Darwin's specimens (Alberdi and Prado 1995).
Magnorder Xenarthra Cope, 1889 Order Pilosa Flower, 1883 Suborder Phyllophaga Owen, 1842 Infraorder Mylodonta McKenna and Bell, 1987 Superfamily Mylodontoidea Gill, 1872 Family Scelidotheriidae Ameghino, 1889 Subfamily Scelidotheriinae Ameghino, 1889 Genus Scelidotherium Owen, 1839b
Scelidotherium leptocephalum Owen, 1839b Fig. 4
 Figure 4: Partial skeleton of Scelidotherium leptocephalum in ventral view (from Owen, 1838b). Scale bar = 10 cm.
Figure 4: Partial skeleton of Scelidotherium leptocephalum in ventral view (from Owen, 1838b). Scale bar = 10 cm.
This taxon was based by Owen (1839b) on the only nearly complete skeleton found by Darwin at Punta Alta (Buenos Aires Province). We are unable to find any preliminary assignment of this specimen by Darwin, although Sulloway (1982, p. 353) noted that Darwin considered the specimen as "allied to the Rhinoceros". Darwin made this conjecture on September 23, 1832 (in Keynes 2001) in the following Beagle Diary entry: "… I walked on to Punta Alta to look after fossils; & to my great joy I found the head of some large animal, imbedded in a soft rock. … It took me nearly 3 hours to get it out: As far as I am able to judge, it is allied to the Rhinoceros. … I did not get it on board till some hours after it was dark". However, the specimen assigned by Owen (1839b) to Scelidotherium was collected by Darwin in Punta Alta in August, 1833, the time during which Darwin had returned to this locality (Keynes 2001, p. 178). Further confirmation of this date appears in a letter from Darwin (1985, Letter 188), dated September 20, 1833, to Caroline S. Darwin, in which he recounts that "I likewise at Bahia Blanca found some more bones more perfect than those I formerly found, indeed one is nearly an entire skeleton". As indicated below, Darwin's "allied to the Rhinoceros" comment may more likely have been a reference to one of the cranial remains that Owen (1840) later assigned to the genus Megatherium, particularly as Owen's generic assignations of the sloths are for the most part still considered correct.
Magnorder Xenarthra Cope, 1889 Order Pilosa Flower, 1883 Suborder Phyllophaga Owen, 1842 Infraorder Mylodonta McKenna and Bell, 1987 Superfamily Mylodontoidea Gill, 1872 Family Mylodontidae Gill, 1872 Subfamily Mylodontinae Gill, 1872 Genus Mylodon Owen, 1839b
Mylodon darwini Owen, 1839b Fig. 5a
 Figure 5: a) Mandible of Mylodon darwini in occlusal view (from Owen, 1839b). Scale bar = 10 cm; b) Cranial fragment of the skull of Glossotherium in lateral view (from Owen 1839b) Scale bar = 10 cm.
Figure 5: a) Mandible of Mylodon darwini in occlusal view (from Owen, 1839b). Scale bar = 10 cm; b) Cranial fragment of the skull of Glossotherium in lateral view (from Owen 1839b) Scale bar = 10 cm.
The specimen assigned to this genus by Owen (1839b, p. 69) consists of "…the lower jaw with the series of teeth entire on both sides: but the extremity of the symphysis, the coronoid and condyloid processes, and the angular process of the left ramus, are wanting". This specimen was recovered by Darwin at Punta Alta (Buenos Aires Province), but we were unable to determine whether it was collected during his first or second sojourn to this area. In a letter sent to John Stevens Henslow in November, 1832, Darwin (1985, Letter 192) noted a completed list of fossils collected in Bahia Blanca, among which he emphasized "… the upper jaw & head of some very large animal, with 4 square hollow molars….& the head greatly produced in front…. I at first thought it belonged either to the Megalonyx or Megatherium…. In confirmation, of this, in the same formation I found a large surface of the osseous polygonal plates, which ``late observations'' (what are they?) show belong to the Megatherium". A footnote on this page, presumably inserted by the editors, indicates that this specimen was "Described in Fossil Mammalia, p. 63-73, by Richard Owen, who identified it as belonging to a distinct subgenus of Megatheroid Edentata, to which he gave the name Mylodon darwinii". However, we believe this to be incorrect - the specimen is not the one that Owen (1839b) later assigned to Mylodon darwini, as Dar-win indicated that the specimen was an upper jaw and skull, rather than the mandible on which Mylodon darwini is based. Darwin's indication of the presence of four molariforms possibly led to the assumption (i.e., in the footnote) that the specimen belonged to Mylodon darwini, given that five upper molariforms are present in Megatherium. However, the earliest descriptions of Megatherium indicate four upper molariforms. As noted above, it was Owen (1840, p. 102) who recognized the presence of a fifth upper molariform: "Upon clearing away the matrix from the palatal and alveolar surface of one of the cranial fragments of the Megatherium in Mr. Darwin's collection, I was gratified by the detection of the crown of a fifth molar". The taxonomic history of Mylodon is among the more complex for the taxa erected by Owen (1838-1840), due mainly to the ambiguity in the type species. Owen (1839b) erected Mylodon for two species, Mylodon darwini and Mylodon harlani. The former species was based on a left dentary from Punta Alta (Buenos Aires Province), whereas the second was based on a cast of a mandible from North America that Harlan (1835) had assigned to Megalonyx laqueatus. Owen (1839b) was unclear about his choice of type species, given that in the title of his description of his new genus he referred to the fossil collected by Darwin, but in the text Owen noted that Mylodon darwini was the second species of the genus. This ambiguity would be trivial if the two species were congeneric, but they have been subsequently considered as belonging to different genera. Consequently, some later researchers, such as Reinhardt (1879), Lydekker (1887) and Brown (1903), considered Mylodon harlani as the type of the genus, whereas other, such as Leidy (1855), bestowed this status on Mylodon darwini. The issue was resolved by Kraglievich (1928) in favor of Mylodon darwini based on the following reasons:
1) the title of the generic description refers to the mandible collected by Darwin; 2) the original material available to Owen was the dentary from Punta Alta; 3) Owen (1842) recognized Mylodon harlani as the second species of the genus; 4) in his "Conspectus familiarum, generum et specierum", Owen (1842, p. 169) listed Mylodon darwini as the first species of the genus.
Kraglievich's (1928) logic has been accepted by the scientific community, so that Mylodon darwini is currently considered the type species of Mylodon, whereas Mylodon harlani is placed in the genus Paramylodon. The taxonomic solution provided by Kraglievich (1928) not only resolved the taxonomic problem with Mylodon, but also helped clarify the problematic nomenclature of the genus Glossotherium (see below).
Magnorder Xenarthra Cope, 1889 Order Pilosa Flower, 1883 Suborder Phyllophaga Owen, 1842 Infraorder Mylodonta McKenna and Bell, 1987 Superfamily Mylodontoidea Gill, 1872 Family Mylodontidae Gill, 1872 Subfamily Lestodontinae Ameghino, 1889 Tribe Glossotheriini McKenna, 1987 Genus Glossotherium Owen, 1839b
Glossotherium sp. Fig. 5b
This genus was erected by Owen (1839b) based on the posterior half of a skull recovered from Arroyo Sarandí (Soriano Departament, Uruguay). In his Beagle Diary, Darwin (in Keynes 2001) noted that "We heard of some giants bones, which as usual turned out to be those of the Megatherium - With much trouble extracted a few broken fragments". As with Mylodon darwini, the taxonomic history of Glossot-herium is complex. Owen (1842) erected the new species Mylodon robustus on remains incluiding a nearly complete skull; and assigned to Mylodon darwini (based on a dentary) the cranial fragment previously described by Owen (1839b) as Glossotherium. Reinhardt's (1879) detailed description of a fossil sloth skull and mandible from Pergamino (Buenos Aires Province) recognized, 1) that the mandible was very similar to that described as Mylodon darwini by Owen (1839b), 2) that its skull features were sufficiently distinct as to suggest generic separation from Mylodon robustus, and 3) that the cranial fragment originally assigned by Owen (1839b) to Glossotherium was closely allied generically to Mylodon robustus. Not recognizing Mylodon darwini as type of the genus, Reinhardt (1879) proposed the new genus Grypotherium, in which he placed the dentary assigned by Owen (1839b) to Mylodon darwini and the specimen from Pergamino as the species Grypotherium darwini; and recognized Mylodon robustus as the type species of Mylodon. Ameghino (1889) accepted the generic differences noted by Reinhardt (1879), but considered, as Owen (1842) had before, that the cranial fragment of Glossotherium and the dentary of Mylodon darwini belonged to the same species, and so included these specimens - as well as the skull and mandible from Pergamino - in Glossotherium darwini, given that in this scenario Glossotherium has priority over Grypotherium. Smith-Woodward's (1900) revision of Darwin's South American fossil sloth collection concluded that the cranial fragment originally assigned to Glossotherium was congeneric with the specimen assigned to Mylodon robustus by Owen (1842) and that the dentary of Mylodon darwini and the specimen from Pergamino, described by Reinhardt (1879), were conspecific. However, as Smith-Woodward did not recognize Mylodon darwini as type species of Mylodon, he resurrected Reinhardt's (1879) Grypotherium. Kraglievich (1928) modified the taxonomy of these taxa based on a detailed revision of the group. This author held that the root of the problem was the rejection of Mylodon darwini as type species of Mylodon and the lack of agreement on the assignment of the cranial fragment assigned to Glossotherium by Owen (1839b). Once Kraglievich (1928) had established Mylodon darwini as type species of Mylodon, Grypotherium fell as a synonym of Mylodon. Kraglievich (1928) agreed with Reinhardt (1879) and Smith- Woodward (1900) that the cranial fragment of Glossotherium was congeneric with Mylodon robustus but not conspecific with it. Consequently, and with the general understanding that Mylodon robustus was generically distinct form Mylodon darwini, Kraglievich (1928) revalidated Glossotherium, but with two species, i.e., Glossotherium robustus and Glossotherium uruguayense, the latter including Owen's (1839b) cranial fragment. These nomenclatural conclusions were accepted by Cabrera (1936), although in this author's revision of the species of Glossotherium, he recognized its two valid species as Glossotherium robustum and Glossotherium lettsomi (Owen), with Glossotherium uruguayense a synonym of the latter (Glossotherium lettsomi was originally assigned to Pleurolestodon lettsomi by Gervais and Ameghino (1880), based on observation of a skull exhibited in the Natural History Museum, London, that had been labeled by Owen as Mylodon lettsomi (see Ameghino 1889). Esteban's (1996) review of Mylodontinae considered Glossotherium lettsomi (sensu Cabrera 1936) a synonym of Glossotherium robustum, so that the cranial fragment collected by Darwin in Uruguay is currently assigned to the latter species.
Magnorder Xenarthra Cope, 1889 Order Pilosa Flower, 1883 Suborder Phyllophaga Owen, 1842 Infraorder Megatheria McKenna and Bell, 1987 Superfamily Megatheroidea Gray, 1821 Family Megatheriidae Gray, 1821 Subfamily Megatheriinae Gray, 1821 Genus Megatherium Cuvier, 1796
Megatherium cuvieri Desmarest, 1822 Fig. 6a
 Figure 6: a) Parasagittal section through the maxilla of Megatherium cuvieri (= Megatherium americanum, see text) showing the five molariforms (from Owen 1840). Scale bar = 10 cm; b) Mandible of Megalonyx jeffersoni (= Mylodon darwini, see text) in occlusal view (from Owen 1840). Scale bar = 10 cm
Figure 6: a) Parasagittal section through the maxilla of Megatherium cuvieri (= Megatherium americanum, see text) showing the five molariforms (from Owen 1840). Scale bar = 10 cm; b) Mandible of Megalonyx jeffersoni (= Mylodon darwini, see text) in occlusal view (from Owen 1840). Scale bar = 10 cm
Owen (1840) assigned to this species various cranial fragments collected by Darwin in Punta Alta (Buenos Aires Province). Although Darwin referred most of the large remains he recovered to Megatherium, as shown by his field notes, Beagle Diary, and personal correspondence, Owen (1840) did likewise only for the specimens collected from Punta Alta. In discussing Darwin's (in Keynes 2001, p. 107) "allied to the Rhinoceros" comment, we noted that it did not pertain to the Scelidotherium skeleton but to various skulls assigned by Owen (1840) to Megatherium. Indeed, Darwin's comment, dated September 23, 1832, could only refer to those skulls collected up to this time, effectively excluding Scelidotherium (see above) from consideration, and leaving Megatherium as the only taxon to which he assigned skull material from Punta Alta. Owen's (1840) assignment to the species Megatherium cuvieri was based on acceptance of the specific name change proposed by Desmarest (1822) for Megatherium americanum. However, this proposed nomenclatural alteration is unsupported and the name was considered a nomen illegitimum by Mones (1986). In this context, the specimens assigned to Megatherium cuvieri by Owen (1840) correspond to Megatherium americanum.
Magnorder Xenarthra Cope, 1889 Order Pilosa Flower, 1883 Suborder Phyllophaga Owen, 1842 Infraorder Megatheria McKenna and Bell, 1987 Superfamily Megatheroidea Gray, 1821 Family Megalonychidae Gervais, 1855 Subfamily Megalonychinae Gervais, 1855 Tribe Megalonychini Gervais, 1855 Subtribe Megalonychina Gervais, 1855 Infratribe Megalonychi Gervais, 1855 Genus Megalonyx Harlan, 1825
Megalonyx jeffersonii (Desmarest, 1822) Fig. 6b
Owen (1840) assigned a mandible collected by Darwin at Punta Alta (Buenos Aires Province) to this species. A particular feature of this specimen, figured by Owen (1840, pl. 29), is that it preserves only one of the molariforms of the right dentary. This agrees with the description given by Darwin (in Keynes 2001, p. 109) for a mandible collected October 8, 1832: "After breakfast I walked to Punta Alta, the same place where I have before found fossils… I obtained a jaw bone which contained a tooth: by this I found out that it belongs to the great ante-diluvial animal the Megatherium". Leidy (1852) erected a new taxon, Gnathopsis oweni on this specimen, which Kraglievich (1928) assigned to Mylodon darwini.
Magnorder Xenarthra Cope, 1889 Order Cingulata Illiger, 1811 Superfamily Glyptodontoidea Gray, 1869 Family Glyptodontidae Gray, 1869 Subfamily Hoplophorinae Huxley, 1864 Tribe Hoplophorini Huxley, 1864 Genus Hoplophorus Lund, 1938
Hoplophorus euphractus Lund, 1938 Fig. 7a
 Figure 7: a) Osteoderms assigned by Owen (1840) to Hoplophorus euphractus (= Neosclerocalyptus ornatus, see text) in dorsal and lateral views (from Owen 1840). Scale bar = 1 cm; b) Maxilla and mandible assigned by Owen (1840) to Ctenomys priscus (= Actenomys priscus, see text) (from Owen 1840). Scale bar = 5 mm.
Figure 7: a) Osteoderms assigned by Owen (1840) to Hoplophorus euphractus (= Neosclerocalyptus ornatus, see text) in dorsal and lateral views (from Owen 1840). Scale bar = 1 cm; b) Maxilla and mandible assigned by Owen (1840) to Ctenomys priscus (= Actenomys priscus, see text) (from Owen 1840). Scale bar = 5 mm.
In general, the carapace fragments and osteoderms collected by Darwin in South America were assigned preliminarily to Megatherium. Certainly such assignments were due to Cuvier's (1823) tacit approval of Larrañaga's (in Cuvier 1823) suggestion that megatheres were armored. Owen (1840, p. 107) recognized the close relationship, still considered correct, between glyptodonts and living armadillos and remarked the following about Hoplophorus euphractus: "The portions of the tesselated bony dermal covering of a Dasypodoid quadruped, figured in Pl. XXXII. figs. 5 and 4, of the natural size, were discovered folded round the middle and ungueal phalanges, figs. 2 and 3, at Punta Alta, in Bahia Blanca, in an earthy bed interstratified with the conglomerate containing the remains of the fossil Edentals. In one of these fragments, measuring six inches long by five broad, the tesserae are arranged in rosettes, and so closely correspond in size and pattern with the bony armour described by M. Lund [1839], as characterizing his species, Hoplophorus euphractus, that I feel no hesitation in referring them to that animal". Burmeister (1870-1874) assigned the osteoderms figured by Owen (1840, pl. 32, figs 4 and 5) to Hoplophorus ornatus (= Neosclerocalyptus ornatus) and Hoplophorus elegans, respectively. Ameghino (1889) synonymized the latter with Lomaphorus elegans.
Magnorder Xenarthra Cope, 1889 Order Cingulata Illiger, 1811 Superfamily Glyptodontoidea Gray, 1869 Family Glyptodontidae Gray, 1869 Subfamily Glyptodontinae Gray, 1869 Tribe Glyptodontini Gray, 1869 Genus Glyptodon Owen, 1839c
Glyptodon clavipes Owen, 1839c
Owen (1840) tentatively assigned to this species distinct osteoderms collected by Darwin from the Uruguayan locality of Arroyo Sarandí (Mercedes, of Soriano Departament, Uruguay); and the Argentinean localities of Bajada Santa Fe (Paraná, Entre Ríos Province) and Guardia del Monte (Buenos Aires Province).
Magnorder Epitheria McKenna, 1975 Superorder Preptotheria McKenna, 1975 Grandorder Anagalida Slazay and Mc Kenna, 1971 Mirorder Simplicidentata Weber, 1904 Order Rodentia Bowdish, 1821 Suborder Hystricognatha Woods, 1976 Infraorder Hystricognathi Tullberg, 1899 Parvorder Caviida Bryant and McKenna, 1995 Superfamily Cavioidea Fisher de Waldheim, 1817 Family Hydrochoeridae Gray, 1825 Subfamily Hydrochoerinae Gray, 1825 Genus Hydrochoerus Brisson, 1762
Hydrochoerus sp.
The fossils that Owen (1940) assigned to this genus were collected by Darwin (in Keynes 2001) in Monte Hermoso on October 19, 1832. Owen (1840, p. 10) commented that "Mr. Darwin discovered the decomposed molar of a Rodent, equalling in size, and closely resembling in the disposition of its oblique component laminae, the hinder molar of the Capybara (Hydrochoerus.) The fossil differs, however, in the greater relative breadth of the component laminae. I have, lastly, to notice the head of a femur, and some fragments of pelvic bones from the same formation which bear the same proportion to the tooth above alluded to, as subsists between the teeth and bones of the Capybara, and which are sufficient to prove that there once has existed in South America a species of the family Caviidæ, as large as the present Capybara, but now apparently extinct". These elements were among those that Darwin (in Keynes 2001, p. 110-111) encountered as follows: "The Captain landed for half an hour at Monte Hermoso, (or Starvation point as we call it) to take observations. - I went with him & had the good luck to obtain some well preserved fossil bones of two or three sorts of Gnawing animals. - One of them must have much resembled the Agouti but it is smaller". Even so, it is not possible to determine which of the remains Darwin thought similar to an agouti, though it is worth noting that what Darwin considered an agouti was a Patagonian hare (Darwin, in Keynes 2001, p. 103). The following Hydrochoeridae are currently recognized from the fossiliferous horizons of Monte Hermoso: Phugatherium cataclisticum, Anchimysops villalobosi and Chapalmatherium perturbidum. Vucetich et al. (2005) considered the type specimens of the first two species to represent juvenile individuals and that they might be conspecific with Chapalmatherium perturbidum. If this is correct, then the valid name, as determined by Vucetich et al. (2005), for the species would be Phugatherium cataclisticum. In this respect, the specimen collected by Darwin might represent this latter species.
Magnorder Epitheria McKenna, 1975 Superorder Preptotheria McKenna, 1975 Grandorder Anagalida Slazay and Mc Kenna, 1971 Mirorder Simplicidentata Weber, 1904 Order Rodentia Bowdish, 1821 Suborder Hystricognatha Woods, 1976 Infraorder Hystricognathi Tullberg, 1899 Parvorder Caviida Bryant and McKenna, 1995 Superfamily Octodontoidea Waterhouse, 1839 Family Octodontidae Waterhouse, 1839 Subfamily Octodontinae Waterhouse, 1839 Genus Ctenomys Blainville, 1826
Ctenomys priscus Owen, 1840 Fig. 7b
This species was erected by Owen (1840) based on remains collected by Darwin (in Keynes 2001) on October 11, 1832. Owen (1840, p. 109) allocated to this species a "…fragment of the upper jaw, figured in Pl. XXXII. fig. 6" and a "...fragment of the lower jaw of the same fossil Rodent . figured at fig. 10 and 11". It should be noted that Owen's (1840) figure 6 possibly does not represent an upper jaw, but a posterior fragment of a mandible that may correspond to the posterior part of the mandible figured by Owen (1840, figs. 10, 11) (D. Verzi, pers. comm.). Mones (1994) provided a detailed discussion of the taxonomy of Ctenomys priscus, concluding that it might belong to the genus Dicoelophorus, erected by Ameghino (1888). However, the generic name currently in use for this species is Actenomys, erected by Burmeister (1888). The reasons for maintaining Actenomys were clearly set forth by Verzi (1994, p. 183), as follows "Dicoelophorus Ameghino, 1888 es, por prioridad, un sinónimo "senior" de Actenomys Burmeister… . Sin embargo, luego de que Reig (1958) adoptara el nombre genérico Actenomys para todos los materiales referibles a las formaciones Monte Hermoso y Chapadmalal, este nombre ha sido ampliamente usado hasta el presente por lo cual nosotros nos inclinamos a mantenerlo".
Rodentia
A third rodent was briefly described by Owen (1840) but not assigned to a genus. Owen (1840, p. 110) described "The portion of the right hind-foot of the Rodent figured at fig. 12, includes the calcaneum, astragalus, cuboides, external and middle cuneiform bones, and the metatarsals and proximal phalanges of the toes corresponding with the three middle toes of five-toed quadrupeds. The metatarsals are chiefly remarkable for the well-developed doubletrochlear articular surface, and intermediate ridge. These remains, as well as the jaws and teeth of the Ctenomys, were discovered at Monte Hermoso in Bahia Blanca". These pedal remains are possibly those that Darwin (1985, Letter 192) described in a letter, dated November 12, 1832, to John Stevens Henslow as "the Tarsi & Metatarsi very perfect of a Cavia" although it is not possible to determine this with certainty.
Mastodonts and armored megatheres
As noted in the preceding historical summaries, the large South American fossil mammals recognized by Cuvier (1823) amounted to an armored megathere and three mastodonts. A detailed consideration of Darwin's preliminary assignments demonstrates that he was strongly influenced by Cuvier's taxonomic scheme. In essence, the fossil specimens recognized by Darwin may be grouped as large vs. medium and small mammals. Among the latter is the first fossil of a South American horse, obviously identified as such by Darwin, and three rodents, two of which were assigned by Darwin to Cavia and the Patagonian hare (Dolichotis patachonica), whereas the third was later considered by Owen (1840) as representing the genus Hydrochoerus. Among the large mammals, Darwin recognized an enormous rodent and four genera: Rhinoceros, Megalonyx, Megatherium and Mastodon. The first two listed do not appear in Cuvier's (1823) taxonomic list of fossil South American mammals. The assignment to Rhinoceros was the first such attempt by Darwin (in Keynes 2001, p. 107) for a South American fossil skull, collected during his first visit to Punta Alta (September, 1832). However, this assignment does not appear in the taxonomic list of specimens collected from Punta Alta and Monte Hermoso sent to John Stevens Henslow (Darwin 1985, Letter 192). Darwin may possibly have changed his first impression, but it is not easily ascertainable with the available evidence. The only mention of Megalonyx appears in the previously noted list as "2nd the upper jaw & head of some very large animal, with 4 square hollow molars.-& the head greatly produced in front.- I at first thought it belonged either to the Megalonyx or Megatherium.- In confirmation, of this, in the same formation I found a large surface of the osseous polygonal plates, which "ate observations" (what are they?) show belong to the Megatherium.- Immediately I saw them I thought they must belong to an enormous Armadillo, living species of which genus are so abundant here…". In this as in many other cases (see below) Darwin decided in favor of Megatherium based on the presence of osteoderms collected in the same formation. Owen (1838-1840) recognized the specimens assigned by Darwin to Megatherium as glyptodonts, toxodonts, and large ground sloths. In our estimation, the only factor that led Darwin to assign remains of such a heterogeneous group of forms to Megatherium was the presence of osteoderms or carapace fragments in the same horizon. In effect, all the specimens that Darwin assigned to Megatherium were recovered from the Arroyo Sarandí (Uruguay) and Punta Alta (Argentina), and Darwin also reported carapace elements. Further evidence supporting our contention appears in Darwin's (1985, Letter 215) letter of September 20, 1833 sent to Caroline S. Darwin, in which he wrote "At the Guardia del Monte, I found some more of the armour of the giant Megatherium, which was to me very interesting, as connecting the Geology of the different parts of the Pampas". Indeed, Darwin used the presence of osteoderms as a diagnostic feature of Megatherium. In this context, one might hypothesize that it was the absence of osteoderms in the layers yielding the only Santa Cruz specimen, which led Darwin to regard this specimen as follows: "At Port St Julian I found some very perfect bones of some large animal, I fancy a Mastodon" (Darwin 1985, Letter 238) and Darwin (1839) later admitted that "I had no idea at the time, to what kind of animal these remains belonged". Certainly, this is the only mention of Mastodon that was not later corroborated by Owen (1838-1840). Lastly, Toxodon was the only species the remains of which Darwin (1985, Letter 192, 2002-2008) assigned to different species. Darwin assigned a molar collected in Punta Alta to an enormous rodent and a skull from Arroyo Sarandí to Megatherium. The latter assignment makes sense in that the horizon yielding the skull also preserved osteoderms, whereas Darwin's inference on the molar is understandable given that the teeth of Toxodon, as noted by Owen (1838), bear a certain resemblance to those of rodents.
Implications of the "erroneous" assignments of the youthful Darwin
Twenty years before the publication of the Origin of Species and only two years after he had begun recording his thoughts on transmutation, Darwin (1839, p. 209-210) had this to say about the South American paleofauna: "The most important result of this discovery, is the confirmation of the law that existing animals have a close relation in form with extinct species. As the guanaco is the characteristic quadruped of Patagonia, and the vicuna of the snow-clad summits of the Cordillera, so in bygone days, this gigantic species of the same family must have been conspicuous on the southern plains. We see this same relation of type between the existing and fossil Ctenomys, between the Capybara (but less plainly, as shown by Mr. Owen) and the gigantic Toxodon; and lastly, between the living and extinct Edentata. At the present day, in South America, there exist probably nineteen species of this order, distributed into several genera; while throughout the rest of the world there are but five. If, then, there is a relation between the living and the dead, we should expect that the Edentata would be numerous in the fossil state. I need only reply by enumerating the Megatherium, and the three or four other great species, discovered at Bahia Blanca; the remains of some of which are also abundant over the whole immense territory of La Plata. I have already pointed out the singular relation between the armadilloes and their great prototypes, even in a point apparently of so little importance as their external covering". Similarly, Darwin (1845, 1859, p. 339) noted that "In South America, a similar relationship is manifest, even to an uneducated eye, in the gigantic pieces of armour like those of the armadillo, found in several parts of La Plata; and Professor Owen has shown in the most striking manner that most of the fossil mammals, buried there in such numbers, are related to South American types…… I was so much impressed with these facts that I strongly insisted, in 1839 and 1845, on this 'law of the succession of types,'-on 'this wonderful relationship in the same continent between the dead and the living". As noted by Sulloway (1982), the role of the naturalists who studied the South American fauna was critical to the development of Darwin's transformationist ideas in that they provided a systematic framework. In reference to Darwin's erroneous taxonomic assignments, Sulloway (1982, p. 353) recognized that "The general effect of these confusions during the Beagle voyage was to minimize the evolutionary implications of the diverse fossil forms". Paradoxically, many of Owen's (1838-1840) proposed relationships that so influenced Darwin (1839, 1845, 1859) in the development of his transformationist theory were later rejected. Indeed, toxodonts and macrauchenids are highly derived native South American ungulates distantly related phylogenetically to rodents and guanacos, whereas the large glyptodonts are not the ancestors of armadillos, but to the contrary, the latter are antecedent to the former. In this respect, we may suppose that the affinities proposed by Owen (1838-1840) maximized the evolutionary implications of the South American paleofauna.
List of fossil mammal specimens collected by Charles Darwin and housed in the Natural History Museum, London, England (provided by Andrew Currant, Curator of Fossil Mammals), and in the Royal College of Surgeons of England (provided by Simon Chaplin, Director of Museums and Special Collections). Abbeviations. RCSHM/CO: Royal College of Surgeons of England Hunterian / College Museum. RCS: Royal College of Surgeons of London. NHM: Natural History Museum, London.
Equus sp. NMH M16557: left upper molar (ex RCS2012). NMH M16558: left upper molar (ex RCS2013). Glossotherium sp. NMH M16586 part of cranium (ex RCS3491). Machrauchenia patachonica. Holotype NMH M43402: vertebral fragments (ex NMH M16556 and ex RCS2209); detached proximal and distal ends of right tibia and fibula (ex NMH M16559 and ex RCS 2215); right femur (ex NMH M16561 and ex RCS2214); right astragalus (ex M16569 and ex RCS2216); middle cervical vertebrae (ex NMH M16570 and ex RCS2208); fragments of pelvis (ex NMH M16571 and ex RCS2210); proximal end of fused radius and ulna (ex NMH M16572 and ex RCS2212); metacarpals and phalanges of the right manus (ex NMH M16573 and ex RCS2213); incomplete left scapula (ex NMH M16574 and ex RCS2211); metatarsal (ex M16575 and ex RCS2216). Megatherium americanum. NMH M16585, posterior part of cranium (ex RCS3445); NMH M16588, fragment of left maxilla with M2 and M3 in horizontal section (ex RCS3446); NMH M16589, part of left temporal (ex RCS3457). RCSHM/CO 3443 fragment of cranium with teeth. Mylodon darwini. Holotype NMH M16563: mandible (ex RCS3490). Mylodon darwini. NMH M16587: fragment of left dentary with transverse section of teeth (ex RCS3491). Scelidotherium leptocephalum. Holotype: NMH M16579, deformed skull with horizontal section taken from right dentary (ex RCS3506); Holotype: NMH M16580, upper molariform (ex RCS 3507); Holotype: NMH M16581, cervical vertebrae (ex RCS3510); Holotype: NMH M16582, parts of ribs (ex RCS 3511); Holotype: NMH M16583, left astragalus (ex RCS3520); Holotype: M16584, right astragalus with distal end of right tibia (ex RCS3519). Toxodon platensis. Holotype NMH M16560: incomplete cranium lacking teeth (ex RCS2223). Toxodon platensis. NMH M16562 right M2 (ex RCS2224); NMH M16564 part of left dentary with fragments of four molars (ex RCS2226); NMH M16565 right lower molar (ex RCS2227); NMH M16566 fragment of right dentary with incisor roots and most of the molars (ex RCS2225); NMH M16567 left lower incisor (ex RCS2228); M16568 fragmentary teeth (ex RCS 2229).
ACKNOWLEDGMENTS
We are grateful to the editors for the invitation to participate in this special issue of Revista de la Asociación Geológica Argentina, and to Greg McDonald and Mariano Bond for their thoughtful comments and suggestions that improved the manuscript. Andrew Currant, Curator of Fossil Mammals Palaeontology, Natural History Museum London, Simon Chaplin, Director of Museums and Special Collections and Sarah Pearson, Assistant Curator, The Royal College of Surgeons of England, for information on Darwin's fossil mammal collections, and Louise King, Assistant Archivist of The Royal College of Surgeons of England, for invaluable help on the bibliography.
WORKS CITED IN THE TEXT
1. Alberdi, M.T. and Prado, J.L. 1995. Los Mastodontes de América del Sur. In Alberdi, M. T., Leone, G. and Tonni, E.P. (eds.) Evolución climática y biológica de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental, Consejo Superior de Investigaciones Científicas, Museo Nacional de Ciencias Naturales, Monografías: 279-292, Madrid.
2. Ameghino, F. 1888. Lista de las especies de mamíferos fósiles de Mioceno superior de Monte Hermoso, hasta ahora conocidas. P.E. Coni, 21 p., Buenos Aires.
3. Ameghino, F. 1889. Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias 6: 1-1027, Córdoba.
4. Bond, M. 1999. Quaternary native ungulates of Southern South America. A synthesis. In Rabassa, J. and Salemme, M. (eds.) Quaternary of South America and Antarctic Peninsula 12: 177-206.
5. Brown, B. 1903. A new genus of ground sloth from the Pleistocene of Nebraska Paramylodon nov. gen. Bulletin of the American Museum of Natural History 19: 569-583.
6. Burmeister, G. 1866. Sobre el género Toxodon. Actas de la Sociedad Paleontológica de Buenos Aires 1: 16-17.
7. Burmeister, G. 1870-1874. Monografía de los glyptodontes en el Museo Público de Buenos Aires. Anales del Museo Público de Buenos Aires 2: 1-412.
8. Burmeister, G. 1888. Restos de diez diferentes cuadrúpedos fósiles que ha mandado mi hijo Carlos al Museo Nacional. Anales del Museo Nacional de Buenos Aires 3(15): 178-180.
9. Cabrera, A. 1929. Una revisión de los mastodontes argentinos. Revista del Museo de La Plata 32: 61-144.
10. Cabrera, A. 1936. Las especies del género Glossotherium. Notas del Museo de La Plata Paleontología 1(5): 193-206.
11. Cave, A.J.E. 1942. Museum Royal College of Surgeons of England. Scientific Report for the year 1940-1941: 3-16.
12. Cifelli, R.L. 1993. The phylogeny of the Native South American Ungulates. In Szalay, F. S., Novacek, M.J. and McKenna, M.C. (eds.) Mammal Phylogeny: Placentals, Springer- Verlag, p. 195-266, New York.
13. Cieza de León, P. 1553. La Crónica del Perú. Edición de Calpe 1922, 367 p., Madrid.
14. Clift, W. 1835. Some account of remains of the Megatherium sent to England from Buenos Aires by Woodbine Parish. Transactions of the Geological Society 2: 437-50.
15. Cuvier, G. 1796. Notice sur le squelette d'une très-grande espèce de quadrupède inconnue jusqu'à présent, trouvé au Paraquay, et déposé au cabinet d'histoire naturelle de Madrid. Magasin Encyclopèdique: ou Journal des Sciences, des Lettres et des Arts 1: 303-310.
16. Cuvier, G. 1806. Sur differéntes dents du genre des mastodontes, mais d'espèces moindres que celle de olió, trouvées en plusieurs Linux des deux continens. Annales du Muséum d' Histoire Naturelle 8: 401-420.
17. Cuvier, G. 1823. Réchèrches sur les ossemens fossiles. 2nd ed. Dufour & D'Ocagne, Paris.
18. Darwin, C.R. 1839. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Journal and remarks. 1832-1836. Henry Colburn, 620 p., London.
19. Darwin, C.R. 1845. Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. Beagle round the world, under the Command of Capt. Fitz Roy, R.N. John Murray (2d edition), 520 p., London.
20. Darwin, C.R. 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. John Murray (1st edition),502 p., London.
21. Darwin, C.R. 1985. The Correspondence of Charles Darwin. Burkhardt, F. and Smith, S. (eds.) Cambridge University Press. Volume 1, 1821-1836. Cambridge.
22. Darwin C.R. 2002-2008. The Complete Work of Charles Darwin Online. University of Cambridge. [http://darwin-online.org.uk/]
23.Desmarest, A.G. 1822. Mammalogie ou déscription des espèces de mammifères 2: 27-555.
24. Esteban, G.I. 1996. Revisión de los Mylodontinae cuaternarios (Edentata-Tardigrada) de Argentina, Bolivia y Uruguay. Sistemática, Filogenia, Paleobiología, Paleozoogeografía y paleoecología. Tesis Doctoral Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucumán, 235 p. (unpublished).
25. Falkner, T. 1774. A description of Patagonia and the adjoining parts of South America: containing an account of the soil, produce, animals, vales, mountains, Rivers, lakes, etc. of those countries; the religion, government, policy, customs, dress, arms, and language of the Indian inhabitants; and some particulars relating to Falkland's Islands. C. Pugh, 144 p., London.
26. Gervais, H. and Ameghino, F. 1880. Los mamíferos fósiles de la América del Sur . F. Savy-Igon Hnos, 225 p., París-Buenos Aires.
27. Harlan, R. 1835. Medical and physical researches or original memoirs in medicine, surgery, physiology, geology, zoology, and comparative anatomy. Lydia R. Bailey, 653 p., Philadelphia.
28. Keynes, R.D. 2001. Charles Darwin's Beagle Diary. Cambridge University Press. 466 p., Cambridge.
29. Kraglievich, L. 1928. Mylodon darwini Owen es la especie genotipo de Mylodon Owen. Physis 9: 169-185.
30. Leidy, J. 1852. On extinct Edentata of North America. Proceedings of the Academy of Natural Science 6(4): 117.
31. Leidy, J. 1855. A memoir on the extinct Sloth Tribe of North America. Smithsonian Contribution to Knowledge 7: 1-68.
32. López Piñeiro, J.M. and Glick, T.F. 1993. El megaterio de Bru y el presidente Jefferson. Una relación insopechada en los albores de la paleontología. Cuadernos Valencianos de Historia de la Medicina y de la Ciencia Serie A (Monografías) 42, 157 p., Valencia.
33. Lydekker, R. 1887. Catalogue of the fossil Mammalia in the British Museum of Natural History. Part V. 346 p., London.
34. McKenna, M.C. and Bell, S.K. 1997. Classification of mammals above the species level. Columbia University Press, 631 p., New York.
35. Mones, A. 1986. Palaeovertebrata Sudamericana. Catálogo sistemático de los vertebrados fósiles de America del Sur, Parte1. Lista preliminar y bibliografía. E. Schweizerbart, Courier Forschungsinstitut Seneckenberg 82: 1-625, Stuttgart.
36. Mones, A. 1994. Los silencios de Florentino Ameghino. Problemas de bibliografía, sistemática y nomenclatura. Senckenbergiana biologica 74(1-2): 9-20.
37. Owen, R. 1837. A description of the cranium of the Toxodon platensis, a gigantic extinct mammiferous species, referrible by its dentition to the Rodentia, but with affinities to the Pachydermata and the herbivorous Cetacea. Proceeding of the Geological Society 2: 541-542, London.
38. Owen, R. 1838. Fossil Mammalia. In Darwin, C.R. (ed.) Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitz-roy, during the years 1832 to 1836. 1(1): 13-40.
39. Owen, R. 1839a. Description of a tooth and part of the skeleton of the Glyptodon clavipes, a large quadruped of the Edentate order, to which belongs the tesselated bony armour described and figures by Mr. Clift in the former Volume of the Transactions of the Geological Society; with a consideration of the question whether the Megatherium possessed an analogous dermal armour. Transactions of the Geological Society 2: 81-106, London.
40. Owen, R. 1839b. Fossil Mammalia. In Darwin, C.R. (ed.) Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitz-Roy, during the years 1832 to 1836. 1(2-3): 41-80.
41. Owen, R. 1839c. Description of a tooth and part of the skeleton of the Glyptodon, a large quadruped of the edentate order, to which belong the tesselated bony armour figured by Mr. Clift in his memoir on the remains of the Megatherium, brought to England by Sir Woodbine Parish, F. G. S. Proceedings of the Geological Society 3: 108-113, London.
42. Owen, R. 1840. Fossil Mammalia. In Darwin, C.R. (ed.) Zoology of the Voyage of H.M.S Beagle, under the command of Captain Fitzroy, during the years 1832 to 1836. 1(4): 81-111.
43. Owen, R. 1842. Description of the skeleton of an extinct gigantic sloth, Mylodon robustus, Owen, with observations on the osteology, natural affinities, and probable habits of the megatherioid quadruped in general. R. and J.E. Taylor, 176 p., London.
44. Owen, R. 1845. Descriptive and illustrated catalogue of the fossil organic remains of Mammalia and Aves contained in the Museum of the Royal College of Surgeons of England. R. and J.E. Taylor, 391 p., London.
45. Prado, J.L. and Alberdi, M.T. 1995. A quantitative review of the horses Equus from South America. Palaeontology 37: 459-481.
46. Reig, O.A. 1958. Notas para una actualización del conocimiento de la fauna de la Formación Chapadmalal. 1. Lista faunística preliminar. Acta Geológica Lilloana 2: 241-253.
47. Reinhardt, J.T. 1879. Beskrivelse af Hovedskallen af et Kaempedovendyr, Grypotherium darwinii. Videnskavernes Selskabs Skrifter. 5 Raekke. Naturvidensakbelig og Mathematisk Afdeling 12(4): 351-381.
48. Smith-Woodward, A. 1900. On some remains of Grypotherium (Neomylodon) listai and associated mammals from a cavern near Consuelo Cave, Last Hope Inlet, Patagonia. Proceeding of the Zoological Society 5: 64-79, London.
49. Sulloway, F.J. 1982. Darwin's Conversion: The Beagle Voyage and Its Aftermath. Journal of the History of Biology 15(3): 325-396.
50. Verzi, D.H. 1994. Origen y Evolución de los Ctenomyinae (Rodentia, Octodontidae): un análisis de anatomía cráneo-dentaria. Tesis Doctoral, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, (unpublished), 227 p., La Plata.
51. Vucetich, M.G., Deschamps, C.M., Itatí Olivares, A. and Dozo, M.T. 2005. Capibaras, size, shape, and time: A model kit. Acta Paleontologica Polonica 50(2): 259-272.
52. Weiss, C.S. 1830. Ueber das südliche Ende des Gebigszuges von Brasilien in der Provinz S. Pedro do Sul und der Banda Oriental oder Staate von Monte Video: nach den Samlungen des Herrn Fr. Sellow. Abhandlungen der physikalischen Klasse del Königlichen Akademie der Wissenschaften zu Berlin, 217-293.
Young Darwin and the ecology and extinction of pleistocene south american fossil mammals
Sergio F. Vizcaíno1, Richard A. Fariña2 and Juan Carlos Fernicola3
1 División Paleontología Vertebrados, Museo de La Plata, La Plata, Argentina. CONICET. Email: vizcaino@fcnym.unlp.edu.ar 2 Departamento de Paleontología, Facultad de Ciencias, Universidad de la República, Montevideo. Uruguay. Email: fari~a@fcien.edu.uy 3 Sección Paleontología de Vertebrados, Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", Buenos Aires. Argentina. Email: jctano@macn.gov.ar
ABSTRACT
During his two years in South America Charles Darwin became fascinated not only with the lush vegetation of Brazil, but also with the gigantic Pleistocene mammals that he found in the drier areas of Uruguay, and in the pampas and Patagonian coast of Argentina. These findings included various ground sloths and glyptodonts among xenarthrans, and hoofed herbivores like Toxodon and Macrauchenia, in addition to horses and small rodents. He concluded that the general assumption that large animals require luxuriant vegetation was false and that vitiated the reasoning of geologists on some aspects of Earth's history. He also reflected on the evident changes that occurred in the continent, the extinct fauna of which suggested to him an analogy to southern parts of Africa. He wondered about our ignorance of biological traits in extinct creatures and the reasons for their extinction. Thus, not only did Darwin inspire phylogenetic studies on fossil mammal lineages, he also opened a gate to the research on their behaviour, physiology and extinction; i.e., their palaeobiology. Whereas the first approach was largely developed in South America beginning about the second half of the 19th century due to the intellectual influence of Florentino Ameghino, palaeobiology became a much more recent line of work, in apparent relation to innovations in methodology and technology. This contribution provides an overview of recent contributions on the palaeobiology of Pleistocene fossil mammals of South America as attempts to provide answers for Darwin's questions.
Keywords: Darwin; Ecology; Extinction; South America; Mammals.
RESUMEN: El joven Darwin y la ecología y extinción de los mamíferos fósiles sudamericanos. Durante los dos años que Charles Darwin estuvo en América del Sur no sólo se deslumbró con la profusa vegetación de Brasil, si no también con los gigantescos mamíferos pleistocenos que colectó en áreas más secas de Uruguay y en la pampa y la costa patagónica de Argentina. Sus hallazgos incluyeron distintos perezosos y gliptodontes, ungulados herbívoros como Toxodon y Macrauchenia, además de caballos y pequeños roedores. Darwin, desechó la presunción general de que los grandes animales requieren una exuberante vegetación, reconociendo que la misma condicionó las interpretaciones que hicieran los geólogos sobre algunos aspectos de la historia de la Tierra. También reflexionó sobre los cambios acaecidos en el continente, cuya fauna le sugirió una clara analogía con lo observado en el sur de África. Darwin se preguntó acerca de nuestro desconocimiento de las características biológicas y de las causas que llevaron a su extinción. Así, no sólo inspiró el estudio filogenético de distintos linajes de mamíferos fósiles, si no que abrió la puerta a las investigaciones sobre su comportamiento, fisiología y extinción; i.e. su paleobiología. En América del Sur, la influencia intelectual de Florentino Ameghino permitió apuntalar el estudio filogenético de los mamíferos durante la segunda mitad del Siglo XIX, mientras que el fortalecimiento de la paleobiología se dio en tiempos recientes en relación con innovaciones metodológicas y tecnológicas. Esta contribución provee una visión global de las contribuciones realizadas en paleobiología de mamíferos pleistocenos de América del Sur como un intento de responder los cuestionamientos que se hiciera Darwin.
Palabras clave: Darwin; Ecología; Extinción; América del Sur; Mamíferos.
INTRODUCTION
When HMS Beagle reached the coast of Brazil in February, 1832, Charles Darwin was a 22-year old theology student at Cambridge University and his ambition was to become a rural pastor. He had started to develop an interest in natural history some time before, while studying medicine at Edinburgh University. The overwhelming power of South American biodiversity that greeted the young Charles Darwin led him to declare that "The elegance of the grasses, the novelty of the parasitical plants, the beauty of the flowers, the glossy green of the foliage, but above all the general luxuriance of the vegetation, filled me with admiration." (Voyage of the Beagle, Chapter I, Feb. 1832) However, it was not only the luxuriance that influenced his mind but also the lesser plains of the south and their fossils. Actually, he became fascinated by the discovery of gigantic Pleistocene mammals during his journeys in Uruguay, and the Pampean region and Patagonian coast of Argentina, before leaving the continent in May 1834 (see letters sent by Charles Darwin to Caroline Darwin and John Stevens Henslow in Burkhardt and Smith 1985, p. 276 and 280 respectively). As developed more fully in another contribution (see Fernicola et al., 2009) the extraordinary zoological collection of fossil and extant specimens made by Charles Darwin during his voyage to South America was studied by an important group of naturalists who published their conclusions between February 1838 and October 1843 in the "The Zoology of the Voyage of H.M.S. Beagle". All fossils mammals included in this work were studied by Richard Owen, who recognized the South American ungulates Toxodon platensis and Macrauchenia patachonica, a fossil horse identified as Equus sp., the mastodont Mastodon angustidens, and the ground sloths Glossotherium sp., Mylodon darwinii, Scelidotherium leptocephalum, Mega-lonyx jeffersonii, Megatherium cuvierii, and the glyptodonts Glyptodon clavipes, Hoplophorus euphractus among xenarthrans, as well as some smaller forms like rodents. The taxonomic history of several of these taxa is complex, including issues as varied as nomenclatural problems (e.g. Megatherium cuvierii nomen illegit.) to the mixing of specimens of different species or genera assigned to the same species (e.g. Megatherium americanum), an issue that is treated by Fernicola et al. (2009). It is widely accepted that by the time Darwin boarded the Beagle he had been influenced by Lamarck's ideas on evolution (see Woodward 1987). Also, that the giant fossil quadrupeds he found were significant toward the development of his evolutionary theory, as very early they suggested to him that the similarities between extinct and living forms should be explained by the existence of common ancestors, and that the transformation of species to a large degree was not a vertical sequence, as Lamarck had proposed, but a tree with asymmetric branches (see Huxley and Kettlewell 1965). In his own words: "to my view, in S. America parent of all armadilloes might be brother to Megatherium - uncle now dead" (Darwin, 1837-1838 in Barret 1960). Woodward (1987) stated that by that time Darwin was also familiar with adaptation (The red notebook of Charles Darwin in Herbert 1980, p. 67), another important biological issue related to evolution, but an idea then associated with Natural Theology (Paley 1802), which argued that every organism was intentionally perfectly designed to its particular life conditions by God. But it was not only the idea, means and processes of evolution that impressed Darwin. During his journey between Buenos Aires and Santa Fe he wrote "We may therefore conclude that the whole area of the Pampas is one wide sepulchre for these extinct quadrupeds" (Voyage of the Beagle, Chapter VII, Oct. 1833). By that time his sharp mind had already noted "That large animals require luxuriant vegetation has been a general assumption, which has passed from one work to another. I do not hesitate, however, to say that is completely false; and that it has vitiated the reasoning of geologists, on some points of great interest in the ancient history of the world" (Chapter V, of the Beagle, Aug. 1833, The red notebook of Charles Darwin in Herbert 1980, p. 54). In January 1834, Darwin collected remains of Macrauchenia in Patagonia near San Julián, in what today is Santa Cruz province. He made inferences about the environment in which this beast lived and reflected on its extinction: "Mr. Owen... considers that they form part of an animal allied to the guanaco or llama, but fully as large as the true camel. As all the existing members of the family of Camelidae are inhabitants of the most sterile countries, so we may suppose was this extinct kind… It is impossible to reflect without the deepest astonishment, on the changed state of this continent. Formerly it must have swarmed with great monsters, like the southern parts of Africa, but now we find only the tapir, guanaco, armadillo, capybara; mere pigmies compared to antecedents races... Since their loss, no very great physical changes can have taken place in the nature of the Country. What then has exterminated so many living creatures?…We are so profoundly ignorant concerning the physiological relations, on which the life, and even health (as shown by epidemics) of any existing species depends, that we argue with still less safety about either the life or death of any extinct kind" (Voyage of the Beagle, Chapter IX, Jan. 1834). In this way, Darwin not only triggered the studies on the genealogical interpretation of the fossil mammal lineages, but also opened a gate to the research on their behaviour, physiology and extinction; in others words, on their palaeobiology. Phylogenetic studies, combined with morphological and taxonomic analyses, flourished in South America, particularly beginning during the latter half of the 19th century due to the intellectual influence of Florentino Ameghino, but palaeobiology became a much more recent line of work, in apparent relation to innovations in methodology and technology. Indeed, Pleistocene South American fossil mammals show a greater morphological diversity than their living counterparts, as they include representatives of great body size and very peculiar features. Their peculiarity and general lack of modern analogues have encouraged creative palaeobiological approaches that will be outlined below. The aim of this contribution is to overview the recent contributions on the palaeobiology and palaeoecology of the Pleistocene fossil mammals of South America, in order to investigate if we can provide answers for the questions that the young Darwin made himself when he first collected them.
RECONSTRUCTING PALAEOBIOLOGY OF SOUTH AMERICAN PLEISTOCENE MAMMALS
Palaeobiologists are interested in reconstructing the form of the fossils as living animals, their habitat, ecological role, behaviour, and basic biology. Vizcaíno et al. (2004) and Vizcaíno et al. (2008) describe a basic protocol for palaeobiological studies that identifies three biological attributes that are essential for each taxon: size, diet and usage of substratum or type of locomotion. Such principles have been used for the last three decades (e.g., Andrews et al. 1979, Van Couvering 1980, Reed 1998, etc.) though not fully applied to South Aerican mammals. Morphological study of the masticatory and locomotor apparatuses allows predictions on the movements for which the apparatuses are optimized. In addition, analyses of mastication are useful for formulating hypotheses about the diet of the organism, while analyses of the locomotor apparatus allow inferences about the type of locomotion or preferences in the usage of substratum: runner, hopper, digger, burrower, etc. Obviously, these two aspects, added to body size, yield relevant data for the interpretation of an organism in a palaeobiological context. Palaeomammalogists have largely applied actualism, according to which past events are surmised by analogy with currently observable processes assuming that fossil species had similar habits to their current relatives. However, when phylogenetic affinity is not very close or fossil lineages possess morphologies not represented in extant species (Vizcaíno et al. 2004), this methodology does not provide reliable results, a circumstance that is particularly applicable to the mammalian faunas that evolved in relative isolation in South America during a good part of the Tertiary. Vizcaíno et al. (2004) and Vizcaíno et al. (2008) provide accounts on the reconstruction of palaeobiology through the application of the "form-function correlation approach" (Radinsky 1987), according to which function can be inferred from form, to make good use of the main sources of information like fossilized bones and teeth (though indirect evidence can also be used). Form-function relationships can be studied through different approaches, like functional morphology, biomechanics and ecomorphology (see definitions in Vizcaíno et al. 2008, and references therein) It was not until the second part of the 1990s that authors began to apply biomechanic, morpho-geometrical, and ecomorphological methods to the study of morphology as part of a major project aimed at understanding the great palaeobiological diversity of the South American extinct forms. These results allowed the development of novel interpretations of their modes of life that, coupled with palaeoenvironmental data (geology, palaeoclimatology and reconstruction of palaeovegetation), provide insightful information on the paleoecological context in which these animals existed.
Body size
Body size has a remarkable influence on an animal's life because it can be correlated, among other features, with metabolism, limb bone dimensions and biomechanics of locomotion, or particular solutions for food intake. Body mass in Pleistocene xenarthrans was estimated using scale and computer generated (geometric) models, and allometric equations (see Vizcaíno et al. 2008 and references therein). Using these approaches, Fariña et al. (1998), Bargo et al. (2000) and Christiansen and Fariña (2003) estimated the masses of most Lujanian megamammals. Among the late Quaternary mammals of South America, three species rival for the title of the largest of them: the giant ground sloths Megatherium americanum and Eremotherium laurillardi whose body masses must have reached between three and six tonnes, depending on the approach used (Casinos 1996, Fariña et al. 1998), and the mastodont Stegomastodon superbus, estimated at four or five tonnes. The mylodontid ground sloth Lestodon armatus (Fig. 1) follows closely, with an estimated mass of three to four tonnes, while the other my-lodontids are smaller: between one and two tonnes for Glossotherium robustum and Mylodon darwini, and with Scelidotherium leptocephalum, at 900 kg, falling just short of the megamammal category (Bargo et al. 2000). Among glyptodonts the largest was Glyptodon clavipes, individuals which may have reached nearly two tonnes. Others, though smaller, were also very large mammals: Doedicurus clavicaudatus at 1400 kg, Panochthus tuberculatus at (1100 kg), and Glyptodon reticulatus at 850 kg.
 Figure 1: Life reconstruction of Lestodon sp. The different size of the tusks suggest sexual dimorphism. Drawing by Néstor Toledo. Scale=100 cm.
Figure 1: Life reconstruction of Lestodon sp. The different size of the tusks suggest sexual dimorphism. Drawing by Néstor Toledo. Scale=100 cm.
There were also non-xenarthran giants, such as the camel-like Macrauchenia and the rhinoceros-like Toxodon (Fig. 2), whose body masses must have surpassed the one tonne limit (Fariña and Álvarez 1994, Fariña et al. 2005). The Carnivora, in turn, reached impressive sizes, although well below one tonne, with the sabre-tooth Smilodon populator and the short faced bear Arctotherium spp. attaining sizes of three or four hundred kilograms (Fariña et al. 1998).
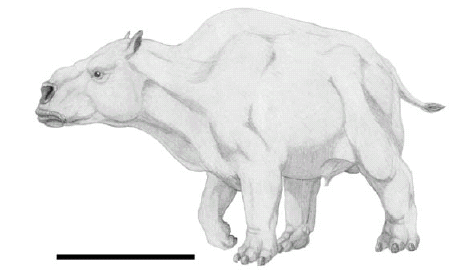 Figure 2: Life reconstruction of Toxodon sp. Drawing by Néstor Toledo. Scale=100 cm.
Figure 2: Life reconstruction of Toxodon sp. Drawing by Néstor Toledo. Scale=100 cm.
Limbs, locomotion and habits
Biomechanical studies performed in the last decade on large glyptodonts and ground sloths provided insight into the capacity of the limb bones to withstand bending forces, forearm extension and velocity, bipedalism or digging abilities. Within cingulates, Fariña's (1995) analyses of limb bones and locomotory habits in some glyptodonts indicated that femur strength indicators of large Pleistocene forms were equivalent to those of large living mammals capable of galloping (i.e. buffalos and rhinos), but values of the humerus were similar to those of elephants, which cannot gallop. The muscular insertions suggest that glyptodonts were able to adopt bipedal postures to perform strenuous activities, such as the intraspecific fighting proposed by Fariña (1995). For the ground sloths, different specializations may have been derived from a primitive quadrupedal way of locomotion in both main lineages of Pleistocene forms, megatheriids and mylodontids. The giant sloth Megatherium americanum has been formally proposed as bipedal based on ichnologic and biomechanical evidence (Aramayo and Manera de Bianco 1996, Blanco and Czerwonogora 2003). The latter includes analyses of body size, speed, Froude number, indicator of athletic ability, bending and resistance moments of the vertebral column, as well as a complete geometric and biomechanical analysis of the footprints assigned to this species found in Pehuén-Có, Buenos Aires Province, Argentina. Bipedalism also implies that the forelimb could have been free to perform activities other than locomotion. Fariña and Blanco (1996) tested the possibility that the forearms of Megatherium americanum were designed for optimizing speed rather than strength of extension, and concluded that such a trait may have been associated with a potentially aggressive use of the animal's large claws. Bargo et al. (2000) analysed limb proportions and resistance to bending forces in mylodontids to infer their locomotor adaptations. The analysis indicates that some of them were well adapted for strenuous activities in which force was enhanced over velocity, such as digging. Based on this work, Vizcaíno et al. (2001) considered these taxa as possible builders of large Pleistocene burrows reported in the Pampean region (Zárate et al. 1998). As for the strange hoofed mammals collected by Darwin, their athleticism seems to have been impressive in the case of Toxodon, which must have been capable of fast locomotion, and even more impressive in Macrauchenia, albeit for different reasons. Indeed, limb bone strength of the latter was larger if measured transversely rather than anteroposteriorly, which has been interpreted as the capability of suddenly turning while being pursued by a predator (Fariña et al. 2005).
Feeding apparatus
Functional morphology and biomechanics have been applied to the study of feeding in a wide range of xenarthrans (Vizcaíno et al. 2008, and references therein). Within the Pleistocene armoured forms, plant-eating was determined in eutatines (Vizcaíno and Bargo 1998), pampatheres (Vizcaíno et al. 1998, De Iuliis et al. 2000) and glyptodonts (Fariña and Vizcaíno 2001), although different kinds of herbivory may have occurred in each group. The studies also revealed that some cingulates evolved mechanical solutions not present in any related taxa, and do not have current analogues that can be used as models to investigate and interpret adaptations of lineages without living representatives. For instance, the masticatory apparatus in glyptodonts underwent a telescoping process that placed it well below the cranium (Fariña 1985, 1988), creating problems in the way that stresses produced by mastication were absorbed by the mandible and implying unusual jaw mechanics (Fariña and Vizcaíno 2001). Bargo (2001), Bargo et al. (2006a, b), and Bargo and Vizcaíno (2008) studied the masticatory apparatus of the large South American Pleistocene ground sloths. Jaw mechanics, morphogeometric analyses, and the correlation between cranio-dental variables (hypsodonty, dental occlusal surface area and relative width of the muzzle) and diet, all suggested probable niche differentiation among ground sloths based on dietary categories. While the masticatory pattern of mylodontids is rather generalized with a clear anteromedial powerstroke, as previously proposed by Naples (1989), Megatherium americanum was well adapted for strong, mainly vertical biting. This information, in addition to tooth shape, suggests that teeth were mainly used for cutting, rather than grinding, and that fibrous food was not the main dietary component. Hypsodonty is the relative increase in crown height of a tooth. It has been traditionally viewed as a response to dietary shifts toward abrasive vegetation, although recent work indicates that evolution of hypsodonty is also due to the higher prevalence of grit and dust in more open environments (Bargo et al. 2006a and references therein). Bargo et al.'s (2006a) comparative analyses of eleven species of Pleistocene sloths suggest that differences in hypsodonty may be explained by diet, habitat and behaviour. Among mylodontids, hypsodonty was unlikely due solely to dietary preferences, such as grazing. As mentioned above, some mylodontids were capable diggers that likely dug for food, and ingestion of abrasive soil particles probably played a considerable role in shaping their dental characteristics. Geographical distributions of the megatheriids Eremotherium and Megatherium indicate differing habitats as possible factors in hypsodonty differences. Vizcaíno et al. (2006) investigated the relationship between dental occlusal surface area (OSA) and diet, and other biological factors in fossil xenarthrans. They found that for most fossil xenarthrans OSA is smaller than expected compared to extant herbivorous mammals of equivalent body size. Within xenarthrans, cingulates show the highest OSA values, suggesting more extensive oral food processing than in tardigrades. Among ground sloths, mylodontids have extremely low OSA values, suggesting low efficiency in oral food processing that was probably compensated for by high fermentation in the digestive tract, and/or lower metabolic requirements. On the other hand, Megatherium americanum has an OSA as high as, or even higher than, that expected for a mammal of its size, which indicates higher oral food processing, lower fermentation capacity, and/or higher metabolic requirements. Other features besides teeth are also important. For instance, Bargo et al. (2006b) used muzzle shape and facial musculature reconstructions to develop models of food intake in five species of South American Pleistocene giant ground sloths. Ground sloths with wide muzzles (Glossotherium robustum and Lestodon armatus) had a square, non-prehensile upper lip that, coupled with the tongue, were used to pull out grass and herbaceous plants (mostly bulk-feeders). Sloths with narrow muzzles (Mylodon darwini, Scelidotherium leptocephalum and Megatherium americanum) had a cone-shaped and prehensile upper lip (Fig. 3) that was used to select particular plants or plant parts (mixed or selective feeders).
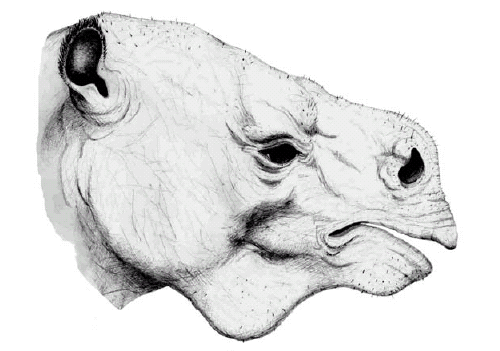 Figure 3: Life reconstruction of the head of Megatherium sp. Drawing by Néstor Toledo. Scale=50 cm.
Figure 3: Life reconstruction of the head of Megatherium sp. Drawing by Néstor Toledo. Scale=50 cm.
A morphofunctional approach has been less often applied to other Lujanian mammals because they are more readily comparable to living analogues. The upper incisors of Toxodon were strongly arched, whereas the lower ones were horizontally arranged. Their great lateral expansion gave the lower jaw a giant, spade-like appearance. In Macrauchenia the retracted position of the large, elliptical nostrils suggests the presence of a trunk. Considering morphological and isotopic evidence, they have been mainly considered as grazers and mixed-feeders respectively (MacFadden and Shockey 1997). Among horses, Hippidion shows a narrower nasomaxillar region, with well developed preorbital fossae and a retracted nasal notch, a combination of features that has been interpreted as an adaptation to more closed habitats, such as a savanah. Also, Hippidion shows teeth with relatively lower degree of folding in the enamel and less hypsodont than Equus (Amerhippus), which suggest a diet less rich in silica. Congruently, biogeochemical data suggest that the species of Equus (Amerhippus) had a more grazing diet than Hippidion (MacFadden et al. 1996, Mac Fadden and Shockey 1997, MacFadden et al. 1999).
PALAEOECOLOGY
As for palaeoecological interpretations, there have been several approaches to the study of Cenozoic South American faunas, including feeding habits, locomotion, and trophic relationships. Fariña (1996) analysed the trophic relationships of the South American Lujanian (late Pleistocene-early Holocene) megamammals, from the perspective of the ecological implications of their body sizes. Based on an estimation of population density derived from body sizes Fariña (1996), as Darwin realized 160 years before, emphasized that the fauna contained a significant diversity of large herbivores. Conversely, according to Fariña, it did not contain a proportionally diverse suite of large carnivores. Assuming a basal metabolism in agreement with the body size of the beasts, the food energy required to sustain the fauna was calculated. The results suggest that based on the requirements of the species under consideration (i.e., mammals over 10 kg), they alone must have needed about 1.8 megajoules per square metre per year (hereafter, MJ m-2 year-1) of the vegetation which sustained them. Since a primary productivity of 7.3 MJ m-2 year-1 is considered excellent for modern open field ecosystems, it is difficult to explain how the smaller species of mammals could have survived, let alone reptiles, birds, insects and other consumers. If the Lujanian plains had been as productive as the African savannah is today, about 38 MJ m-2 year-1, the consumption efficiency would fall to little more than 3%, which is a typical value for modern grassland systems. However, available evidence points to a different scenario. The fossiliferous Guerrero Member of the Luján Formation was deposited between about 20 to 10 thousand years before present, when the Last Glacial Maximum (LGM) was established and, due to the extensive glaciation in the Andes, the climate was much dryer and decidedly cooler than present conditions in that region (Clapperton 1983). Different sources of evidence are congruent with this paleoclimatic interpretation (Cantú and Becker 1988, Tonni 1990). As noted above, this was precisely Darwin's (1839) intuition about the landscape being less luxuriant than today. However, it seems that it was even more arid than surmised by the great naturalist. Current biogeographic reconstructions for the period of the LGM show that the Pampean plains underwent intense aeolian activity that redeposited large masses of silt and fine sand of periglacial origin. Southwest from the rivers where the sand was trapped in, a sand-sea in the southwestern half of the Pampas was formed, as well as a broad loessic belt over the remainder of the area. Also, the remains of the still-extant mammals, i.e., those whose habitat preference can be safely assigned, belong to species confined to Central and Patagonian faunistic provinces (Tonni 1985, Prado et al. 1987, Alberdi et al. 1989). The same can be said about the birds of this age (Tonni and Laza 1980), and analyses of pollen and ostracods have yielded congruent results (Quattrocchio et al. 1988, Markgraf 1989, Prieto 1996). According to Iriondo and García (1993), the shift was about 750 km south west relative to present conditions. Hence, the place where the current city of Luján lies would have had climatic conditions similar to the climate that exists currently in the northern Patagonian locality of Choele-Choel (39ºS, Río Negro province), whose climogram indicates a lower mean annual temperature (2.5-3ºC less than at the present), with more marked seasonality (summers only about 1ºC colder but winters up to 4ºC colder). More importantly, the aridity must have been higher, with rainfall considerably lower, about 350 mm per year as compared with the nearly 900 mm current for that area. These figures are considered as approximations, given the likely influence of other factors, such as the well-known high edaphic quality of the Pampean region, and perhaps due to the impact of the local water bodies, a topic discussed below. Thus, primary productivity in the mid-latitude Lujanian might have been higher on average than that in today's Choele-Choel area but it does not seem likely that it could have been higher than the most productive present-day cattle field of Uruguay, and, hence, it must have been dramatically lower than the African savannah. As a consequence of this reasoning, Fariña (1996) suggested that the coexistence of so many large herbivores in a poor environment led to strong competition for resources. It is worth to mention here that by the Pleistocene large carnivorous birds, like the phorusrhacids and theratorns had long before become extinct in the Pampean region. He concluded that some of the mammals previously considered strict herbivores might have been flesh-eaters to some degree. After eliminating large ungulates for varied morphological reasons, Fariña (1996) proposed that ground sloths were opportunistic carrion eaters. This challenging view, in turn, renewed interest in other ecological topics, such as niche partitioning in the Pleistocene and reinterpretation of the systematics of some South American Carnivora, among others. For instance, Vizcaíno's (2000) brief analysis of plant resource exploitation among sympatric Lujanian herbivorous armoured xenarthrans suggested that the main dietary difference among these cingulates was in the coarseness of the vegetation they were capable of processing. Bargo (2001) and Bargo et al. (2006b) proposed a niche differentiation among Lujanian ground sloths based on the different degrees of ability for plant selection due to muzzle morphology. Moreover, Vizcaíno et al. (2006) proposed that, like living sloths (see Gilmore et al. 2008 and references therein), mylodontids had very low metabolism, which suggests they were probably neither particularly abundant nor did they require as much food as originally calculated. A recent revision of the bears from South America proposed that during the Late Pleistocene there were three species of bears (Soibelzon 2004), instead of one as considered by Fariña (1996). Bears may have acted as large scavengers, which was probably true for other carnivores such as felids and canids as well, forcing reexamination of Fariña's (1996) estimates of trophic diversity, at least to a certain extent. Prevosti and Vizcaíno (2006) reviewed carnivore richness in the Lujanian of the Pampean Region, describing the palaeoecology of these species (including their probable prey choices) and assessing the available information on taphonomy, carnivore ecology, and macroecology to test the hypothesis of "imbalance" of the Río Luján fauna. They found that the carnivore richness of the Río Luján fauna comprises five species: Smilodon populator, Panthera onca, Puma concolor, Arctotherium tarijense, and Dusicyon avus, plus two other species that may be added when the Lujanian of Buenos Aires province is included: Arctotherium bonariense and Canis nehringi. With the exception of D. avus and A. tarijense, these are hypercarnivores that could prey on large mammals (100-500 kg) and juveniles of megamammals (>1000 kg). Smilodon populator could also hunt larger prey with body mass between 1000 and 2000 kg. The review of the "imbalance" hypothesis reveals contrary evidence and allows the proposal of alternative hypotheses. If high herbivore biomass occurred during the Lujanian, a higher density of carnivores could be supported.
EXTINCTION
The study of extinctions has become a particularly relevant issue in palaeontology. Over the last few decades, analyses of biodiversity during the history of life and its ups and downs, both gradual and sudden (Sepkoski 1978, 1979, Raup and Sepkoski 1984) have become the cornerstone of the way we look into the deep past. Moreover, the spectacularity of mass extinction and of the non-actualistic proposals of extraterrestrial causes (Álvarez et al. 1980) attracts the attention of academics as well as of the general public. The extinction of the giant mammals has long been attributed to the purported competition that followed the interchange of mammalian contingents across the Panama isthmus when it emerged some three million years ago. Known as the Great American Biotic Interchange, this asymmetrically reciprocal invasion brought carnivores (cats, dogs, mustelids, and bears), insectivores, rodents (in addition to the already present caviomorphs), rabbits, mastodons, tapirs, horses, camels, deer and peccaries to South America, where many of those groups still thrive. On the other hand, some South American lineages trekked north across the bridge: opossums, caviomorph rodents, toxodonts, glyptodonts, armadillos (including pampatheres), anteaters and ground sloths, and platyrhine primates, although not all were equally successful or ventured equally far in their new land. According to a proposal long held for the first half of the 20th century (Matthew 1930, Webb 1976, Simpson 1950), the evolutionarily advantaged northern mammalian lineages, on their triumphal path from their alleged evolutionary cradle in central Asia to all the corners of the globe, outcompeted their South American counterparts, driving them to extinction or forcing them to take refuge in marginal habitats. In the last part of that century, some authors critically revised the success of the North American immigrants in South America (Marshall et al. 1982, Marshall 1988, Webb 1985), and Webb (1991) provided an ecogeographic model that explains the assymmetrical results of the land-mammal interchange between both land masses. An analysis of the pattern of Pleistocene extinction (Lessa and Fariña 1996) revealed that large body size, rather than continent of origin, was the leading factor in determining which mammals went extinct. Another interesting issue is that of human impact, as it relates both to the timing of the peopling of the Americas and to ethical, environmentally oriented issues. Up to present, evidence of this human impact was scarce, although some remains do show interesting connections. Particularly, a clavicle of a mylodontid ground sloth found in Arroyo del Vizcaíno, Uruguay, shows marks assigned to human activities. This specimen, as well as others, have been dated at about 29,000 years before present, a much older age than the 12,000 or 13,000 ybp currently accepted for human presence in the Americas (Arribas et al. 2001, Fariña and Castilla 2007).
ANSWERS TO DARWIN'S QUESTIONS
As noted above, we intend here to overview the palaeobiological and palaeoecological contributions on the Pleistocene mammals of South America and assess their measure of progress in answering young Darwin's questions and comments on such matters. As conclusions of this article, his assertions will be considered one by one and our current point of view will be added. "We may therefore conclude that the whole area of the Pampas is one wide sepulchre for these extinct quadrupeds", said Darwin, and time has corroborated this statement in its fullest sense. Beginning with the collections made by Francisco Muñiz and Da'maso Larrañaga, as well as the subsequent work of the Ameghino brothers, the Museo de La Plata, the Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", the Museo Nacional de Historia Natural de Montevideo and a multitude of smaller museums exhibit exquisite specimens of those amazing fossil beasts, as well as of many other remains that are housed in collection rooms. "That large animals require luxuriant vegetation has been a general assumption, which has passed from one work to another. I do not hesitate, however, to say that is completely false; and that it has vitiated the reasoning of geologists, on some points of great interest in the ancient history of the world", claimed the young travelling naturalist. It seems he was absolutely right: as far as modern evidence is concerned, climates in middle latitudes seem to have shifted some 750 km towards the South-West since pleniglacial times, and hence the places where this impressive fauna must have lived then in a rather arid environment, as suggested by studies on small mammals, invertebrates, pollen and sediments, although more precise biogeographic and ecological hypotheses are yet in great need. "Mr. Owen... considers that [the remains of what today is called Macrauchenia] form part of an animal allied to the guanaco or llama..." This statement would have been right if referred to the fossil camelid Palaeolama. However, Macrauchenia is not a camelid but a Litoptern, a group not related to any living order of mammals and its phylogenetic relationships are yet to be established. Nevertheless it is worth to consider his reflection on the palaeoenvironments: "As all the existing members of the family Camelidae are inhabitants of the most sterile countries, so may we suppose was this kind…It is impossible to reflect without the deepest astonishment, on the changed state of this continent. Formerly it must have swarmed with great monsters, like the southern parts of Africa, but now we find only the tapir, guanaco, armadillo, capybara; mere pigmies compared to antecedents races... Since their loss, no very great physical changes can have taken place in the nature of the Country". Darwin's astonishment is to be shared. As is evident from the preceding sections, not so long ago, in geological terms, the mid latitude plains of south eastern South America were home to perhaps the most spectacular mammalian fauna of all times. How an environment of apparently such a low productivity could support so many herbivores remains a mystery. On the other hand, advancements in studies in astronomical forcing and palaeoclimatology partially refutes Darwin's impression, as the nature of the country has changed since its coldest period during the LGM (approximately 18 kybp) and sea level has risen more than 100 m, causing vast areas to become submerged. Ironically, the large beasts went extinct when the climate became less arid and the vegetation more luxuriant. That leads us to the following question: What then has exterminated so many living creatures? Extinction, a major topic in current evolutionary thinking was surprisingly not a major issue for Darwin. References to the subject are scarce, although important, in his writings and we are left to wonder whether he accepted the ideas common in his time of catastrophes (Cuvier's global revolutions) having wiped species from the Earth. On the other hand, his proposal, many years later as a settled gentleman living in the quiet comfort of his Kent home, that species were originated by means of natural selection has been interpreted as the foundation of modern views about extinction. In other words, species were easily viewed as prone to extinction if their individuals were not fit, if their members lost in great numbers the struggle for life. As for the giant mammals that inspired his thoughts as a young traveller, he was more than eager to accept they no longer lived, although the question why it may have happened did not surface frequently enough, probably accepting Cuvier's point of view on the subject. Our knowledge has increased considerably since then. Today, we pay enormous attention to this subject, due to our growing concern about present, dwindling biodiversity, our fascination with the spectacular extraterrestrial causes that have been proposed, and the ongoing human impact on megamammals. However, we end this article with one last reflection. "We are so profoundly ignorant concerning the physiological relations, on which the life, and even health (as shown by epidemics) of any existing species depends, that we argue with still less safety about either the life or death of any extinct kind" (Voyage of the Beagle, Chapter IX, Jan. 1834). Science has made considerable progress since those words were written by the person who changed the way humankind views itself and the rest of nature. However, we should be persuaded that his claim, which emphasized ignorance, be taken as a call to continue his approach. In this context, modern research on the marvellous beasts that roamed South America during the Pleistocene has indeed continued forward on the path set out by Darwin, contributing to the understanding of evolution intermingled with ecology and phylogeny. Thus, in the process of expanding our knowledge, as often happens in science, further ignorance has been revealed. However, we hope that this will only whet our appetite for the acquisition of more knowledge.
ACKNOWLEDGEMENTS
We want to acknowledge the editors for inviting us to participate in this volume. Gerry De Iuliis read critically an early version of the manuscript. Susana Bargo, Teresa Manera, Christine Janis and Darin Croft reviewed the manuscript. Néstor Toledo made the life reconstructions.
WORKS CITED IN THE TEXT
1. Alberdi, M.T., Menegaz, J.L., Prado, J.L. and Tonni, E.P. 1989. La fauna local de Quequén Salado - Indio Rico (Pleistoceno tardío) de la provincia de Buenos Aires, Argentina. Aspectos paleoambientales y bioestratigráficos. Ameghiniana 25: 225-236.
2. Alvarez, L.W., Alvarez, W., Asaro, F. and Michel, H.V. 1980. Extraterrestrial cause for the Cretaceous-Tertiary extinction: Experimental results and theoretical interpretation. Science 208: 1095-1108.
3. Andrews, P., Lord, J.M. and Evans, E.M.N. 1979. Patterns of ecological diversity in fossil and modern mammalian faunas. Biological Journal of the Linnean Society 11: 177-205.
4. Aramayo, S.A. and Manera de Bianco, T. 1996. Edad y nuevos hallazgos de icnitas de mamíferos y aves en el yacimiento paleo iconológico de Pehuen-Co (Pleistoceno Tardío), Provincia de Buenos Aires, Argentina. Asociación Paleontológica Argentina, Publicación Especial 4: 47-57.
5. Arribas, A., Palmqvist, P., Pérez-Claros, J.A., Castilla, R., Vizcaíno, S.F. and Fariña, R.A. 2001. New evidence on the interaction between humans and megafauna in South American. Publicaciones del Seminario de Paleontología de Zaragoza 5: 228-238.
6. Bargo, M.S. 2001. The ground sloth Megatherium americanum: skull shape, bite forces, and diet. In Vizcaíno, S.F., Fariña, R.A. and Janis, C. (eds.) Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica (Special Issue) 46: 41-60.
7. Bargo, M.S. and Vizcaíno, S.F. 2008. Paleobiology of Pleistocene ground sloths (Xenarthra, Tardigrada): biomechanics, morphogeometry and ecomorphology applied to the masticatory apparatus. Ameghiniana 45:175-196.
8. Bargo, M.S., Vizcaíno, S.F., Archuby, F.M. and Blanco, R.E. 2000. Limb bone proportions, strength and digging in some Lujanian (Late Pleistocene-Early Holocene) mylodontid ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20: 601-610.
9. Bargo, M.S., Vizcaíno, S.F. and Kay, R.F. 2004. Evidence for Predominance of Orthal Masticatory Movements in Early Sloths. 7° International Congress of Vertebrate Morphology, Journal of Morphology 260: 276.
10. Bargo, M.S., De Iuliis, G. and Vizcaíno, S.F. 2006a. Hypsodonty in Pleistocene ground sloths. Acta Paleontologica Polonica 51: 53-61.
11. Bargo, M.S., Toledo, N. and Vizcaíno, S.F. 2006b. Muzzle of South American ground sloths (Xenarthra, Tardigrada). Journal of Morphology 267: 248-263.
12. Barret, P. 1960. Transcription of Darwin`s first notebook on "Transmutation of species". Bulletin of the Museum of Comparative Zoology at Harvard College 122(6A): 248-296.
13. Blanco, R.E. and Czerwonogora, A. 2003. The gait of Megatherium Cuvier 1796 (Mammalia, Xenarthra, Megatheriidae). Senckenbergiana Biologica 83: 61-68.
14. Burkhardt, F. and Smith, S. 1985. The Correspondence of Charles Darwin Volume 1 1821-1836. Cambridge University Press, 752 p., Cambridge.
15. Cantú, M. and Becker, A. 1988. Holoceno del arroyo Spernanzoni, Dpto. Río Cuarto, Prov. Córdoba, Argentina. International Symposium Holocene in South America, Abstracts: 24.
16. Casinos, A. 1996. Bipedalism and quadrupedalism in Megatherium: an attempt at biomechanical reconstruction. Lethaia 29: 87-96.
17. Christiansen, P. and Fariña, R.A. 2003. Mass estimation of two fossil ground sloths (Xenarthra; Mylodontidae). In Fariña, R.A., Vizcaíno, S.F. and Storch, G. (eds.) Morphological studies in fossil and extant Xenarthra (Mammalia). Senckenbergiana Biologica 83: 95-101.
18. Clapperton, C. 1993. Quaternary geology and geomorphology of South America. Elsevier, 779 p., Amsterdam.
19. Darwin, C.R. 1839. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Volume III journal and remarks 1832-1836. Henry Colburn Press, 615 p., London.
20. De Iuliis, G., Bargo, M.S. and Vizcaíno, S.F. 2000.Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution. Journal of Vertebrate Paleontology 20: 743-754.
21. Fariña, R.A. 1985. Some functional aspects of mastication in Glyptodontidae (Mammalia). Fortschritte der Zoologie 30: 277-80.
22. Fariña, R.A. 1988. Observaciones adicionales sobre la biomecánica masticatoria en Glyptodontidae (Mammalia; Edentata). Boletín de la Sociedad Zoológica (2a. época) 4: 5-9.
23. Fariña, R.A. 1995. Limb bone strength and habits in large glyptodonts. Lethaia 28: 189-96.
24. Fariña, R.A.1996. Trophic relationships among Lujanian mammals. Evolutionary Theory 11: 125-34.
25. Fariña, R.A. and Álvarez, F. 1994. La postura de Toxodon: una nueva reconstrucción. Acta Geologica Leopoldensia 39: 565-571.
26. Fariña, R.A. and Blanco, R.E. 1996. Megatherium, the stabber. Proceedings of the Royal Society London, Series B 263: 1725-1729.
27. Fariña, R.A. and Castilla, R. 2007. Earliest evidence for human-megafauna interaction in the Americas. In Corona, M.E. and Arroyo-Cabrales, J. (eds.) Human and Faunal Relationships Reviewed: An Archaeozoological Approach, British Archaeological Reports, International Series 1627: 31-34, Oxford.
28. Fariña, R.A. and Vizcaíno, S.F. 2001. Carved Teeth And Strange Jaws: How Glyptodonts Masticated. In Vizcaíno, S.F., Fariña, R.A. and Janis, C. (eds.) Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica (Special Issue) 46: 87-102.
29. Fariña, R.A., Vizcaíno, S.F. and Bargo, M.S. 1998. Body mass estimations in Lujanian (Late Pleistocene-Early Holocene of South America) mammal megafauna. Mastozoología Neotropical 5: 87-108.
30. Fariña, R.A., Blanco R.E. and Christiansen, P. 2005. Swerving as the escape strategy of Macrauchenia patachonica (Mammalia; Litopterna). Ameghiniana 42: 751-760.
31. Fernicola, J.C., Vizcaíno, S.F. and De Iuliis, G. 2009. The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. Revista de la Asociación Geológica Argentina 64(1): 147-159.
32. Gilmore, D., Fittipaldi Duarte, D. and Peres da Costa, C. 2008. The physiology of two- and three-toed sloths. In Vizcaíno, S.F. and Loughry, W.J. (eds.) The Biology of the Xenarthra, p. 130-142. University Press of Florida. Gainesville.
33. Herbert, S. 1980. The red notebook of Charles Darwin. Bulletin of the British Museum (Natural History) Historical Series 7: 1-164.
34. Huxley, J. and Kettlewell, H.B.D. 1965. Charles Darwin and his world. Viking Press, 144 p., New York.
35. Iriondo, M. and García, N.O. 1993. Climatic variations in the Argentine plains during the last 18.000 years. Palaeogeography, Palaeoclimatology, Palaeoecology 10: 209-220.
36. Lessa, E.P. and Fariña, R.A. 1996. Reassessment of extinction patterns among the late Pleistocene mammals of South America. Palaeontology 39(3): 651-662.
37. MacFadden, B.J. and Shockey, B.J. 1997. Ancient feeding ecology and niche differentiation of Pleistocene mammalian herbivores from Tarija, Bolivia: morphological and isotopic evidence. Paleobiology 23: 77-100.
38. MacFadden, B.J., Cerling, T.E. and Prado, J.L. 1996. Cenozoic Terrestrial Ecosystem in Argentina Evidence from Carbon isotopes of Fossil Mammal Teeth. Palaios 11: 319-327.
39. MacFadden, B.J., Cerling, T.E., Harris, J.M. and Prado, J.L. 1999. Ancient latitudinal gradients of C3/C4 grasses interpreted from stable isotopes of New World Pleistocene horses. Global Ecology and Biogeography 8: 137-149.
40. Markgraf, V. 1989. Palaeoclimates in Central and South America since 18,000 BP based on pollen and lake-level records. Quaternary Science Review 8: 1-24.
41. Marshall, L.G. 1988. Land Mammals and the Great American Interchange. American Scientist 76(4): 380-388.
42. Marshall, L.G., Webb, S.D., Sepkoski, J.J. Jr. and Raup, D.M. 1982. Mammalian Evolution and the Great American Interchange. Science 215(4538): 1351-1357.
43. Matthew, W.D. 1930. Range and limitations of species as seen in fossil mammal faunas. Bulletin of the Geological Society of America 41: 271-274.
44. Naples, V.L. 1989. The feeding mechanism in the Pleistocene ground sloth, Glossotherium. Contributions in Science, Los Angeles County Museum of Natural History 415: 1-23.
45. Paley, W. 1802. Natural Theology. Oxford University Press, 384 p., Oxford.
46. Prado, J.L., Menegaz, A.N., Tonni, E.P. and Salamme, M.C. 1987. Los mamíferos de la fauna local Paso Otero (Pleistoceno tardío), provincia de Buenos Aires. Aspectos paleoambientales y bioestratigráficos. Ameghiniana 24: 217-233.
47. Prevosti, F. and Vizcaíno, S.F. 2006. Paleoecology of the large carnivore guild from the late Pleistocene of Argentina. Acta Palaeontologica Polonica 51: 407-422.
48. Prieto, A.R. 1996. Late Quaternary Vegetational and Climatic Changes in the Pampa Grassland of Argentina. Quaternary Research 45: 73-88.
49. Raup, D.M. and Sepkoski, J.J. Jr. 1984. Periodicity of extinctions in the geologic past. Proceedings National Academy of Sciences 81: 801-805.
50. Quattrocchio, M., Deschamps, C., Martínez, D., Grill, S. and Zavala, C. 1988. Caracterización paleontológica y paleoambiental de sedimentos cuaternarios, Arroyo Napostá Grande, Provincia de Buenos Aires. 2º Jornadas Geológicas Bonaerenses, Actas: 37-46, Buenos Aires.
51. Radinsky, L.B. 1987. The Evolution of Vertebrate Design. The University of Chicago Press, 188 p., Chicago.
52. Reed, K.E. 1998. Using large mammal communities to examine ecological and taxonomic structure and predict vegetation in extant and extinct assemblages. Paleobiology 24: 384-408.
53. Sepkoski, J.J. Jr. 1978. Kinematic model of Phanerozoic taxonomic diversity 1: analysis of marine orders. Paleobiology 4: 223-251.
54. Sepkoski, J.J. Jr. 1979. Kinematic model of Phanerozoic taxonomic diversity 2: early Phanerozoic families and multiple equilibria. Paleobiology 5: 222-251.
55. Simpson, G.G. 1950. History of the fauna of Latin America. American Journal of Science 38: 361-389.
56. Soibelzon, L. 2004. Revisión sistemática de los Tremarctinae (Carnivora, Ursidae) fósiles de América del Sur. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 6: 107-133.
57. Tonni, E.P. 1985. Mamíferos del Holoceno del Partido de Lobería, Provincia de Buenos Aires. Aspectos paleoambientales y bioestratigráficos del Holoceno del sector oriental de Tandilia y Área Interserrana. Ameghiniana 22: 283-288.
58. Tonni, E.P. 1990. Mamíferos del Holoceno en la Provincia de Buenos Aires. Paula-Coutiana 4: 3-21.
59. Tonni, E.P. and Laza, J.H. 1980. Las aves de la Fauna local Paso de Otero (Pleistoceno tardío) de la provincia de Buenos Aires. Su significación ecológica, climática y zoogeográfica. Ameghiniana 17: 313-322.
60. Van Couvering, J.A.H. 1980. Community evolution in Africa during the Cenozoic. In Berensmeyer, A.K. and Hill, A. (eds.) Fossils in the Making, University of Chicago Press, p. 272-298, Chicago.
61. Vizcaíno, S.F. 2000. Vegetation partitioning among Lujanian (Late Pleistocene-Early Holocene) armored herbivores in the pampean region. Current Research in the Pleistocene 17: 135-137.
62. Vizcaíno, S.F. and Bargo, M.S. 1998. The masticatory apparatus of Eutatus (Mammalia, Cingulata) and some allied genera. Evolution and paleobiology. Paleobiology 24: 371-383.
63. Vizcaíno, S.F., Zárate, M., Bargo, M.S. and Dondas, A. 2001. Pleistocene burrows in the Mar del Plata area (Buenos Aires Province, Argentina) and their probable builders. In Vizcaíno, S.F., Fariña, R.A. and Janis, C. (eds.) Biomechanics and Paleobiology of Vertebrates. Acta Paleontologica Polonica, Special Issue 46(2): 157-169.
64. Vizcaíno, S.F., De Iuliis, G. and Bargo, M.S. 1998. Skull shape, masticatory apparatus, and diet of Vassallia and Holmesina (Mammalia: Xenarthra: Pampatheriidae). When anatomy constrains destiny. Journal of Mammalian Evolution 5: 293-321.
65. Vizcaíno, S.F., Fariña, R.A., Bargo, M.S. and De Iuliis, G. 2004. Phylogenetical assessment of the masticatory adaptations in Cingulata (Mammalia, Xenarthra). Ameghiniana 41: 651-664.
66. Vizcaíno, S.F., Bargo, M.S. and Cassini, G.H. 2006. Dental occlusal surface area in relation to food habits and other biologic features in fossil Xenarthrans. Ameghiniana 43: 11-26.
67. Vizcaíno, S.F., Bargo, M.S. and Fariña, R.A. 2008. Form, Function and Paleobiology in Xenarthrans. In Vizcaíno, S.F. and Loughry, W.J. (eds.) The Biology of the Xenarthra, University Press of Florida, p. 86-99, Gainesville.
68. Webb, S.D. 1976. Mammalian faunal dynamics of the Great American interchange. Paleobiology 2: 216-234.
69. Webb, S.D. 1985. Late Cenozoic mammal dispersal between the Americas. In Stehli, F.G. and Webb, S.D. (eds.) The Great American Biotic Interchange, Plenum Press, p. 201-217, New York.
70. Webb, S.D. 1991. Ecogeography and the great American interchange. Paleobiology 17(3): 266-280.
71. Woodward, J. 1987. Darwin. Editorial Alianza, 141 p., Madrid.
72. Zárate, M.A., Bargo, M.S., Vizcaíno, S.F., Dondas, A. and Scaglia, O. 1998. Estructuras biogénicas en el Cenozoico tardío de Mar del Plata (Argentina) atribuibles a grandes mamíferos. Revista de la Asociación Argentina de Sedimentología 5: 95-103.
Darwin at Puente del Inca: observations on the formation of the Inca's bridge and mountain building
Victor A. Ramos
Laboratorio de Tectónica Andina, FCEN, Universidad de Buenos Aires - CONICET. Email: andes@gl.fcen.uba.ar
ABSTRACT
The analyses of the observations of Charles Darwin at Puente del Inca, during his second journey across the High Andes drew attention on two different aspects of the geological characteristics of this classic area. Most of his descriptions on the characteristics and the origin of the natural bridge were not published, mainly due to his poor impression of Puente del Inca. However, the application of the uniformitarian principles shows that it was formed as an ice bridge associated with snow and debris avalanches later on cemented by the minerals precipitated by the adjacent hot-water springs. Darwin's observations on the complex structural section at Puente del Inca, together with his findings of shallow water marine fossil mollusks in the thick stratigraphic column of the area interfingered with volcanic rocks, led him to speculate on several geological processes. Based on his geological observations, Darwin argued on the mountain uplift, the subsidence of the marine bottom, the episodic lateral growth of the cordillera, and their association with earthquakes and volcanic activity, which was an important advance in the uniformitarian hypothesis of mountain uplift proposed by Charles Lyell. Darwin was able to recognize the episodic nature of mountain uplift, and based on these premises he concluded that the Andes were still undergoing uplift. Taken as a whole, his ideas anticipated in many years some of the premises of the geosynclinal theory, and current hypothesis on foreland migration of the fold and thrust belts.
Keywords: Andes; Subsidence; Volcanism; Mountain uplift; Lateral growth; Ice-bridge.
RESUMEN: Darwin en Puente del Inca: observaciones sobre la formación de Puente del Inca y el levantamiento de las montañas. El análisis de las observaciones geológicas realizadas por Charles Darwin en Puente del Inca, durante su segundo cruce de la alta cordillera de los Andes, atrajo la atención a dos diferentes aspectos de las características geológicas de esta clásica área. La mayor parte de sus descripciones sobre las características y el origen de este puente natural no fueron publicadas, principalmente debido a su pobre impresión sobre el Puente del Inca. Sin embargo, la aplicación de principios uniformitaristas muestra que fue formado como un puente de hielo asociado con avalanchas de nieve y detritos y posteriormente cementado por los minerales precipitados por las termas adyacentes de agua caliente. Las observaciones de la compleja sección estructural en las vecindades de Puente del Inca, junto con sus hallazgos de moluscos marinos fósiles de aguas someras en la potente columna estratigráfica del área intercalada con rocas volcánicas, lo condujeron a especular sobre diversos procesos geológicos. Basado en sus observaciones geológicas, Darwin argumentó sobre el levantamiento de las montañas, la subsidencia de los fondos marinos, el crecimiento lateral y episódico de la cordillera, y su asociación con terremotos y actividad volcánica, que fue un importante avance en las hipótesis uniformitaristas de levantamiento de montañas propuestas por Charles Lyell. Darwin fue capaz de reconocer la naturaleza episódica del levantamiento de montañas, y basado en estas premisas, concluyó que los Andes estaban todavía creciendo.Tomadas en conjunto, sus ideas se anticiparon en varios años a algunas de las premisas de la teoría geosinclinal y las hipótesis actuales de la migración de las fajas plegadas y corridas.
Palabras clave: Andes; Subsidencia; Volcanismo; Levantamiento de montañas; Crecimiento lateral; Puente de hielo.
INTRODUCTION
It is well established in the biological science community that Darwin's theory on the evolution of the species is a milestone in the comprehension of Life, and that this theory started to develop in Darwin's mind during his research and observations in South America and later consolidated after his return to Britain. However, his important ideas on the formation of the mountains and his hypothesis on the origin of the Puente del Inca are not well known (Inca's Bridge, see Figs. 1 and 2). It is well documented that he considered himself a geologist, receiving in his early days a strong influence of the uniformitarian ideas of Charles Lyell (Judd 1909). It is also important to state that after his Beagle journeys around the world, he contributed to improve and exerted a strong influence on Lyell's ideas. When we read that in 1838 he chose to characterize himself this way: "I a geologist" (Herbert 2005, p. 2), it should not come as a surprise that his monumental contribution - as stated in his Geological Observations in South America (Darwin 1846) - to the evolution of the Patagonian Coast and the High Andes is still a milestone in the comprehension of the geological history of these regions.
 Figure 1: Location map of Puente del Inca in the High Andes of Mendoza.
Figure 1: Location map of Puente del Inca in the High Andes of Mendoza.
 Figure 2: Present view to the southeast of Puente del Inca, High Andes of Mendoza. Note the thermal springs developed on the southern side where the ruins of the old hotel bath are preserved (photo by Marcelo G. Armentano).
Figure 2: Present view to the southeast of Puente del Inca, High Andes of Mendoza. Note the thermal springs developed on the southern side where the ruins of the old hotel bath are preserved (photo by Marcelo G. Armentano).
There are several contributions highlighting his geological thoughts, his ideas on mountain building, and his geological descriptions of the many different geological environments visited (see for example Judd 1909, Rhodes 1991, Herbert 2005, and references therein). In the present contribution I examine only his observations and his thoughts - first in the origin of the bridge, and later on his ideas on the uplift and deformation of the Andes - gained during his excursion to the Puente del Inca region.
ORIGIN OF PUENTE DEL INCA
The pioneer observations
The first mention of the natural bridge in the headwaters of Río Mendoza is attributed to Alonso de Ovalle in 1646. This natural bridge later drew the attention of several 19th Century travelers, who in their reports described what began to be called the Puente del Inca (Inca's Bridge). The earliest description was by Schmidtmeyer in 1820 to 1821, although the illustration was drafted by A. Aglio, a lithographer that had never seen the bridge. In spite of his unrealistic representation, the two major springs are depicted in the southern part of the valley (Fig. 3a, after Schmidtmeyer 1824). The second description was by John Miers in 1826, and he again related the bridge to the two thermal springs and presented the first analyses of the composition and temperature of the hot water (Fig. 3b). Darwin had a copy of Miers' (1826) book in the library of the Beagle, and followed his trail on his way back to Chile. Another picture of the bridge was published in a later edition of Darwin's Voyage (Darwin 1890, Fig. 4).

 Figure 3: a) First picture of Puente del Inca. Note the two thermal springs on the southern side of the valley as depicted by the lithographer A. Aglio (after Schmidtmeyer 1824); b) Puente del Inca and hot mineral springs described by Miers (1826). Note the ravines developed behind the thermal springs where nowadays are the ruins of the hotel baths (compare with Fig. 2).
Figure 3: a) First picture of Puente del Inca. Note the two thermal springs on the southern side of the valley as depicted by the lithographer A. Aglio (after Schmidtmeyer 1824); b) Puente del Inca and hot mineral springs described by Miers (1826). Note the ravines developed behind the thermal springs where nowadays are the ruins of the hotel baths (compare with Fig. 2).
 Figure 4: Puente del Inca as illustrated in a new edition of Darwin's Voyage (Darwin 1890).
Figure 4: Puente del Inca as illustrated in a new edition of Darwin's Voyage (Darwin 1890).
New samples of the thermal springs were obtained by another British traveler, Charles Brandt in 1827, and the chemical analyses were done by the famous physicist and chemist of that time Michael Faraday (Brandt 1827), whose results were analyzed by Darwin (1846). However the first geologic description and hypothesis on the origin of the bridge were proposed by Darwin, in spite of the poor impression that the bridge produced on him: "When one hears of a Natural bridge, one pictures to oneself some deep & narrow ravine across which a bold mass of rocks has fallen, or a great archway excavated. Instead of all this the Incas bridge is a miserable object" Darwin's Diary of the Beagle (1831-1835) reproduced by Keynes (2001). In his original field notebook he depicted a pencil sketch (Figs. 5a and b) and described "Incas bridge irregular hilly plain of valley filled up with pebbles & detritus a fan of ferruginous cellular Tufa covering a part: the river having cut as far as (x). continued to scoop out to the Southward; rubbish (B) fell down from plain I supported circled [page 202a] (m) whilst river, continued forming arch. - the oblique junction is very evident (horizontal & confined) plain generally horizontal gravel this not so; hence rubbish: - My hypothesis of Tufa is that it was deposited after valleys excavated & just before sea retired; matter before that generally deposited - hence Tufa from these Springs extends above ... in the slope, above their level: Springs hot - violent emission of gaz: Concretions, where water drips" (Darwin field notebook, pages 201-202a).
 Figure 5: Puente del Inca after Darwin: a) Reproduction of Darwin's pencil sketch from his field notebook, April 5th, 1835 (with the permission of English Heritage); b) Trace of the author from the original sketch (see explanation on the text).
Figure 5: Puente del Inca after Darwin: a) Reproduction of Darwin's pencil sketch from his field notebook, April 5th, 1835 (with the permission of English Heritage); b) Trace of the author from the original sketch (see explanation on the text).
The most complete description is preserved in the Diary of the Beagle (1831-1836) where he described "the bottom of the valley is nearly even & composed of a mass of Alluvium; on one side are several hot mineral springs, & these have deposited over tha pebbles [page 554] a considerable thickness of hard stratified Tufa; The river running in a narrow channel, scooped out an archway beneath the hard Tufa; soil & stones falling down from the opposite side at last met the overhanging part & formed a bridge. The oblique (D) junction of the stratified (A) rock & a confused mass is very distinct & this latter is different from the general character of the plain (B).- This Inca's bridge is truly a sight not worth seeing" (Darwin's The Diary of the Beagle 1831-1836, pages 553-554, see Fig. 6).
 Figure 6: Puente del Inca after Darwin: Small sketch in the margin with letter A, B, C, and D drawing in his Diary of the Beagle (1831-1836).
Figure 6: Puente del Inca after Darwin: Small sketch in the margin with letter A, B, C, and D drawing in his Diary of the Beagle (1831-1836).
Later on in Darwin's Journal of Researches he said "April 4th.- ...when one hears of a natural Bridge, one pictures to oneself some deep and narrow ravine, across which a bold mass of rock has fallen; or a great arch hollowed out like the vault of a cavern. Instead of this, the Incas Bridge consists of a crust of stratified shingle,[page 335] cemented together by the deposits of the neighbouring hot springs. It appears, as if the stream had scooped out a channel on one side, leaving an overhanging ledge, which was met by earth and stones falling down from the opposite cliff. Certainly an oblique junction, as would happen in such a case, was very distinct on one side. The Bridge of the Incas is by no means worthy of the great monarchs whose name it bears" (Darwin 1845, p. 334-335). In his geological observations he quoted "at this place (Puente del Inca), there are some hot and cold springs, the warmest having a temperature, according to Lieut. Brand (Travels, p. 240), of 91°; they emit much gas. According to Mr. Brande, of the Royal Institution, ten cubical inches contain forty-five grains of solid matter, consisting chiefly of salt, gypsum, carbonate of lime, and oxide of iron. The water is charged with carbonic acid and sulphuretted hydrogen. These springs deposit much tufa in the form of spherical balls. They burst forth, as do those of Cauquenes, and probably those of Villa Vicencio, on a line of elevation" (Darwin 1846, p. 190). As a common factor in all these descriptions it is clear that Darwin recognized that the river first excavated its channel, and later the thermal springs cemented the gravels. He also recognized that the bridge gravels where some kind of stratified crust in contrast with the northern side, where colluvial debris was falling from a disorganized deposit. He identified the obliquity between the river valley and the bridge, interpreting a younger age for the bridge. He accepted that concretions were formed were water drips, explaining the long stalactites descending from the roof of the bridge. He considered the origin of the bridge so simple that "this Inca's bridge is truly a sight not worth seeing". His sketches were not published until recently (Keynes 2001), and therefore his detailed descriptions were ignored in subsequent publications.
Modern proposals on the origin
Most of the subsequent visitors recognized - as Darwin did - the natural origin of Puente del Inca, with the only exception of Christiano Junior (1902). This author claimed that perhaps it was an old Inca bridge built with wood and rattan (bejuco, a climbing wild plant used by the Incas to make ropes), that after several centuries was cemented by the thermal springs and thus obtaining its present shape. Schiller (1907) described the deposits involved in the formation of the bridge, although his hypothesis on its origin is known from other people's references (see Sekelj 1947, Monteverde 1947). His hypothesis was that a crust growth from the thermal springs reached the opposite margin of the valley by means of slow lateral precipitation, a criteria accepted by Reichert (1924, 1929, p. 51). The bridge was again studied by Kittl (1941), who improved Darwin's original hypothesis. He postulated that the river was deflected southwards, increasing the incision that favored the slumps from the southern margin, later on cemented by the thermal springs. This proposal was refined by Monteverde (1967), who also supported the excavation of the river, but did not accept the lateral growth. He postulated that the gravels of the bridge were prior to its formation and were preserved from glacial erosion. He claimed that the excavation was enhanced by lateral erosion coming from the northern side, which deflected the river valley southwards. See the analyses of Ramos (1993) for further details in these hypotheses.
Present knowledge
To analyze the origin of Puente del Inca it is necessary to refer to the original ideas of Darwin and to apply Lyell's (1835) classical concept: "the present is the key of the past". From time to time, abundant and frequent snowfall during the winter is followed by the ENSO (El Niño Southern Oscillation) over the high subtropical Andes (Compagnucci and Vargas 1998). This abundant and frequent winter snowfall produces numerous ice bridges that last until the following summer, and sometimes even for two or three years. I had the opportunity to observe several ice bridges along the Horcones, Las Cuevas and Blanco rivers in the vicinities of the Puente del Inca region in the summer of 1983. The avalanches reached the opposite side of the valley producing a run-up (see Fig. 7).

 Figure 7: a) Ice bridge in the Río Blanco, a few kilometers south of Puente del Inca as seen in the summer of 1983. An avalanche during a hard winter remains for one or two years. Note the concentration of the gravel covering the ice and the run-up formed by the avalanche on the right side of the bridge; b) Partially collapsed ice bridge located upstream of Puente del Inca in the Río Cuevas, during the same field season.
Figure 7: a) Ice bridge in the Río Blanco, a few kilometers south of Puente del Inca as seen in the summer of 1983. An avalanche during a hard winter remains for one or two years. Note the concentration of the gravel covering the ice and the run-up formed by the avalanche on the right side of the bridge; b) Partially collapsed ice bridge located upstream of Puente del Inca in the Río Cuevas, during the same field season.
Thawing concentrated the gravels of the avalanche in the upper surface, and these gravels are supported in a mud matrix. This process hardens the gravel conglomerate and allows crossing these temporary bridges even with loaded mules. If that happens near a thermal spring, cementation of the gravels with precipitation of sulphates and carbonates would be possible, as observed in Puente del Inca. Puente del Inca is the only natural bridge of its type presently standing, but collapsed bridges have been observed in the headwaters of Río Plomo. In the sources of Río Morado de Las Toscas, a river that joins the southern margin of the Río Plomo one kilometer upstream of the Refugio Las Toscas, Padva (2000) described travertines and other deposits associated with thermal springs. There is evidence in these deposits that the travertines reached the opposite margin, but they are now collapsed. It is interpreted that during the last glaciation the headwaters of the Río Mendoza valley were covered by snow avalanches developing a series of ice bridges as the ones depicted in figure 7. After thawing, only those with an extra hardening were preserved, as the one cemented by thermal spring waters. Figure 8 illustrates the different stages in the development of Puente del Inca.

 Figure 8: Schematic evolution of Puente del Inca based on Ramos (1993) and Aguirre-Urreta and Ramos (1996). Note that when the avalanche is compacted and melting starts, the pebbles are concentrated in the upper part, where they are slowly cemented by the products of the thermal spring. Compare stage 2 and 3 with the ice bridges of figure 7.
Figure 8: Schematic evolution of Puente del Inca based on Ramos (1993) and Aguirre-Urreta and Ramos (1996). Note that when the avalanche is compacted and melting starts, the pebbles are concentrated in the upper part, where they are slowly cemented by the products of the thermal spring. Compare stage 2 and 3 with the ice bridges of figure 7.
As established by Rubio et al. (1993), the precipitation of carbonates and sulphates is controlled by the presence of cyanobacteria. These blue-green algae produce thin layers of carbonates coating the surface. The hot water flux is linked to the amount of seasonal rainfall in the region, which controls the temperature and the concentration of the spring waters. During the dry seasons, fluvial and aeolian erosion dominate over carbonate and sulphate precipitation, thus drying and cracking the bridge. These periods alternate with more humid ones that produce an intense coating of the bridge structure. A delicate equilibrium between erosion and precipitation, sometimes modified by anthropic activity, preserved the bridge until nowadays. In spite of Darwin's negative impression of Puente del Inca, the comprehension of its genesis is another good example that the "the present is a key of the past". No doubt that if Darwin would have seen an ice bridge he would have not hesitated in applying the uniformitarian ideas he defended - alongside Lyell - to understand the processes that formed the bridge.
ANDES MOUNTAIN BUILDING
There is no doubt on the importance that the journey across the Andes had for Darwin's geological thought. He stated in a letter to her sister: "I returned a week ago from my excursion across the Andes to Mendoza. Since leaving England I have never made so successful a journey...how deeply I have enjoyed it; it was something more than enjoyment; I cannot express the delight which I felt at such a famous winding-up of all my geology in South America. I literally could hardly sleep at nights for thinking over my day's work. The scenery was so new, and so majestic; everything at an elevation of 12,000 feet bears so different an aspect from that in the lower country...To a geologist, also, there are such manifest proofs of excessive violence; the strata of the highest pinnacles are tossed about like the crust of a broken pie" (Burkhardt and Smith 1985). Darwin's observations on the geology around the Puente del Inca region, when he examined the Mesozoic sections at both sides of the valley (Fig. 9) coming back to Valaparaíso, were important in several aspects of the development of his ideas on mountain building. In this area the complexity of the structure competes in impressiveness with the extraordinary exposures of the marine sequences interfingered with volcanic rocks. The stratigraphic sequence is repeated by a series of thrusts from Puente del Inca up to the drainage divide along the present border with Chile. Figure 10 shows Darwin's interpretation and the present understanding of that structure.
 Figure 9: Geologic map of Puente del Inca (after Ramos 1988 and Cegarra and Ramos 1996) with indication of some fossil localities (I-3, I-10) described in the text. Legend: Cat: Carboniferous slates; Trch: Triassic Choiyoi volcanics; Jurassic: Jlm: La Manga Formation carbonates where level No. 3 was described; Y: Auquilco Gypsum; Jt: Tordillo Formation redbeds; JKvm: Vaca Muerta Formation black shales; Cretaceous: Km: Mulichinco Formation redbeds; Ka: Agrio Formation limestones and sandstones; Kd and Kv: red sandstones and volcanic rocks; Most of these rocks are interfingered with volcanic flows and sills; Tsm: synorogenic Miocene deposits; Q: Quaternary Alluvium
Figure 9: Geologic map of Puente del Inca (after Ramos 1988 and Cegarra and Ramos 1996) with indication of some fossil localities (I-3, I-10) described in the text. Legend: Cat: Carboniferous slates; Trch: Triassic Choiyoi volcanics; Jurassic: Jlm: La Manga Formation carbonates where level No. 3 was described; Y: Auquilco Gypsum; Jt: Tordillo Formation redbeds; JKvm: Vaca Muerta Formation black shales; Cretaceous: Km: Mulichinco Formation redbeds; Ka: Agrio Formation limestones and sandstones; Kd and Kv: red sandstones and volcanic rocks; Most of these rocks are interfingered with volcanic flows and sills; Tsm: synorogenic Miocene deposits; Q: Quaternary Alluvium
Figure 10: a) Classical structural cross-section of the Cumbre Pass to the Uspallata valley drawn by Darwin (1846); b) Present interpretation of the structure based on Cegarra and Ramos (1996). Cerro Almacenes correlates with k k in Darwin's cross section.
Note how Darwin was aware of the complex structure of Río Horcones and the fine details in the classical Puente del Inca Section. He described in detail the anticline and the fault of Los Horcones as follows: "A little further on, the north and south valley of Horcones enters at right angles our line of section; its western side is bounded by a hill of gypseous strata [F], dipping westward at about 45°, and its eastern side by a mountain of similar strata [G] inclined westward at 70°, and superimposed by an oblique fault on another mass of the same strata [H], also inclined westward, but at an angle of only about 30°: the complicated relation of these three masses [F, G, H] is explained by the structure of a great mountain-range lying some way to the north, in which a regular anticlinal axis (represented in the section by dotted lines) is seen, with the strata on its eastern side again bending up and forming a distinct uniclinal axis, of which the beds marked [H] form the lower part"(Darwin 1846, p. 189). This is the overturned faulted anticline nowadays recognized in Los Horcones valley as shown in figure 10b. The observation of these sections influenced his geological thoughts in several central facts. I would like to emphasize the followings aspects.
Evidence of uplift
As Charles Lyell, Darwin was impressed by the localized uplift denoted by the evidence of variation of sea level through time. The example of the Temple of Serapis near Naples that was the frontispiece illustration of the "Principles of Geology" was clearly showing uplifts in the order of tens of meters, only affecting a small portion of the coast. But if we compare these small variations with what he was seeing during his journeys across the Andes - where marine "shells that once were crawling in the bottom of the sea" are now standing over 10,000 feet above sea level - we can understand his exciting comments. Puente del Inca was a key area for this observation by him. During his examination of the sedimentary and volcanic sequences he found remains of fossil shells in his level No. 3 a few hundred meters to the southeast of the bridge (see location (I-3) in Fig. 9), almost 3,000 meters above sea level (Darwin 1846). Some specimens fallen from the outcrop were identified as Gryphea, a typical benthic mollusk of Jurassic - Cretaceous age. He associated this uplift to some sort of injection of the volcanic rocks. The observation of fossiliferous strata interfingered with the volcanic sills, interpreted as evidence of submarine volcanism, matched early 19th Century ideas on mountain building. Darwin's paper "On the connexion of certain volcanic phaenomena, and on the formation of mountainchains and volcanoes, as the effects of continental elevations" (Darwin 1838) read in the Geological Society on the 7th of March, produced a deep impact in the members of the society. As already established by Rhodes (1991), he favored the slow formation of the mountain chains, based on the relation between the small elevations in the order of a few meters produced by his observed earthquake displacements of Concepción, and the supposed subsequent volcanic activity in the Osorno volcano. A few months later in the Presidential Address of 1839 Whewell - then the President of the Society - expressed the antagonism between the Uniformitarians and the dominant Catastrophists views of society at that moment. Darwin's ideas quoting that "the formation of mountain chains and volcanoes, which he (Darwin) conceives to be the effect of gradual, small, and occasional elevation of continental masses" contrast with the paroxysmal turbulence accepted in the current theories of that epoch (Whewell 1839, Rhodes 1991).
Evidence of subsidence
Another important contribution of Darwin derived from his observations in the Puente del Inca region was his analysis of the sea bottom subsidence. He claimed that "the fossils … from the limestone-layers in the whitish siliceous sandstone, are now covered … by strata, from 5,000 to 6,000 feet in thickness. Professor E. Forbes thinks that these shells probably lived at a depth of … 180 to 240 feet; … in this case, as in that of the Puente del Inca, we may safely conclude that the bottom of the sea on which the shells lived, subsided, so as to receive the superincumbent submarine strata: and this subsidence must have taken place during the existence of these shells; … The conclusion of a great subsidence during the existence of these cretaceo-oolitic fossils, may, I believe, be extended to other districts" (Darwin 1846). His reasoning on an active subsidence anticipated James Hall's similar ideas of 1857, who proposed - based on the subsidence inferred for the Appalachians' during Paleozoic times - that the great sediment load caused crustal failure and downwarp, opening the way to geosyncline theory (Hall 1857). Darwin used as reference the stratigraphic thickness between the first fossil bed at (I-3), and the youngest one located at (I-10) (see location in Fig. 9). The presence of several packages with similar fossil shells from the top to the bottom of the marine sequence was for Darwin clear evidence of a great subsidence. When combined with the observation of the thick conglomerates of the Tunuyán Formation these packages indicated to him a second, and more recent period of subsidence. This perception of renewed subsidence and subsequent uplift was the embryonic stage of his global mountain uplift hypotheses later discussed.
Evidence of lateral growth
As quoted by Giambiagi et al. (2009) clast provenance analyses of the synorogenic conglomerates of Miocene age provided the key for Darwin's claim for an episodic and lateral migration of the mountain uplift. The finding of the succession of marine fossils derived from the Cordillera Principal at the base, and clasts from the high grade metamorphic basement of Cordillera Frontal above in the sequence, clearly indicated to him the lateral and episodic growth of the Andes. This concept was analyzed in the same region many years later by Polanski (1964, 1972), who also arrived at the same conclusions about the foreland migration of the uplift during Andean deformation. This notion is part of the ideas about fold and thrust belt deformation as inferred in modern plate tectonics.
Association with volcanoes
In 1838 Darwin read at a meeting of the Geological Society perhaps the most important of all his geological papers, relating deformation, earthquakes, and mountain uplifts. After describing the great earthquakes which he had experienced in South America, and the evidence of their connection with volcanic outbursts, he proceeded to show that earthquakes originated in fractures, gradually formed in the earth's crust, and were accompanied by movements of the land on either side of the fracture (Darwin 1838, 1846). In conclusion he boldly advanced the view "that continental elevations, and the action of volcanoes, are phenomena now in progress, caused by some great but slow change in the interior of the earth; and, therefore, that it might be anticipated, that the formation of mountain chains is likewise in progress: and at a rate which may be judged of by either actions, but most clearly by the growth of volcanoes." (Darwin 1838, pages 654-660).
CONCLUDING REMARKS
The region of Puente del Inca was very important in Darwin's geological observations and considerably contributed to his post-fieldtrip interpretations of the collected data. His work there can be summarized under two different aspects, i.e., the origin of the Puente del Inca and his contributions to mountain building.
a) Puente del Inca Although Darwin was not impressed at all with the natural bridge of Puente del Inca, he spent some time analyzing its origin. For him this had been a simple process associated with lateral growth and cementation aided by hot-spring water. Most of his observations were left unpublished in his field notebook and in his Diary of the Beagle (1831-1836). His basic ideas were right, but because they remained unpublished they were not considered for many years by later authors that hypothesized on the genesis of the bridge. The origin of the bridge is simple to understand when Darwin's ideas on uniformitarian processes are combined with actual observations of hard winters with exceptional snowfall, such as in those seasons preceding the El Niño Southern Oscillation. The sequence of processes involved in the formation of the bridge is straightforward, if classical uniformitarian concepts are applied. First it was formed as an ice bridge, probably during the last maximum glacial; the ice was covered by avalanches debris from the adjacent slope; then during thawing the bridge was supported by the mud matrix of the conglomerates, which subsequently were cemented by the minerals of the hot-thermal springs.
b) Mountain building The observations that led Darwin to infer gradual, small, and episodic mountain uplift combined with periods of important subsidence, together with the perception of lateral growth of a mountain chain to the foreland, are the base of modern concepts in orogeny. The evidence connecting episodic earthquakes and volcanic activity, partially seen by Darwin, but also mentioned to him by some other witnesses, were complementary to understand the processes associated with the uplift of the Andes. However, the notion that the uplift was produced by a succession of earthquakes, an active process still ongoing, was the clue for the comprehension that the Andes are still undergoing uplifting, and one of the best examples of application of uniformitarian ideas in tectonics. It is easy to understand the satisfaction of Charles Lyell with the new evidence drawn from Darwin's observations, and the close friendship developed by these two scientists in the Geological Society, that at that time was dominated by Catastrophists. There is no doubt now that Darwin's and Lyell's ideas slowly pervaded the geological community, and that in those early years Darwin was a geologist trained mainly by several years of fieldwork and observations obtained during his research in Argentina, where the Andes played a central role in his hypotheses.
ACKNOWLEDGMENTS
The author wants to acknowledge the access to the field notebooks to English Heritage and to Marcelo G. Armentano for his consent to use the panorama of Puente del Inca. Roberto Page, Carlos Costa, Suzanne M. Kay and Miguel Griffin contributed with their reviews to improve the preliminary version of the manuscript. Thanks are also due to the staff of the Laboratorio de Tectónica Andina for many years of cooperation and work in the region.
WORKS CITED IN THE TEXT
1. Aguirre-Urreta, M.B. and Ramos, V.A. 1996. Áreas de interés. In Ramos, V.A. et al., Geología de la Región del Aconcagua, Provincias de San Juan y Mendoza. Dirección Nacional del Servicio Geológico, Anales 24: 471-480, Buenos Aires.
2. Brandt, C. 1827. Journal of a voyage to Peru; a passage across the Cordillera of theAndes, in the winter of 1827, performed on foot in the snow; and a journey across the Pampas. H. Colburl, 346 p., London.
3. Burkhardt, F. and Smith, S. 1985. The Complete Correspondence of Charles Darwin. Vol. 1. 1821-1836. Cambridge: Cambridge University Press, 752 p.
4. Cegarra, M.I. and Ramos, V.A. 1996. La faja plegada y corrida del Aconcagua. In Ramos, V.A. et al. Geología de la Región del Aconcagua, Provincias de San Juan y Mendoza. Dirección Nacional del Servicio Geológico, Anales 24: 387-422, Buenos Aires.
5. Christiano Junior, J. 1902. En Los Andes. Diario La Provincia 1° de Marzo de 1902, Corrientes.
6. Compagnucci, R.H. and Vargas, W.M. 1998. Inter-annual variability of the Cuyo rivers' streamflow in the Argentinean Andean mountains and ENSO events. International Journal of Climatology 18(14): 1593-1609.
7. Darwin, C. R. 1838. On the connexion of certain volcanic phaenomena, and on the formation of mountain-chains and volcanos, as the effects of continental elevations. Proceedings of the Geological Society 2: 654-660, London.
8. Darwin, C.R. 1845. Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. Beagle round the world, under the Command of Capt. Fitz Roy, R.N. John Murray, 2d edition, 519 p., London.
9. Darwin, C.R. 1846. Geological observations on South America. Being the third part of the geology of the voyage of the Beagle, under the command of Capt. Fitzroy, R.N. during the years 1832 to 1836. Smith Elder and Co., 280 p., London.
10. Darwin, C.R. 1890. Journal of researches into the natural history and geology of the countries visited during the voyage round the world of H.M.S. Beagle under the Command of Captain Fitz Roy. R.N. John Murray (A new edition with illustrations by R.T. Pritchett of places visited and objects described), 598 p., London.
11. Giambiagi, L., Tunik, M., Ramos, V.A. and Godoy, E. 2009. The High Andean Cordillera of central Argentina and Chile along the Piuquenes Pass-Cordón del Portillo transect: Darwin's pioneering observations compared with modern geology. Revista de la Asociación Geológica Argentina 64(1): 44-54.
12. Hall, J. 1857. Contributions to the geological history of the American continent. American Association for Advancement of Science, Proceedings 3: 29-69.
13. Herbert, S. 2005. Charles Darwin, Geologist. Cornell University Press. 485 p., Ithaca.
14. Judd, C.B. 1909. Darwin and Geology. In Seeward, A.C. (ed.) Darwin and Modern Science, 358 p., Cambridge.
15. Keynes, R.D. 2001. Charles Darwin's Beagle Diary. Cambridge Univesity Press. 464 p., Cambridge.
16. Kittl, E. 1941. El Puente del Inca: su formación y conservación. Revista Minera 12(3-4): 110-122, Buenos Aires.
17. Lyell, C. 1830-33. Principles of Geology. John Murray, Albemarle-Street, (1830) 1, 511 p.; (1832) 2, 330 p.; (1833) 3, 109 p., London.
18. Miers, J. 1826. Travels in Chile and La Plata, including accounts respectives the geography, geology, statistics, government, finances, agriculture, manners and constumes and the mining operations in Chile. Baldwin, Cradock and Joy, v. 1, 494 p.; v. 2, 532 p., London.
19. Monteverde, A. 1947. Origen del Puente del Inca. Revista La Ingeniería 866: 775-791, Buenos Aires.
20. Monteverde, A. 1967. Preservación del Puente del Inca. Revista Caminos 34(295): 28-36, Buenos Aires.
21. Padva, D. 2000. Geología del valle superior del Río del Plomo, provincia de Mendoza. Trabajo Final de Licenciatura, Universidad de Buenos Aires, (unpublished), 87 p., Buenos Aires.
22. Polanski, J. 1964 . Descripción geológica de la Hoja 25a Volcán San José, provincia de Mendoza. Dirección Nacional de Geología y Minería, Boletín 98: 1-94, Buenos Aires.
23. Polanski, J. 1972. Descripción geológica de la Hoja 24a-b Cerro Tupungato, provincia de Mendoza. Dirección Nacional de Geología y Minería, Boletín 128: 1-110, Buenos Aires.
24. Ramos, V.A. 1988. The Tectonics of the Central Andes: 30° 33°S latitude. In Clark, S., Burchfiel, D. and Suppe, J. (eds.), Processes in Continental Lithospheric Deformation, Geological Society America, Special Paper 218: 31-54.
25. Ramos, V.A. 1993. Geología y estructura de Puente del Inca y el control tectónico de sus aguas termales. Simposio sobre Puente del Inca. 12° Congreso Geológico Argentino and 2° Congreso de Exploración de Hidrocarburos (Mendoza), Actas 5: 8-19.
26. Reichert, F. 1924. Sobre la radioactividad de las fuentes termales de Puente del Inca y Cacheuta. 1° Congreso de Química, 2, Sección Científica: 55-56, Buenos Aires.
27. Reichert, F. 1929. La exploración de la Alta Cordillera de Mendoza. Biblioteca del Oficial, vol. 138-139. 1 tomo, 1-401, Buenos Aires.
28. Rhodes, F.H.T. 1991. Darwin's search for a theroy of the earth: symmetry, simplicity and speculation. British Journal for the History of Science 24: 193-229.
29. Rubio, H.A., Santili, C.A. and Salomon, M.A. 1993. Puente del Inca: Restauración y preservación. 12° Congreso Geológico Argentino and 2° Congreso de Exploración de Hidrocarburos (Mendoza), Actas 5: 20-23.
30. Sekelj, T. 1944. Tempestad sobre el Aconcagua. Editorial Albatros, 287 p., Buenos Aires.
31. Schiller, W. 1907. Geologische Unterschungen bei Puente del Inca (Aconcagua). Vorläufige Mitteilung, Neues Jahrbuch für Mineralogie, Paläontologie und Geologie, Beilageband 24: 716-736.
32. Schmidtmeyer, P. 1824. Travels into Chile over the Andes, in the Years 1820 and 1821. Longman, Hurst and Rees, 378 p., London.
33. Whewell, W. 1839. Address to the Geological Society. Proceedings Geological Society, 3p., London.