On some Alcyonaria in the Cambridge Museum. By SYDNEY J. HICKSON, M.A., F.R.S., Professor of Zoology in the University of Manchester.
[Read 7 March 1921.]
Clavuaria dura n.sp.
A very small specimen of a creeping Clavularia was found in the collection made by Dr J. C. Verco in 20-30 fathoms off Adelaide, S. Australia. The stolon consists of a few flat strands about 5 mm. in width attached to the horny tubes of a Gymnoblastic hydrozoon. On this stolon there stand at considerable distances apart five calices in the shape of inverted cones 1·5 mm. in height with a diameter of ·8 mm. at the distal end and 0·3 mm. at the end where it is attached to the stolon. At the free base of each cone (i.e. the distal end of the calyx) there are eight grooves radiating from the centre but the circumference of the cone is quite smooth.
The body wall of the stolen and of the calices is rendered perfectly rigid by a dense amalgamation of calcareous spicules, as in Tubipora, Telesto rubra end Sarcodictyon. So hard are the calices that I was unable to break them open With a pair of needles and it required a sharp blow on the cover-glass to crush them. When a calyx had been thus crushed the tentacles were seen to be armed with numerous spicules in the form of curved rods 0.1 mm. in length with a few small tubercles. The whole colony was pure white in colour. In form and habit this new species approaches very closely the Clavularia ramosa from the coast of Victoria* but differs from it in the hardness of the walls and the restriction of the eight grooves to the distal end of the calyx. Moreover in the new species I have not been able to find any of the double club spicules which are so characteristic of C. ramosa.
Sarcodictyon catenata Forbes.
An Alcyonarian having very much the same appearance as the British Sarcodictyon catenata of Forbes was sent to the Museum by Dr J. C. Verco from 20-35 fathoms off Adelaide, S. Australia, in 1904.
The interest of this specimen lies in the fact that the genus Sarcodictyon has been found hitherto only within the British Sea area.
In a former paper† I expressed agreement with Sars in suggesting that the genus should be merged with Clavularia and I
* Hickson, Trans. Zool. Soc. Lond. XIII. 1894, p. 310.
† Op. cit. p. 332.
said that the genus had been only imperfectly described. In doing so I did not do justice to the excellent description of Sarcodictyon catenata by Herdman* and I wish to make a sincere although belated apology. The submergence of the genus in Clavularia has unfortunately been accepted by May, Kükenthal and other writers, but since 1894 I have examined several species of Clavularia and by the kindness of Prof. Herdman several specimens of Sarcodictyon catenata, and I have come to the conclusion that it is desirable to retain the generic name Sarcodictyon. Sarcodictyon differs from all the species of Clavularia I have examinedexcept Clavularia dura—in having a stolon protected by hard inflexible walls of fused calcareous Spicules and in having long retractile anthocodiae which can be withdrawn into shallow convex calices situated on the strands of the stolon. The stolon consists of flattened strands about 1·5–2 mm. in width, forming a network slightly expanded at the nodes and in the places where the zooids are situated. The colour of the stolon seems to be almost invariably red, pink or yellow.
Clavularia dura appears to be a connecting link between the two genera in having hard inflexible walls but differs from Sarcodictyon in the large conical Calices and in the absence of colour.
The specimen from Australia is growing on a dead cockle shell about 35 mm. in length and breadth. The stolon is of a pale pink colour and forms a very irregular network of strands with meshes 5 or 6 mm. across, and the strands are about 1 mm. in width. As the zooids are all completely retracted it is very difficult to determine the exact distribution of the spicules in the anthocodiae but in these parts of the colonies free spicules can be found up to 0·2 mm. in length which have the form of irregular spindles provided with numerous irregular tubercles. The wall of the calices and of the stolon can be seen to be formed of spicules of the same form which in further growth have become jammed together to form a solid structure.
On comparing the specimen with specimens from the British sea area I can find no characters to separate it from S. catenata. It is true that the pale pink colour distinguishes it from all the other specimens I have seen; but as S. catenata is known to vary from red to yellow the colour character is obviously not reliable. There seems to me, therefore, to be no other course to adopt than to name this specimen S. catenata notwithstanding the enormous distance that separates the Australian from the British habitat.
*Roy. Phys. Soc. Edin. VIII, 1833, p. 31.
24—2
Pseudocladochonus hicksoni Versluys.
This very interesting Alcyonarian was first described by Versluys from the collections of the Siboga Expedition near ceram and Halmaheira*. It exhibits a curious similarity in its method of gemmation and general form to the fossils belonging to the family Auloporidae and particularly to the genus Cladochonus from the carboniferous strata, but according to Versluys, who has made an elaborate study of the genus and of the Auloporidae, it affords us an example of convergence rather than one of genetic affinity with the fossil family.
In a small collection of Alcyonaria from the Uraga Channel near Tokyo, Japan 40-200 fathoms, I found a few small specimens of this species which exhibit all the principal characters described in detail by Versluys.
The size of the retracted zooids 2 mm. x 1 mm. and the diameter of the stems from which they arise 1 mm. are the same as in the type.
The yellow bands where the stem is slightly constricted just above the origin of the zooids can be clearly seen in some of the older fragments but are obscure in the younger branches. Sections through the stem show an arrangement of solenia divided by septa similar to that of the type and the precise arrangement also seems to vary in different parts of the stem. In one of my sections there are four septa meeting in a central column, in another eight septa which do not meet in the centre, but show some fusions of their free borders. There can be little doubt that the 4-septate condition of the first-named section has been derived from an 13-septate condition by an increase in the thickness of the septal walls and fusion at the centre.
The small amount of the materiel at my disposal has prevented me from making a further investigation of this structure but sufficient has been done to prove that the identification of the genus is correct and that the specimens do not represent en aberrant species of the genus Telesto.
The only difference that I can observe between the Japanese and the Molluccan specimens is in the character of the spicules. In all the preparations I have made which show clearly isolated spicules or spicules which have not yet become firmly joined with their neighbours to form the solid wall or septa of the tubes, the spicules of the Japanese specimens possess more numerous and larger spines and tubercles than they do in the type specimens. The length of the larger spicules (c. 0·12 mm.) does not show any material difference but there is such a wide range of variation both in size and shape of the spicules in any one preparation, that
*Siboga Expedition. Monograph XIII c, 1907.
obviously it would be absurd to suggest a specific difference on this single very variable character.
In conclusion it may be said that the comments of Versluys on the relation of this Interesting genus to the family Telestidae appear to be sound as also is his conclusion that there is no evidence at present as to any direct relationship with the fossil genus Cladochonus and the Auloporidae.
Telesto trichostemma Dana.
A few branches of a Telesto that must be attributed to the widely distributed species T. trichostemma Were found in the Uraga Channel off Tokyo, Japan, in 40–200 fathoms. The walls of the main and lateral zooids are so densely crowded with large spindle-shaped and profusely tuberculated spicules that they are quite rigid and they show eight shallow longitudinal ridges as in other specimens of the species. These walls, however, become soft and flexible on prolonged boiling in potash and then the outline of the individual spicules can be clearly distinguished. In this respect the Species differs from T. rubra in which the walls are also rigid, but do not soften or show the outlines of spicules clearly after prolonged boiling in potash. The colour is pale red. An interesting feature of these of these specimens is that they support a number of specimens of the rare entoproctous polyzoon Barentsia discreta.
Leptogorgia sp.?
The specimen was obtained by Charles Darwin in the Galapagos Islands during the voyage of the "Beagle" in 1835. It is evidently only a small fragment of a much larger specimen.
The branches freely anastomose in one plane forming meshes of 6 sq. mm. or less but of very variable size. The branches are about 2 mm. in diameter and almost cylindrical in shape. They are of a dark red brown colour spotted on both sides with flat yellow calices.
The spicules are spindle-shaped with five or six encircling rows of prominent compound tubercles and have a length of about 0.1 mm. and a breadth, including the tubercles, of 0.04 mm.
The axis is composed of a horny substance without any deposit of calcium carbonate and is perforated longitudinally by a series of chambers filled with a light transparent spongy substance. In a branch of the axis 0.4 mm. in diameter 20 of these chambers can be counted in a millimeter of length.
No attempt has been made to identify the species of this gorgonid because the specimen is only a fragment and because I have not bad the opportunity of studying the systematic part of Kükenthal's monograph on the Gorgonaria in the "Valdivia" series of publications. Until this monograph or some other mono-
graph that gives a critical examination of the most unsatisfactory and confusing literature of the family, comes to hand, the attempt to identify the species is little better than a waste of time.
Virgularia mirabilis sp.? O. F. Müller.
In 1889 I recorded the occurrence of specimens of the genus Virgularia from the coast of Victoria under the name Virgularia löwenii, but as recent researches on the stages of growth of the northern species of the genus have shown that the type of this species is but a growth form of V. mirabilis, it is clear that the name I gave to the Australian specimen must be changed.
The question is whether these specimens, however, are correctly identified with a species that has hitherto been recorded only from the N. Atlantic and Mediterranean waters, and I have therefore re-examined the specimens in order to compare them with examples of V. mirabilis from our own coast. The result of this examination has been the failure to discover any satisfactory characters to distinguish them. In the number of the autozooids in the leaves (about 30), in the characters of the calices, and in the number and position of the single row of siphonozooids the Australian specimens resemble V. mirabilis and correspond with the description and figures of V. löwenii as given by Kölliker.
But before assuming that this identification is sound and that we have in this case an example of a species with bipolar distribution, it is really necessary to examine a large series of growth forms of the Australian Virgularias.
Many years ego specimens of Virgularia from Australia were recorded by Gray and named V. elegans, and A. Thomson has recently recorded a Virgularia under the same specific name from the Ceylon seas*. It is difficult to determine from the published descriptions of these specimens what are the specific differences between V. elegans, and V. mirabilis, as no account is given of the position and number of the siphonozooids. It seems possible, however, that they are all representatives of a species that has a very wide distribution where a suitable habitat occurs. For the present, therefore, I am content to leave the name V. mirabilis for the Victoria specimens.
Cavernularia chuni Kükenthal and Broch†.
Seven specimens of a species of Cavernularia were found by Dr C. Hose washed ashore after a storm on the beach at Miri, Sarawak, Borneo. As this is, I believe, only the second record of a pennatulid being "washed ashore"—the other being Cavernu-
* Report Pearl Fish., Suppl. XXVIII, 1905.
† "Valdivia," Pennatulacea, XIII, p. 190.
laria malabarica recorded by Fowler*—the fact is of some interest in the natural history of the sea-pens, and I was surprised to find that the species of the Bornean Cavernularia is not the same as that which is washed ashore in the Bay of Bengal.
The principal measurements of the five perfect specimens were as follows:
| 1 | 2 | 3 | 4 | 5 | |
| Length of rachis | 25 | 35 | 40 | 48 | 50 mm. |
| " stalk | 7 | 15 | 8 | 16 | 10 mm. |
From these figures it is clear that the stalk is relatively short, the rachis-stalk ratio varying from 2·5 : 1 in specimen 2 to 5 : 1 in specimens 3 and 5.
As in the type specimen, there is no sharp distinction in diameter between the stalk and rachis, the stalk passing abruptly into the rachis by the appearance of the zooids only. This feature is in marked contrast to that of C. malabarica—in which the passage from the stalk to rachis is marked by a great increase in diameter.
The specimens are so much contracted and distorted that any figures that might be given of the diameter would be untrust-worthy. As a guide, however, to the proportions in the species, I may say that I estimated the greatest diameter of the rachis of specimen 5 to be about 15 mm. There is no axis in the two specimens that were dissected.
The spicules of the rachis are needles 0·3–0·5 mm. in length by 0·04 mm. in breadth arranged vertically to the surface and penetrating down almost to the centre of the rachis. There are no spicules that are divided at the extremities.
In the stalk the spicules at the surface are small rods and oval in shape, ·05–0·1 mm. in length, but in the depths there are numerous rod-shaped spicules of the same type as those that occur in the rachis.
The character and arrangement of the spicules are like those described for Cavernularia chuni by Kükenthal and Broch, with which species the Bornean specimens also agree in the total absence of an axis.
In the type, however, the stalk is relatively much longer, the stalk-rachis ratio being 1–1·4.
As the species was founded on a single specimen and this stalk-rachis ratio may be variable, it seems probable that the specimens from Borneo should be called Cavernularia chuni.
The locality given for the type which is deposited in the Vienna Museum is Coamong (?). (I do not know where Coamong is and cannot find such a place mentioned in Stieler's Atlas.)
* P.Z.S. 1894.
Cavernularia darwinii n.sp.
In the stores of the Cambridge Zoological Museum there is a specimen of the genus Cavernularia from C. Darwin's "Beagle" collection labelled Chatham Island. Galapagos Islands, September 1835. As I can find no record of any species of this genus in the Eastern Pacific Ocean and as this specimen is of special interest from its association with the great English naturalist and his memorable voyage in the "Beagle" I decided to examine it carefully with a view to giving it a definite specific name.
The specimen is unfortunately not very well preserved and is strongly contracted and bent, but there are three autozooids killed expanded, one of which has been mounted as a preparation for the microscope.
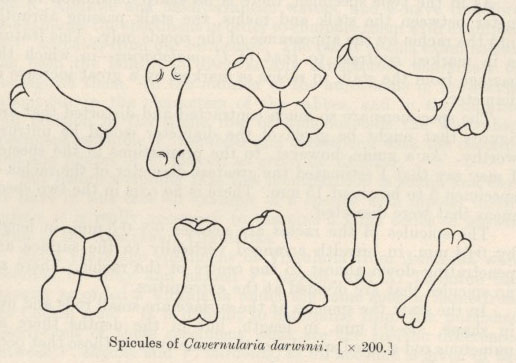
Allowing for the contraction and bends the specimen is about 90 mm. in length, the rachis being 38 mm. and the stalk 12 mm. and therefore the stalk-rachis ratio about 1–3.
It is quite impossible to make any accurate statement about the arrangement of the zooids on the rachis or of the relative number of autozooids and siphonozooids owing to the extremely contracted and convoluted condition of the surface. The only statement that can be made without having recourse to a large series of sections is that apparently there are relatively very few autozooids.
The presence of an axis can be proved by probing with a needle and so far as can be judged by this method it extends about half-way up the rachis and about half-way down the stalk. The
general statement may be made therefore that the specimen has an incomplete axis.
The spicules of the rachis, although very variable in shape, are at the same time, very characteristic and can be easily distinguished from the rachis spicules of any other species I have examined. The most prevalent type is that of a short rod 01–0·13 mm. in length, terminating in swollen extremities divided into two, three, four or sometimes five convex facets having an appearance which is extraordinarily like that of a metacarpal bone of a mammal. The variations even in this type of spicule are numerous as the number of the facets varies at each end independently. Thus, there may be one facet at one end, two at the other or two at each end, three at one end and four at the other and so on; but generally speaking if the number of these facets is not the same at each end the excess et one end is not greater than one over that at the other end. This may be represented as follows in figures:
1–1, 1–2, 2–2, 2–3, 3–3, 3–4, 4–4, 4–5.
In addition to the spicules of this type there are some quadruplets which are either simple crosses with rounded ends or crosses with two or three convexities at the end of each of the branches or, in a few cases, simple square plates with very rounded angles. The most noteworthy thing about these rachis spicules, however, is the absence of plain rods of full size or of oval spicules. Apart from a few small spicules, which are probably growth stages, all the spicules are swollen at the extremities and most of them show divided lines of growth.
In the outer layer of the stalk there is a dense armature of spicules of the same type as those prevalent in the rachis; in the inner structures of the stalk there are apparently very few spicules, but several of these which are found lying vertically to the surface in the fleshy septa are longer than those in the rachis, being 0·2 mm. in length.
Cavernularia malabarica Fowler.
Two specimens of this species were obtained by Dr Imms from Puri, Orissa Coast, Bay of Bengal. Their principal measurements are:
| Length of rachis 27·25 mm. | Diameter of rachis 35·22 mm. |
| " stalk 13·10 mm. | " stalk 10·8 mm. |
The sharp distinction in diameter between rachis and stalk which is a character of this species and of Cavernularia glans was well marked in the specimens. The stalk seems to be a little longer in proportion to the rachis than in the type specimens, but it is nevertheless a short stalk.
The species is of some interest as it afforded the first known examples of pennatulids to be "washed ashore."