Darwin, Coral Reefs, and Global Geology
Brian Roy Rosen
Darwin's coral reef subsidence theory derived from his broader interests in vulca-nicity and global geology. His reef observations were implicitly intended to demonstrate ocean floor subsidence. Although in retrospect his evidence for subsidence of atolls and barrier reefs and of ocean floors was insufficient, both ideas have survived the modern tests of boreholes and sea floor spreading. (Accepted for publication 16 March 1982)
To many people, Darwin's work on coral reefs probably seems like an esoteric geological digression from his much better known biological work. His study of coral reefs, however, is amongst his earliest publications, and of these publications, as many are devoted to geology as to biology. Indeed, in his diaries Darwin gives a strong impression of finding geology his most satisfying interest.
Darwin treated coral reefs in three published works. His first (1837) was a brief statement of his subsidence theory to the Geological Society of London. The second, in Journal and Remarks (1839), though brief too, also contains an account of the reefs he visited during the Beagle voyage. Darwin saw reefs at Tahiti, Cocos Keeling, and Mauritius, but only at Cocos Keeling was he able to make close observations, and then only for 12 days. His third and most important reef work is his main discourse, Coral Reefs (1842), this being the first and separately published part of his geological trilogy of the Beagle voyage. This and the Journal appeared in various subsequent editions, of which the second edition of Coral Reefs (1874) is probably the most important. To these three well known works must be added various letters (e.g., Darwin 1881b) and his manuscript, "Coral islands," posthumously published with editorial comments by Stoddart (1962). Stoddart believes that this was written in 1835 during the Beagle's voyage between Tahiti and
New Zealand (November to December 1835), and so precedes Darwin's main reef visit to Cocos Keeling. The origin of his subsidence theory goes back at least a few months further, however, because Darwin himself ([1887] 1958, p. 98) claimed that his "whole theory was thought out on the west coast of S. America before I had seen a true coral reef." He was last in S. America in September 1835, so we can say on his death centenary that his subsidence theory is now at least 147 years old.
Darwin's theory of coral reef formation is well known, simple, and original. He was able to bring together and explain in a single evolutionary theory the main kinds of reefs (fringing, barrier, and atoll) as successive subsidence stages on foundering volcanic island bases (Figure 1). He also used the geographical distribution of barrier reefs and atolls to show that they dominated the main ocean areas (Figure 2). These regions must therefore have foundered on a large scale, taking their reef-capped volcanic island passengers downward with them as they did so. Previous discussions of corals and reefs existed, but Darwin's was the first and arguably also the last successful attempt at a global synthesis.
Darwin's interest in coral reefs thus represents two geological preoccupations: global geology and the puzzle of how reefs, especially atolls, form. The biological content of his coral reef work, important as it is, is secondary to the geological side, because he needed it primarily to serve these geological ends. On the geological side, however, it seems to have become generally overlooked that global considerations, rather than reef considerations, were really the main generating force behind Darwin's subsidence theory. Priority of global interests is suggested by the same autobiographical passage already quoted, in which Darwin continues:
But it should be observed that I had during the two previous years been incessantly attending to the effects on the shores of S. America of the intermittent elevation of the land, together with the denudation and the deposition of sediment. This necessarily led me to reflect much on the effects of subsidence, and it was easy to replace in imagination the continued deposition of sediment by the upward growth of coral.
The title of his earliest coral reef publication (1837) is also instructive, while his Beagle manuscript (Stoddart 1962) notably concludes with mention of Humboldt's ideas of subsidence and uplift as complementary global processes. In

Figure 1. "If I am wrong, the sooner I am knocked on the head and annihilated so much the better I wish that some doubly rich millionaire would take it into his head to have borings made in some of the Pacific and Indian atolls ..." (Darwin 1881b). The "millionaire" who eventually obliged and first proved Darwin's subsidence beneath an atoll was the US Atomic Energy Commission, who drilled this hole (E-1) on Medren [Parry] 1d. on Eniwetak Atoll, Northern Marshall Is., in 1952. The site was found by a party led by Bill Morgan and marked by the author's temporary plaque in December 1976.
Rosen is with the British Museum (Natural History), London, England SW7 5BD. © 1982 American Institute of Biological Sciences. All rights reserved.
Coral Reefs itself, however, Darwin gives less stress to global geology until he draws everything together in chapter six. Here he says that the fact that atolls and barrier reefs indicate large scale subsidence of the ocean floors is 4 'perhaps the most interesting conclusion in this volume.'' Judd (1890) has pointed out that the appeal of geology to Darwin was, in Darwin's words, that it"creates the same grand ideas respecting this world which Astronomy does for the Universe." Darwin is in fact quite lyrical on the final page of Coral Reefs about this aspect of his subsidence theory, proudly referring to the tectonic implications of his coral reef map as "a magnificent and harmonious picture" (see also Figure 2 caption).
A further sign of the global background of Darwin's coral reef work is found in his geological trilogy of the Beagle voyage. This is not the disconnected set of titles it may at first seem, but the order in which he published them (Coral Reefs 1842, Volcanic islands 1844, South America 1846) gives less insight into the origin of his coral reef theory than if they are related to the route of the Beagle. His first visits were to Atlantic volcanic islands where he developed his ideas on elevation in relation to active vulcanicity and on subsidence of extinct volcanoes. He then spent more than three years in South
America, where amongst his many observations he found evidence for large scale crustal elevation and evidently began to consider possible evidence for complementary crustal subsidence. Where better for this than the ocean basins, vast negative topographic features, occupied as far as anyone knew at the time only by active and extinct volcanoes and by coral islands? Having deduced his coral reef theory from his elevation-subsidence paradigm, he was ready and able to study real reefs for the first time when he reached Tahiti.
Darwin needed to complete this cycle of ideas from the evidence of coral reefs themselves. He sought supporting evidence for subsidence from the way in which coral reefs were distributed with respect to their different kinds and to volcanoes. He needed to find whether coral reefs were indeed built, as others had argued, by shallow water organisms, and that they rose from deep foundations, and he needed to see if there were any features of coral islands that suggested they were actively subsiding.
REEF BIOLOGY
Darwin's (1842) biological objectives were confined to discussing the distribution of "polypifers" and their conditions of growth. With so little time to see reefs properly, and having already formulated his subsidence theory, he evidently concentrated on those things he most needed to know or check. Most important of these was to confirm that coral reefs were built by shallow water organisms, so his biological contribution is primarily ecological (in contrast to, say, Ehrenberg's reef studies [1834]). The answer to the coral reef riddle was determining how oceanic reefs apparently grow up from deep ocean floors when the only organisms that appear to construct them are shallow water dwellers. Clearly it was desirable to check on the thickness of coral reefs, their construction by organisms, and on the depth limits of these organisms to avoid going to the length of answering a nonexistent riddle.
Thus we have Darwin's account of Fitzroy's soundings on the ocean side of Cocos Keeling atoll, from which Darwin concluded that the depth limits of coral growth were about 20 fathoms (37 m). As on almost every other aspect of reefs in his book, Darwin then conscientiously ensured that he had working generalizations rather than isolated data by bringing together numerous observations by other people ("Captain Moresby tells me . . ."), and concluded that 30 fathoms (55 m) was actually the probable limit. The limit is now thought to be about twice this, depending on locality and region (Rosen 1977), but this increase on Darwin's figure over the succeeding 130
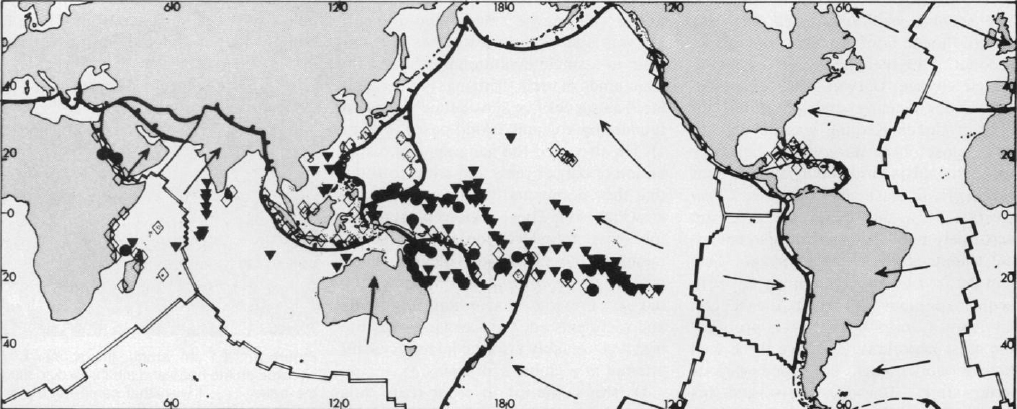
Figure 2. "We may thus, like unto a geologist who had lived his ten thousand years and kept a record of the passing changes, gain some insight into the great system by which the surface of this globe has been broken up, and the land and water interchanged" (Darwin 1845, p. 482). Lithospheric plate boundaries superimposed on Darwin's reef map (1842, less active volcanoes) show how his own views on global geology as supported by reef and volcano evidence (see quotation) fit with the modern plate tectonic view. Note predominance of subsidence reefs in mid-plate regions. For a modern version of Darwin's map, see Schuhmacher 1976.
Key
Subduction zones (destructive plate boundaries)
Transform and strike slip boundaries
Mid-ocean ridges (constructive plate boundaries)
Uncertain boundaries
Directions of plate motions
Fringing reefs (new, stable, or elevated reefs)
Barrier reefs (subsidence reefs)
Atolls (subsidence reefs)
BioScience Vol. 32 No. 6
years of reef research is negligible compared to the point that must have been uppermost in his mind, that of the much greater depth of the ocean floors (3000-5000 m).
Corals may not be the sole contributors to reef building, and perhaps other organisms contributed to the reef structure at depths below reef coral limits. Darwin anticipated this objection by reviewing the depth limits of other likely organisms and showing they also preferred shallow water. This now seems unconvincing, because it mostly reflects how little was known about the marine biology of deeper waters and biological oceanography. Nor is his survey based on what we would now consider to be an accurate knowledge of which organisms contribute to reefs. Darwin evidently regarded corals as the main agents, helped by "nullipores" (cementing red algae). We would now add at least the blue-green and green algae and possibly fora-minifera, sclerosponges, molluscs, and sea grasses. Subsequent discoveries of deep water coral banks (Buchanan 1886) and the generally richer amount of life in deeper water revealed during the Challenger voyage (and others since), especially the latter, led to the famous late nineteenth century phase of the "coral reef controversy" (Murray 1880). If Darwin had known of these discoveries his observations would then have needed careful qualification, so it seems that the lack of knowledge of deep water life actually favored the birth of his subsidence theory.
Developing still further what we should now recognize as the ecological side of his reef observations, Darwin also noted the asymmetrical distribution of organisms on reefs, which he ascribed largely to different degrees of exposure to prevailing wind and wave action. He believed that the most exposed seaward reefs were dominated by massive corals and cementing red algae ("nullipores"), and that these were responsible for the most active reef growth. From this and the inability of reef-building organisms to grow up above the surf spray zone, he was able to argue that reefs have an inherent tendency to grow outward once sea level constrains their upward growth.
Accounts of reef zonation have dominated the reef literature almost ever since (Stoddart 1969), and modern analysis of fossil reefs also depends heavily on this principle (Longman 1981). Darwin's original observations have not been seriously challenged, as subsequent research has only really filled in the habitat and taxonomic details. In fact the accumulating burden of all this detail has almost succeeded in obscuring whatever Darwin-type patterns really exist. The search for broader patterns of zonation was resurrected notably by Wells (1954, 1957) and followed by Rosen (1975, 1981a), Geister (1977) and Chappell (1980).
Finally, Darwin asserted something biological about reefs that has increasingly attracted attention to become almost the main reason for their biological study. He believed that coral diversity was highest where the "water is tranquil and the heat intense" (1842, p. 65). The temperature observation is apparently correct, but on a global oceanographic scale rather than on any one particular reef (Wells 1954, Rosen 1981a). For individual reefs, calmer water environments are certainly more diverse than the very roughest ones, but in contrast to the broad agreement about general zonation features, knowledge of diversity patterns of corals and other reef organisms is hindered by controversy over sampling methods (Head 1981). There are as yet no convincing explanations for the apparent patterns other than the strong correlation between diversity decrease, illumination decrease, and depth increase (Wells 1957, 1977, Rosen 1981a), and Connell's (1979) application of the intermediate disturbance hypothesis is the most attractive explanation so far for the diversity of particular reef environments.
The biological part of Darwin's coral reef work fulfilled his objectives by corroborating (not discovering) that reef building corals' were restricted to shallow water and establishing that reef builders grew most profusely near open water. His observations on coral zonation, though necessarily sketchy and less directly relevant to his main objective, are more original. They have led on naturally to the investigation of reef communities but still pose as yet unanswered questions about causes of community diversity patterns.
REEF GEOLOGY
Coral reefs are like icebergs; only a very small proportion of their bulk is visible above the water. Modern techniques allow us to overcome this a little. We can observe them directly underwater, drill into them, take portions back to laboratories for further studies, fly over them with satellite sensors, and conduct on-the-reef experiments, and yet we still feel that we do not adequately understand reef processes. What Darwin was able to conclude without such aids in his brief glimpse of reefs is therefore all the more remarkable. Much of Darwin's understanding was due to his well known ability to collect facts and to find patterns, but Geikie (1909), referring to Darwin's work on volcanoes, has remarked on his great gift for reconstructing sequences of geological events from his own observations. Making observations was always satisfying to Darwin, not just for its own sake, but because for him the observations chronicled geological change. Great changes on a grand time scale appealed to him most of all, and his study of coral reefs and their global implications illustrates Geikie's point perfectly. It is always clear why even the smallest detail is relevant, as with Darwin's almost incidental and correct observation of the sedimentological roles of parrot fish (e.g., Frydl and Stearn 1978) and holothurians (e.g., Orme 1977) as agents of fine sediment production (which also anticipates, by analogy, his much later study (1881a) of earthworms and soil formation).
Darwin's (1842) stated geological objectives were to describe the principal kinds of coral reefs and to explain the origin of "their peculiar forms," though this hardly does justice to the contents of Coral Reefs. Within this broader intention, he needed to confirm that organisms do actually build rock, that oceanic reefs do really have great thickness, that those reefs that he supposed originated by subsidence bore tangible evidence of it, and that, finally, such reefs occurred together in the ocean basins and away from active (elevating) volcanoes.
For the role of organisms in reef building, we find Darwin anticipating the study of carbonate sedimentology and lithification, now an economically important subject, in his observations of beach rock formation. He also collected specimens (1842) to show the stages of transition of living organisms into limestone (Rosen 1981b). He recognized the reef building role of encrusting red algae in building what have since been called "algal ridges" (compare Adey and Vassar 1975), and he presents a clear picture of framework growth by corals (compare Ginsburg and Lowenstam 1958). These features, together with surge channels, reef passes, and microatolls are now all recognized and understood in ways that can be traced back to Coral Reefs, probably without our always realizing it.
To satisfy himself on the subject of the thickness of coral reefs, Darwin used an old nautical device, that of projecting the shoreline profile beneath sea level, which could only have given a rudimentary estimate of sea floor depth at a given distance from shore, but this device had presumably won empirical credibility amongst seamen looking for moorings or fishing grounds. Darwin projected the shorelines of volcanic islands surrounded by barrier reefs down beneath water and reef top to establish the thickness of the reef. He also believed that a reef could be distinguished from its volcanic foundation by its much steeper (partly constructional) flanks. This approach to the thickness problem was the only one available to Darwin, and bearing in mind what we now know about complex effects of changing sea levels on limestone deposition and shallow water topography, it seems that he was a little lucky with this argument and evidence.
Stoddart (1976) also points out that Darwin was soon called upon to explain why no comparably great thicknesses of coral occurred on the continents, but was unable (with Lyell) to find a satisfactory reply. Darwin felt that the maximum one might actually expect was 5000 ft (1525 m). His guess was prophetic, because the first atoll drillings to successfully reach volcanic foundations actually passed through 4158 ft (1267 m) and 4610 ft (1405 m) of shallow water limestones. These are the celebrated Eniwetak boreholes of 1952 (see Figure 1 and Ladd et al. 1953, Schlanger 1963). The combined evidence of deep boreholes and seismic work on oceanic reefs since then points to a range of thicknesses between a few hundred meters and 2000 m (summary in Steers and Stoddart 1977).
In retrospective reply to the critical question leveled at Darwin, at least two well known continental fossil reef complexes, the Permian of Texas and New Mexico, and the Trias of the Dolomites, are about 1000 m thick (Wilson 1975) and are therefore certainly closer to oceanic reefs in thickness than any fossil reefs Darwin and contemporaries knew. These reefs, it must be admitted however, are very different from modern coral reefs, and the whole question is further complicated by differences of opinion on what constitutes a reef limestone.
The maximum likely amount of oceanic subsidence (exclusive of subduction) is about 3500 m (Parsons and Sclater 1977). To this can be added the maximum amplitude of sea level oscillations (400 m since the Oligocene, 500 m since the Triassic) (Vail and Hardenbol 1979, Vail et al. 1977), and 100 m for the depth limits of reef building organisms. In theory, therefore, one might expect to find an oceanic reef as thick as 4100 m depending on its age. In fact, the Bahama Banks, whose shallow water limestones are 4235 m thick and whose origins have been much debated, have recently been confirmed as an atoll-like structure (Schlanger 1981).
There is, therefore, an observed excess of oceanic reef limestone thicknesses compared with continental fossil reef limestones, and this is now best explained by supposing that oceanic reefs are nearly always lost, broken up, or metamorphosed in subduction zones and not preserved in recognizable form in continental sedimentary sequences. Darwin could hardly have foreseen this solution to the riddle of coral reef thicknesses.
The next part of Darwin's coral reef jigsaw to consider is his search for tangible evidence, for subsidence in the observable shallow water and land features of reefs. Not surprisingly, Darwin devoted much of Coral Reefs to this, discussing the recent history of changes to "reef islets," the depth of lagoons, the origin of faros (small annular reefs with contained lagoons resembling miniature atolls [Stoddart 1969 after Gardiner]), and the "disseverment" of atolls. It is ironic, however, that the modern acceptance of his subsidence theory does not finally rest on this, his most direct evidence, but on the posthumous fulfillment of his uncertain predictions of the thickness of atoll limestones. All his direct evidence was rendered inapplicable by the discovery, and more particularly by the implications of glacial changes of sea level. Daly (1910, and later papers in Steers and Stoddart 1977) is credited with this first and only fundamental breach of Darwin's original idea. There have since been numerous other authors who have articulated Daly's glacial control paradigm (summarized in Purdy 1974, and Steers and Stoddart 1977). Daly's own version is no longer acceptable in its original form, but implications and extensions continue to flow from it.
There has, for example, been an increasing number of theories about causes of sea level change other than directly glacial ones and much effort spent in trying to establish local and global sea level curves (Adey 1978, Vail et al. 1977). We cannot now overlook the probability that the uppermost 200 m of any reef structure measured from present sea level has been subaerially affected as a result of sea level changes within the Pleistocene and Tertiary (Vail and Hardenbol 1979). The same figures used above in discussing reef thicknesses also suggest that tectonic subsidence cannot be safely assumed unless a reef consists of more than 600 m of true reef deposits. In this way we can resolve Darwin and Daly's models by theoretically distinguishing first order features, due to Darwinian tectonic subsidence, from second order features, due to modern versions of Daly's glacial control.
Even so, it would be an oversimplification to say that sea level changes make nonsense of Darwin's observations. From the point of view of sedimentation at an individual site, foundation subsidence and sea level rise both represent marine transgressions, as he himself recognized (1881b). Since the last 20,000 years has been a time of marine transgression from a level of about -140 m relative to present (Adey 1978, Longman 1981), some of Darwin's observations, like those on "reef islets," have proved sound, even though he himself saw them in terms of foundation subsidence (Stoddart and Steers 1977).
To separate causes in explaining shallow features of reefs is difficult. To the above considerations must also be added the possibility of foundation uplift, even though the long-term trend might have been subsidence. Without any such complications of foundation movements, we know from sea level curves (Adey 1978, Longman 1981) that the uppermost part of any reef as measured to present day sea levels and whose history is older than 20,000 years, must previously have been subaerially exposed. The zone affected is about 140 m for structures that are between this age and 120,000 years. This has led various authors to suggest that modern reef structures are therefore resubmerged karstic features; Purdy (1974) has explored this most thoroughly. A further subtle complication lies in the fact that at 120,000 years, sea level was the same as at present, so it has been hard to distinguish reef features graded to this stillstand from those graded to present sea level. Those who specialize in fossil reef interpretation with all its recent economic applications have now recognized that modern reefs can only be used with great caution to provide uniformitarian analogues for fossil reef geometry. The implications for Darwin's theory are that if Purdy's karst antecedent version of Daly's paradigm is correct, though Adey (1978) contests this,
Darwin's three classic reef classes might no longer be an evolutionary progression explicable by subsidence (or any other transgression), even though subsidence is itself proven for oceanic reefs.
Knowledge of sea level oscillations and their likely effect on reefs also has important consequences for reef sedimentology. Darwin gave some especially interesting insights into reef geology that are not evidence for subsidence but predictions from it. In a long and intriguing footnote (1842, pp. 116-118) he attempts a facies and reef growth model that anticipates more recent sedimento-logical preoccupations of the same kind as discussed, for example, by Longman (1981). Darwin's model, based as it is on a single simple transgression, cannot now be regarded as universally applicable either to real fossil reefs or to the sequences in so-called modern reefs (which in reality may have histories as old as the Cretaceous). And yet the Darwinian model lives on almost as an ikon (e.g., the cover motif of Toomey's (1981) recent volume). In response to pleas such as that by Braithwaite (1973), it has taken more than 60 years to begin to develop reef models that are rigorous attempts to combine the Darwin and Daly paradigms (e.g., Adey 1978, Longman 1981), and move away from "century-old axioms that account for much existing confusion" (Adey 1978, p. 836).
Darwin certainly achieved his intention to describe the different kinds of coral reefs, but, in retrospect, his evidence for organic origin of reef rock is understandably incomplete, his evidence for great thickness of reef limestones very slender, and his evidence for foundation subsidence in the surface processes and morphology of reefs almost nonexistent. Nor do his famous reef classes necessarily represent intergradational stages. Yet potentially falsifying boreholes of the kind he could only dream about (1881b) have proved him correct 117 years after he first thought about subsidence of oceanic reefs (Figure 1).
GLOBAL GEOLOGY
Darwin used two lines of reasoning in his theory of reef subsidence; first, reefs that he believed originated by subsidence were not associated with active volcanoes, and second, these same subsidence reefs occurred together over large areas of the oceans. Subsidence reefs were, of course, atolls and barrier reefs, whereas fringing reefs could occur on static or rising foundations or represent recently initiated growth on a sinking foundation.
Stoddart (1976) has shown how pre-Darwin ideas about coral reefs had recognized their link with volcanoes, but thereby supposed them to be caused by uplift. This is turn failed to explain many other conspicuous coral reef features. From his earlier Beagle experience of volcanic islands (1844), Darwin had distinguished the association of active volcanoes with elevation and extinct volcanoes with subsidence. His famous map of coral reefs and active volcanoes (1842 and Figure 2, in part) revealed that subsidence reefs did not coincide with active volcanoes, but only with extinct ones. He was thus also able to maintain consistency between his ideas on reefs, volcanoes, and oceans.
The precise nature of coral foundations beneath atolls and oceanic barrier reefs remained unknown until very recently. Although atoll boreholes like those at Eniwetak confirmed the existence of sunken basaltic foundations, they do not explain the origin of this vulcanicity and its common characteristic of occurring in long chains spanning great distances across the oceans. Deep sea drilling results are now beginning to yield explanations and add broader based confirmation of Darwin's vulcanicity observation. In the case of the Hawaiian-Emperor Seamount chain, there are in fact two active volcanoes (Kilauea and Mauna Loa) at its eastern extremity, and there is now confirmation of extinct volcanoes of progressively increasing age and amounts of subsidence all the way to the northernmost Emperor seamount (Clague 1981). This pattern is matched by a reef sequence from fringing reef through barrier reef to atoll (not completely shown in Darwin's original map, Figure 2). Here we seem to have a perfect confirmation of Darwin's subsidence theory except for the presence of active volcanoes.
Active volcanoes can easily be accommodated by Darwin's subsidence theory if we simply take him literally. The islands closest to Hawaii's volcanoes bear only fringing reefs. If, on the other hand, we consider the whole chain to be a single feature, containing both subsidence reefs and active volcanoes, then plate tectonics helps explain this departure from Darwin's observation, and to distinguish this chain from the reefs and active vulcanicity of, say, Indonesia. Hawaii's vulcanicity is widely attributed to a hot spot within the Pacific plate (Wilson 1963, Morgan 1972), but there is also an alternative hypothesis of volca-nism along propagating fractures (Jarrad and Clague 1977). Most other volcanoes that rise up to sea level or higher occur in association with island arcs along destructive plate margins. As is now well established, the oceanic regions of lithospheric plates subside as they move away from their mid-ocean ridge sites of origin. Intra-plate volcanoes, once they are extinct, subside with their host plates, as well as in their own right (for sinking curves see Parsons and Sclater 1977, Detrick and Crough 1978). Without intra-plate vulcanicity there would therefore be no subsidence reefs at all across most of the ocean basins. Destructive plate margins, by contrast, are generally associated with fringing or raised reefs and uplift. Some of the nineteenth century objectors to Darwin's subsidence theory unwittingly chose reefs that, though apparently oceanic, are actually associated with destructive margins (e.g., Gup-py 1886 on the Solomon Is.).
Does the Hawaii-Emperor story apply to other oceanic reefs? Morgan (1972) believed that this chain might be a model for other major volcanic island and seamount chains, which in turn would have explained the origin of many other oceanic reefs, but this now seems to be doubtful. Clague (1981) and Schlanger (1981), summarizing deep sea drilling results, indicate that for both the Line-Tuamotu and Marshalls chains in the Pacific, their foundations of extinct volcanoes appear to have been simultaneously active, not made active sequentially as the Pacific plate passed over a mantle plume as Morgan envisaged for Hawaii-Emperor. At least from Darwin's point of view there is no current volcanic activity in these chains. These results of deep sea drilling are in this way reviving Menard's (1964) postulated Darwin Rise (Schlanger 1981), an ancient topographic volcanic high in the central Pacific, which he envisaged as having gradually foundered after being active, leaving subsidence reefs in its wake.
Neither the newly discovered pattern, nor the hot spot model can be assumed to apply to other important oceanic reef chains. There are also the isolated atolls to explain, like Aldabra, Bermuda, and Darwin's own Cocos Keeling. While we wait to discover the full range of origins of oceanic intra-plate, reef-surmounted volcanic features, we can at least confirm that Darwin was correct about the mutually exclusive occurrence of subsidence reefs and active volcanoes within the ocean basins. The few exceptions
have a volcanic character completely different from the elevating regions of destructive margins.
Darwin's coral reef map (Figure 2) was intended to reveal that if he was right about his subsidence theory of coral reefs, vast areas of the ocean floors must also have subsided (see solid symbols in Figure 2). To avoid circular reasoning between his reef theory and his ocean theory, Darwin needed independent evidence. He believed he had found this in the geological features of atolls and barrier reefs, but his observations here support only the idea of a marine transgression without unequivocally suggesting subsidence as a cause of this transgression. The combination of great reef thickness with the shallow water restriction of reef building organisms, though a less substantial part of Darwin's own argument at the time, has since proved to be the stronger part of his case. As he surmised (Figure 1 and 1881b), boreholes have been a much less ambiguous test of reef subsidence. The ocean basin distribution of guyots, sunken sea mounts previously truncated at sea level and unknown in Darwin's time, also provided good circumstantial evidence prior to borehole evidence, and unlike the few boreholes sunk up until now, guyots also depict a transoceanic pattern analogous to Darwin's reef map (Menard 1962).
In truth, the reef boreholes drilled to date do not strictly demonstrate ocean-wide subsidence, but merely the subsidence of the few reefs concerned. Until the 1960s, it must have seemed that only a vast program of reef drilling or seismic reef surveys could really confirm oceanic subsidence. Direct evidence was then suddenly provided by the ocean floors themselves in the form of sea-floor spreading, though few if any authors remarked on this at the time. This evidence for subsidence is independent of reef considerations because it is based on the measurement of sea-floor topography and on three different methods of measurement of sea floor ages (strati-graphic, isotopic absolute, and palaeo-magnetic). These provide the data for the empirical ocean floor subsidence curves quoted earlier. Darwin, therefore, guessed right (Figure 2) without having been able to do more than peer down into the depths as Fitzroy made his soundings off Cocos Keeling or walk about a few reef tops with his geological hammer and his "leaping pole."
There is, in any case, a difficulty in using coral reefs as direct evidence for ocean subsidence, which is that their geological and geographical patterns are not directly coupled to the history of the immediate ocean floor from which they rise. There is also considerable "noise" from the effects of sea level oscillations; they may even undergo relatively shortlived phases of uplift, attributed by McNutt and Menard (1978) to lithospheric flexure. These uplift features occur as fringing and raised reefs in the South Pacific (open symbols, Figure 2). But however and wherever they have originated within a plate, and whatever their local history, oceanic reefs ultimately become locked into the lateral and subsiding motions of their host plate. The empirical relationships between age, thickness, depth, and distance of reefs relative to features like mid-ocean ridges make it possible to make or test predictions about oceanic reefs, sea mounts, or the plate movements themselves and I imagine that Darwin would have found it immensely satisfying to see the frequency with which coral reefs appear as a source of data in oceanic geophysics.
For ultimate causes of ocean subsidence, Darwin was, however, almost mystical, believing it to be a "balancing process" for features like the mountain uplift he recognized in the Andes. Thus "as Sumatra rises, ... the other end of the lever [Cocos Keeling Atoll] sinks down" (1839). This is reminiscent of isostasy, a concept that was being refined by geologists before, during, and after Darwin's lifetime. Isostasy remains applicable to all vertical crustal movements, and Darwin's lever can be translated into the contrary vertical components of plate movements at destructive boundaries. Ocean floor subsidence itself, however, is currently attributed to gradual thermal contraction of ocean floor material as it spreads away from midocean ridges (Parsons and Sclater 1977), but we are not really any nearer to knowing why oceanic crust descends in subduction zones—whether it is pushed, pulled, or merely falls (Smith 1980). Menard's (1964, p. ix) comment, though now answered in one respect, lives on with new meaning: "It is evident that Darwin's hypothesis is no longer open to dispute. The cause of the subsidence is the hotly debated question of the day."
ACKNOWLEDGMENTS
The following people provided helpful discussion of the manuscript: Dr. Colin Braithwaite, Jill Darrell, Berta Rosen and Dr. Peter J. Smith. The map was drawn by Jim Darrell.
REFERENCES CITED
Adey, W. H. 1978. Coral reef morphogenesis: a multidimensional model. Science 202: 831-837.
Adey, W. H. and J. M. Vassar. 1975. Colonization, succession and growth rates of tropical crustose coralline algae (Rhodophyta, Cryptonemiales). Phycologia 14: 55-69.
Braithwaite, C. J. R. 1973. Reefs; just a problem of semantics? Am. Assoc. Pet. Geol. Bull. 57: 1100-1116.
Buchanan, J. Y. 1886. On oceanic shoals discovered in the S.S. "Dacia" in October 1883. Proc. R. Soc. Edinb. 13: 428-443.
Chappell, J. 1980. Coral morphology, diversity and reef growth. Nature (Lond.) 286: 249-252.
Clague, D. A. 1981. Linear island and sea-mount chains, aseismic ridges and intra-plate volcanism: results from DSDP. Soc. Econ. Paleontol. Mineral. Spec. Publ. 32: 7-22.
Connell, J. H. 1979. Tropical rain forests and coral reefs as open nonequilibrium systems. Pages 141-163 in R. M. Anderson, B. D. Turner, and L. R. Taylor, eds. Population Dynamics. The 20th Symposium of the British Ecological Society.
Daly, R. A. 1910. Pleistocene glaciation and the coral reef problem. Am. J. Sei. 30: 297-308.
Darwin, C. R. 1837. On certain areas of elevation and subsidence in the Pacific and Indian Oceans, as deduced from the study of coral formations. Proc. Geol. Soc. Lond. 2: 552-554.
______1839. Journal and Remarks 1832-1836. Vol. Ill in R. Fitzroy, ed. Narrative of the Surveying Voyages of His Majesty's Ships Adventure and Beagle. Henry Col-burn, London.
______1842. The Structure and Distribution of Coral Reefs. Being the First Part of the Geology of the Voyage of the Beagle, under the Command of Capt. Fitzroy, R.N. During the Years 1832 to 1836. Smith Elder and Co., London.
______1844. Geological Observations on the Volcanic Islands Visited during the Voyage of H. M. S. Beagle, Together with Some Brief Notices of the Geology of Australia and the Cape of Good Hope. Being the Second Part of the Geology of the Voyage of the Beagle, under the Command of Capt. Fitzroy, R.N. during the Years 1832 and 1836. Smith Elder and Co., London.
______1845. Journal of Researches into the Natural History and Geology of the Countries Visited during the Voyage ofH. M. S. Beagle Round the World, under the Command of Capt. Fitz Roy, R.N. Second edition. John Murray, London.
______1846. Geological Observations on South America. Being the Third Part of the Geology of the Voyage of the Beagle, under the Command of Capt. Fitzroy R.N. during the Years 1832 to 1836. Smith Elder and Co., London.
______1874. The Structure and Distribution
BioScience Vol. 32 No. 6
of Coral Reefs. Second edition Smith Elder and Co., London.
______1881a. The formation of Vegetable Mould, through the Action of Worms, with Observations on Their Habits. John Murray, London.
______1881b. [Letter to A. Agassiz] Pages 183-184 in F. Darwin, ed. 1887. The Life and Letters of Charles Darwin including an Autobiographical Chapter. Vol. III. John Murray, London.
______[1887] 1958. N. Barlow, ed. The Autobiography of Charles Darwin 1809-1882. Collins, London.
Detrick, R. S. and S. T. Crough. 1978. Island subsidence, hot spots and lithospheric thinning. J. Geophys. Res. 83: 1236-1244.
Ehrenberg, G. C. 1834. Über die Natur und Bildung der Corallenbänke des rothen Meeres und über einen neuen Fortschritt in der Kenntniss der Organisation im kleinsten Räume durch Verbesserung des Mikroskops von Pistor und Schick. Phys. Math. Abh. K. Akad. Wiss. Berl. (1832): 381-438
Frydl, P. and C. W. Stearn. 1978. Rate of bioerosion by parrotüsh in Barbados reef environments. J. Sediment. Petrol. 48: 1149-1158.
Geikie, A. 1909. Charles Darwin as Geologist. Cambridge University Press, Cambridge, England.
Geister, J. 1977. The influence of wave exposure on the ecological zonation of Caribbean coral reefs. Proceedings of the Third International Coral Reef Symposium 1: 23-30.
Ginsburg, R. N. and H. A. Lowenstam. 1958. The influence of marine bottom communities on the depositional environment of sediments. J. Geol. 66: 310-318.
Guppy, H. B. 1886. Notes on the characters and mode of formation of the coral reefs of the Solomon Islands, being the results of observations made in 1882-84, during the surveying cruise of H.M.S. "Lark." Proc. R. Soc. Edinb. 13: 857-904.
Head, S. M. 1981. The Ecology of Corals in the Sudanese Red Sea. Unpublished Ph.D. Thesis, University of Cambridge, England.
Jarrad, R. D. and D. A. Clague. 1977. Implications of Pacific island and seamount age for the origin of volcanic chains. Rev. Geophys. Space Phys. 15: 57-76.
Judd, J. W. 1890. Critical introduction. Pages 157-165 in C. R. Darwin, On the Structure and Distribution of Coral Reefs; also Geological Observations on the Volcanic Islands and Parts of South America Visited during the Voyage of H. M. S. Beagle. Minerva ed. Ward Lock, London.
Ladd, H. S., E. Ingerson, R. C. Townend, M. Russell and H. K. Stephenson. 1953. Drilling on Eniwetok Atoll, Marshall Islands. Am. Assoc. Pet. Geol. Bull. 37: 2257-2280.
Longman, M. W. 1981. A process approach to recognizing facies of reef complexes. Soc. Econ. Paleontol. Mineral. Spec. Publ. 30: 1-7.
McNutt, M. and H. W. Menard. 1978. Lithospheric flexure and uplifted atolls. J. Geophys. Res. 83: 1206-1212.
Menard, H. W. 1962. Foreword. Pages v-ix in C. R. Darwin (1842) The Structure and Distribution of Coral Reefs. University of California Press, Berkeley.
______1964. Marine Geology of the Pacific. McGraw-Hill Book Co, New York.
Morgan, W. J. 1972. Plate motions and deep mantle convection. Geol. Soc. Am. Mem. 132: 7-22.
Murray, J. 1880. On the structure and origin of coral reefs and islands. Proc. R. Soc. Edinb. 10: 505-518.
Orme, G. R. 1977. Aspects of sedimentation in the coral reef environment. Pages 267-306 in O. A. Jones and R. Endean, eds. Biology and Geology of Coral Reefs. Vol. IV: Geology 2. Academic Press, New York.
Parsons, B. and J. G. Sclater. 1977. An analysis of the variation of ocean floor bathymetry and heat flow with age. J. Geophys. Res. 82: 803-827.
Purdy, E. G. 1974. Reef configurations: cause and effect. Soc. Econ. Paleontol. Mineral. Spec. Publ. 18: 9-76
Rosen, B. R. 1975. The distribution of reef corals. Rep. Underwater Ass. N.S. 1: 1-16.
______1977. The depth distribution of Recent hermatypic corals and its palaeontolo-gical significance. Mem. Bur. Rech. Geol. Minier. 89: 507-517.
______1981a. The tropical high diversity enigma—the corals'-eye view. Pages 103-129 in P. H. Greenwood and P. L. Forey, eds. The Evolving Biosphere. British Museum (Natural History) and Cambridge University Press, Cambridge, England.
______1981b. Darwin's collection of corals and other reef-building organisms from the Cocos-Keeling Atoll. Page 23 in P. Whitehead and C. Keates. The British Museum (Natural History). Scala/Philip Wilson, London.
Schlanger, S. O. 1963. Bikini and nearby atolls, Marshall Islands. Subsurface geology of Eniwetok Atoll. US Geol. Surv. Prof. Pap. 260-BB: 991-1066.
Schlanger, S. O. 1981. Shallow-water limestones in oceanic basins as tectonic and paleoceanographic indicators. Soc. Econ. Paleontol. Mineral. Spec. Publ. 32: 209-226.
Schuhmacher, H. 1976. Korallenriffe; ihre Verbreitung, Tierwelt und Ökologie. BLV Verlagsgesellschaft, München.
Smith, P. J. 1980. Wegener's legacy. Open Earth 11: 8-9, 27.
Steers, J. A. and D. R. Stoddart. 1977. The origin of fringing reefs, barrier reefs, and atolls. Pages 21-57 in O. A. Jones and R. Endean, eds. Biology and Geology of Coral Reefs. Vol. IV: Geology 2. Academic Press, New York.
Stoddart, D. R. 1962. Coral islands by Charles Darwin with introduction, maps and remarks. Atoll. Res. Bull. 88: 1-20.
______1969. Geology and morphology of Recent reefs. Biol. Rev. Camb. Philos. Soc. 44: 433-498.
______1976. Darwin, Lyell, and the geological significance of coral reefs. Br. J. Hist. Sei. 9: 199-218.
Stoddart, D. R. and J. A. Steers. 1977. The nature and origin of coral reef islands. Pages 59-105 in O. A. Jones and R. Endean, eds. Biology and Geology of Coral Reefs. Vol. IV: Geology 2. Academic Press, New York.
Toomey, D. F., ed. 1981. European fossil reef models. Soc. Econ. Paleontol. Mineral. Spec. Publ. 30.
Vail, P. R. and J. Hardenbol. 1979. Sea level changes during the Tertiary. Oceanus 22: 71-79.
Vail, P. R., R. M. Mitchum, and S. Thompson. 1977. Seismic stratigraphy and global changes of sea level, Part 4: global cycles of relative changes of sea level. Am. Assoc. Pet. Geol. Mem. 26: 83-97.
Wells, J. W. 1954. Bikini and nearby* atolls: (2) oceanography (biologic). Recent corals of the Marshall Islands. US Geol. Surv. Prof. Pap. 260-1: i-iv, 385-486.
______1957. Coral reefs. Pages 609-631 in
J. W. Hedgpeth, ed. Treatise on marine ecology and paleoecology, Vol. 1. Ecology. Geol. Soc. Am. Mem. 67.
Wilson, J. L. 1975. Carbonate Facies in Geologic History. Springer Verlag, Berlin.
Wilson, J. T. 1963. A possible origin of the Hawaiian Islands. Can. J. Phys. 41: 863-870.