THE
PALÆONTOGRAPHICAL SOCIETY.
INSTITUTED MDCCCXLVII.
LONDON:
MDCCCLIV.
A MONOGRAPH
ON THE
FOSSIL BALANIDÆ
AND
VERRUCIDÆ
OF GREAT BRITAIN.
BY
CHARLES DARWIN, F.R.S., F.G.S.
LONDON:
PRINTED FOR THE PALÆONTOGRAPHICAL SOCIETY.
1854.
C. AND J. ADLARD, PRINTERS, BARTHOLOMEW CLOSE.
PREFACE.
As the present short Monograph completes my work on British Fossil Cirripedes, I beg to be permitted again to have the satisfaction of returning my very sincere thanks to the many naturalists who have placed their collections at my disposal, and have given me the freest permission to use the specimens, in whatever manner I might find necessary.—My thanks are most especially due to Mr. Searles Wood, Mr. Bowerbank, and Sir Charles Lyell; and to Mr. J. de C. Sowerby for the use of the original specimens figured in the 'Mineral Conchology.' I lie, also, under much obligation to Mr. D. Sharpe, Mr. Greenough, Mr. Smith of Jordan Hill, Professor Tennant, Mr. Charlesworth, Mr. F. Edwards, Dr. T. Wright, Professor Forbes, Professor Henslow, M. Bosquet, and to many others. I must, also, be permitted to tender my grateful thanks to the Council of the Palæontographical Society for the very liberal manner in which they have allowed my two Monographs to be illustrated.
INTRODUCTION.
——————
CIRRIPEDIA may be divided, as I have recently shown in a monograph on the Balanidæ published by the Ray Society, into three Orders: of these, the Thoracica includes all ordinary Cirripedes, and all ever likely to be found fossil, and therefore the two other orders may be here passed over without notice. The Thoracica contains three Families: the Balanidæ or Sessile Cirripedes, which in a recent state so abound on the shores of almost every quarter of the world, and which are so frequently found in Tertiary deposits; the Verrucidæ, which includes only a single genus very singular from its asymmetrical shell; and the Lepadidæ, or Pedunculated Cirripedes; of the latter, the Fossil species have been already published by the Palæontographical Society. The Balanidæ and Verrucidæ will be treated of in the following pages. As yet only sixteen species in these two families have been found fossil in Great Britain; and of these sixteen, nine are still living forms. As the latter are known only imperfectly in their fossil condition, and as they have lately been described by me in full detail, I have thought it best here only to make a few remarks on such portions of the shells of each species as have hitherto been discovered, adding a few illustrations, such as appeared to me desirable. The extinct species will be fully described: of these, all the figures given are from British specimens. But of the species found both living and fossil, I have in several instances (always so stated) given drawings from recent specimens; some of the valves either not having been found fossil, or found only in an imperfect and not characteristic condition. As so few species in the several genera are known in a fossil condition, I have thought it quite superfluous to give long generic descriptions, which would have required constant references to many species exclusively found living.
In my former monograph on the Fossil Lepadidæ, I remarked how much the natural
history of Cirripedes has been neglected; and this remark is eminently applicable to the Balanidæ, or Sessile Cirripedes. Even the British recent species have not been well made out, and as for the fossil species, scarcely anything has been done, besides the publication of some figures, in very few instances accompanied by the details which are absolutely necessary for the identification of the species.
Owing to the great variation in external characters, to which almost all the species are subject, and likewise in the case of the principal genus, Balanus, to its being a very natural genus, that is, to the species following each other in close order, it is not easy to exaggerate the difficulty of identifying the species, except by a deliberate examination of the internal and external structure of each individual specimen. Every one who has collected Sessile Cirripedes must have perceived to what an extent their shape depends on their position and grouping. The surface of attachment has a great effect on that of the shell; for as the walls are added to at their bases, every portion has at one time been in close contact with the supporting surface; thus I have seen a strongly-ribbed species (B. porcatus) and a nearly smooth species (B. crenatus) closely resembling each other and both having a peculiar appearance, owing to their having been attached to a pecten. Dr. Gray has pointed out to me specimens of the recent B. patellaris, curiously pitted like the wood to which they had adhered; and numberless other instances might be added. Quite independently of the effect produced by the surface of attachment, the degree to which the longitudinal folds and ribs are developed on the parietes, is variable in most of the species, as in B. tintinnabulum and even in B. porcatus; the presence or entire absence of these ribs often surprisingly alters the whole aspect of the shell. The persistence of the so-called epidermis is in some degree variable, though this is of little importance in regard to fossil specimens. Again, some species in certain localities are all subject to the disintegration of the entire outer lamina of the walls; and in such cases (as with B. perforatus) there is not the smallest resemblance between the corroded and perfect specimens. The size of the orifice, and consequently of the operculum, compared with the shell, varies accordingly as the shell is more or less conical or cylindrical; in the latter case, the summits of the radii are generally more oblique and the orifice consequently more deeply toothed than in the more conical varieties. Size is a serviceable character in some cases, but very many specimens are required to ascertain the average or maximum size of each species, for there is no method of distinguishing a half-grown from a full-grown specimen; and I believe, as long as the individual lives, so long does it go on growing. Colour is of very considerable service; but the majority of the species have their white or nearly white varieties, the latter being sometimes as numerous as the coloured ones.
Besides the slight variation in the obliquity of the summits of the radii and alæ, dependent on the more or less cylindrical form of the shell, in some species, as in B. tintinnabulum and porcatus, their obliquity also varies occasionally from unknown causes,
and thus greatly affects the general appearance of the shell. In some few species, as in B. perforatus, the radii are often either not at all developed, or are of very variable width; in others, when the shell has become cylindrical, or when very old, the radii cease to grow, and from the disintegration of the whole upper part of the shell, with the continued growth of the lower part, the radii at last come to exist as mere fissures: I have seen instances of this in B. tintinnabulum and porcatus. Nevertheless, the obliquity of the upper margin, and the breadth of the radii are useful characters; and still more useful is the fact whether the upper margins are smooth and arched, or straight and jagged. The fact of the terga being more or less beaked is useful: as is, likewise, the presence of striæ, or furrows, or rows of pits, radiating from the apices of the scuta; but to ascertain the presence of these marks, it is almost invariably necessary to clean and examine the scuta with a lens; these ridges and furrows, moreover, in some species, as is strikingly the case with B. tintinnabulum, and in less degree with B. concavus, appear and disappear, and vary without any apparent cause.
Now if we reflect that form, size, state and nature of the surface, presence of epidermis, relative size of the orifice, presence of longitudinal ribs, tint, and often the existence of any colour, are all highly variable in most of the species; and that the obliquity of the summits of the radii, and the presence of longitudinal striæ on the scuta, are variable in some of the fossil species, we shall perceive how difficult it must ever be to distinguish the species from external characters. Let no one attempt to identify the species of this genus, without being prepared to separate, clean, and carefully examine with a microscope the basis and parietes, and both the under and upper surfaces of the opercular valves; for I feel convinced, that he will otherwise throw away much labour. Moreover, in many cases, it is almost necessary, on account of the variability of the characters, to possess several specimens. From these facts, I have not hesitated to use characters, for the identification of the species, which require close examination, though I would gladly have seized on external characters could I have found such even moderately constant.
The least varying, and therefore most important characters, must be taken from the internal structure of the parietes, radii, and basis: not that these characters are absolutely invariable; thus the porosity of the parietes is slightly variable in the recent B. glandula, and highly variable in the fossil B. unguiformis. The porosity of the basis is in some degree variable in B. spongicola. Characters derived from the general shape, and from the ridges and depressions on the under side of the scuta and terga, especially of the scuta, are highly serviceable; though even these are variable. The cause of the opercular valves offering more useful characters, as far as outline is concerned, than do the walls of the shell, is no doubt due to their being almost independent of any influence from the nature of the surface of attachment. Even the ridges and depressions on the under side of the scuta, which are in direct connexion with the muscles and soft parts of the animal, vary to a certain extent: thus the length and prominence of the adductor ridge is decidedly
variable in the fossil B. concavus and tintinnabulum; the size and form of the little cavity for the lateral depressor muscle varies in many species; so does the exact shape and degree of prominence of the articular ridge. There is one character in the terga, which at first would be thought very useful, namely, whether an open longitudinal furrow, or a closed fissure runs down outside the valve from the apex to the spur; but it is found that the furrow almost always gradually closes up during growth; and as a consequence of this, the width of the spur compared to that of the whole valve, as well as its distance from the basi-scutal angle, and the form of its basal extremity, all vary in some degree. The length of the spur sometimes varies considerably, as in B. concavus. The summits of the radii are apt to be oblique in the young of some species, whereas they are generally quite square in the old of the same. In some species the scuta become longitudinally striated only with age; on the other hand, in very young specimens of B. tintinnabulum, the scuta sometimes are deeply impressed by little pits placed in rows. I have already alluded to the longitudinal furrow on the tergum so entirely changing its character, owing to the edges becoming, during growth, folded inwards, and to the consequences which result from this. The inner lamina of the parietes generally loses, to a certain extent, its longitudinally ribbed character in old age. The basis is solid instead of being porose, in very young specimens of some species. In all the species, the carino-lateral compartments, in early age, are very narrow in proportion to the width of the lateral compartments; and in all, at this early period, the operculum is large in proportion to the whole shell.
Finally, I must state my deliberate conviction that Sessile Cirripedes can very seldom be satisfactorily identified in a fossil condition, without an examination of the opercular valves: hence when these have not been discovered, I have resolved, with some rare exceptions, not to attach a specific name to a shell without its operculum; for thus, I believe, I should add to the number of useless synonyms, which, as we shall immediately see, already exist. Nothing, indeed, could have been easier than to have affixed names to many groups of specimens, having different aspects, but to feel sure that these were really distinct species requires better evidence than can be afforded by the shell, without the opercular valves. When the specimens are much fossilised, it is, indeed, difficult to make out the primary points of structure in the genus Balanus—namely, whether the parietes, radii, and basis are porose: to do this it is sometimes necessary to rub down, polish, and carefully examine, a transverse section of a piece of the shell.
The ancient history of the Balanidæ is a brief one. No Secondary species has hitherto been discovered; in my former monograph on the fossil Lepadidæ1 I have shown that the negative evidence in this case is of considerable value, and consequently that there is much reason to doubt whether any member of the family did exist before the eocene period.
1 Since the note to page 5 of that work was written, I have been informed that the so-called cretaceous Tubicinella maxima is not a Cirripede.
The existence of a Cretaceous Verruca is an apparent exception to the rule, as this genus has hitherto always been ranked among Sessile Cirripedes; but Verruca, as we now know, must be placed in a family by itself, namely, the Verrucidæ, quite distinct from the Balanidæ. Balanus is the oldest genus as yet known; it first appeared in Europe and North America, during the deposition of the eocene beds; and was at that time, as far as our information at present serves, represented by very few species. In South America, one species of Balanus abounds in individuals in the ancient Patagonian tertiary formation. I have seen, in the British Museum, specimens said to have come from the eocene nummulitic beds, near the mouth of the Indus, belonging to that section of the genus, which has the walls and basis permeated by pores. Generally, the extinct forms belong to the section, which has the parietes not permeated by pores. During the miocene and pliocene ages, Sessile Cirripedes abounded. No extinct genus in this family has hitherto been discovered. It is singular, that though the Chthamalinæ approach much more closely than do the Balaninæ to the ancient Lepadidæ, of which so many species have been found fossil even in the older Secondary formations, yet that only one species of one genus of this sub-family has been hitherto found in any deposit; and that species is the still existing Pachylasma giganteum, in the modern beds of Sicily. During the epoch of the Glacial deposits in Scandinavia, Scotland, and Canada, the still existing species seem to have abounded; and they attained larger average dimensions than the same species now do on the shores of Great Britain, or even on the shores of the northern United States, where the average size seems larger than on this side of the Atlantic.
I have already given my reasons for very seldom naming any Sessile Cirripede without examining the opercular valves: it has been owing to this, as it appears to me, proper want of caution, that there are so many nominal species. Thus it is made to appear in catalogues, that the tertiary seas abounded with species of Balanus to an extent now quite unparalleled in any quarter of the world. Bronn,1 for instance, in his invaluable 'Index Palæontologicus,' gives the names of thirty-five Balani, found fossil in Europe, and I have not counted those found only in alluvial deposits, as they would certainly be the same as the still living species. Now I know only eleven recent Balani on the shores of all Europe, from the North Pole to lat. 30°; and of these I doubt whether B. balanoides and improvisus have been found fossil. In the Red Crag there is one extinct Balanus: in the Coralline Crag, which seems to have been very favorable to the existence of Cirripedes, there are six species of Balani, of which two are absolutely extinct, and one does not occur in any neighbouring sea: in the Eocene formations the species seem to have been rare, and I have seen only one, and that is an extinct form. Taking these several facts into consideration, and bearing in mind that Cirripedes usually range widely,
1 To save any other person interested in fossil Cirripedia, going through the several works quoted by Bronn, I have given some remarks on his list of species, in an appendix at the end of the Balanidæ, in my volume published by the Ray Society.
I do not believe, if all the specimens of Balani hitherto found in the several tertiary formations, from the Eocene to the Glacial deposits, throughout Europe, were collected together, they would amount to twenty species. I have myself seen, in a recognisable state, only twelve fossil species, of which five are extinct or not found in any neighbouring sea: I think it probable that three other recent species, viz., B. tulipiformis, perforatus, and amphitrite, may occur in the Mediterranean formations: and this would make fifteen species. Therefore, in the above estimate of twenty species, five are allowed for species existing in European collections, but not hitherto seen by me; and this, I believe, is a very full allowance. Consequently, even on the supposition that the five species just admitted as possibly existing in cabinets, and that the other five extinct species, which I have seen and examined, have all been previously named by other authors, a supposition excessively improbable, even then there would be fifteen superfluous names in Bronn.
The following short table shows how the Balanidæ and Verrucidæ were represented in Great Britain, throughout the several TERTIARY STAGES. It includes all the sixteen species described in the following pages, with the exception of one, the cretaceous Verruca prisca, which is the only member of either family hitherto found in any Secondary deposit.
| Living species, but found fossil in some tertiary deposits. | Mammaliferous crag, and glacial deposits. | Red crag. | Coralline crag. | Eocene. | ||
| Name. | ||||||
| Balanus | tintinnabulum . . | * |
.. |
* |
.. |
.. |
| calceolus . . | * |
.. |
.. |
* |
.. |
|
| spongicola . . | * |
.. |
.. |
* |
.. |
|
| concavus . . | * |
.. |
* |
* |
.. |
|
| porcatus . . . |
* |
* |
* |
.. |
.. |
|
| crenatus . . . |
* |
* |
* |
* |
.. |
|
| Hameri . . . |
* |
* |
* |
.. |
.. |
|
| bisulcatus . . | .. |
.. |
.. |
* |
.. |
|
| dolosus . . . |
.. |
* |
* |
.. |
.. |
|
| inclusus . . . |
.. |
.. |
.. |
* |
.. |
|
| unguiformis . . | .. |
.. |
.. |
.. |
* |
|
| Acasta
undulata . . . |
.. |
.. |
.. |
* |
.. |
|
| Pyrgoma anglicum . . | * |
.. |
.. |
* |
.. |
|
| Coronula
barbara . . . |
.. |
.. |
* |
.. |
.. |
|
| Verruca
Strömia . . . |
* |
* |
* |
* |
.. |
|
| Total 15, recent and extinct, found fossil in Great Britain, in some tertiary deposit. | 9 |
5 |
8 |
9 |
1 |
|
As affording some standard of comparison by which to compare the number of species found fossil in any Tertiary deposit, in relation to the number of species probably existing in the neighbouring seas during the same epoch, I may state that there are now living and propagating on the shores of Great Britain, eleven species belonging to the two Families included in the above table. In the Coralline crag, which seems to have been eminently favourable for the existence and subsequent preservation of Cirripedes, and which has been so well worked, nine fossil species of these two families, as may be seen in the table, have been discovered.
NOMENCLATURE OF THE SHELL OF A SESSILE CIRRIPEDE
——————
ARCHETYPE SHELL. Fig. 1.
Orifice of shell, surrounded by the sheath. Sheath formed by the alæ (a-a) and by portions of the upper and inner surfaces of the parietes (p-p).

N.B. In Balanus, and all known fossil genera, the Rostrum and Rostro-lateral compartments are confluent, and hence the Rostrum has the structure of Fig. 2.
COMPARTMENTS.
Fig. 2. Fig. 3. Fig. 4.
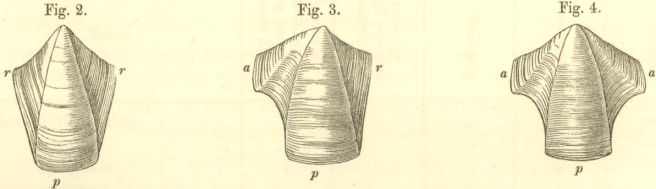
Fig. 2. Compartment with two radii, serving,
in fossil specimens, always as a Rostrum.
Fig. 3 serves as a Lateral and Carino-lateral Compartment. Fig. 4. serves as a Carina.
OPERCULAR VALVES.
Fig. 5. SCUTUM (internal view).

Fig. 6. TERGUM (internal view).
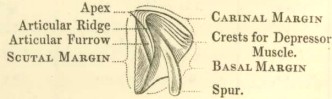
Fig. 7. TERGUM (external view).
On the Names given to the different parts of Cirripedes.
In my former volume I stated that I found it indispensable, in part owing to the extreme confusion of the nomenclature previously used, to attach new names to several of the external parts of Cirripedes. Almost all these names are applicable to the Balanidæ, or Sessile Cirripedes, and to the Verrucidæ; but a few additional names are requisite, which, together with the old names, will, I hope, be rendered clear by the accompanying woodcuts. In Sessile Cirripedes, the whole of that which is externally visible, may, for convenience sake, be divided into the operculum or opercular valves (valvæ operculares), and the shell (testa), though these parts homologically present no real difference. The operculum is seated generally some little way down within the orifice of the shell; but in very young specimens, and in Verruca, the operculum is attached to the summit of the shell, and in these cases the shell, without the operculum be removed, can hardly be said to have any orifice; though, of course, the opercular valves themselves have an aperture for the protrusion of the cirri.
The shell consists of the basis (called the support by some authors), and of the compartments (testæ valvæ), which in recent specimens vary from eight to four in number, and occasionally are all calcified together.
The compartment, at that end of the shell (fig. 1) where the cirri are exserted through the aperture or lips of the operculum, is called the carina (fig. 4); the compartment opposite to it, is the rostrum (in all fossil specimens, like fig. 2),—these two lying at the ends of the longitudinal axis of the shell. Those on the sides are the lateral compartments; that nearest the carina, being the carino-lateral (fig. 3) (testæ valva carino-lateralis), that nearest the rostrum, the rostro-lateral, and middle one simply the lateral compartment (fig. 3): but these three compartments are rarely present together. The rostro-lateral compartment, which always resembles fig. 2, and may be always known by having radii on both sides, is not known to occur in any fossil species; and hence we are here only concerned with the lateral and carino-lateral compartments. The compartments are separated from each other by sutures, which are often so fine and close as to be distinguished with difficulty. The edge of a compartment, which can only be seen when disarticulated from its neighbour, I have called the sutural edge (acies suturalis).
Each separate compartment consists of a wall (paries), or parietal portion (pp in figs. 1 and 4), which always grows downwards, and forms the basal margin; and is furnished on the two sides either with alæ (fig. 4), or with radii (fig. 2), or with an ala on one side and a radius (fig. 3) on the other.
The radius (adopting the name used by Bruguière, Lamarck, and others) differs remarkably in appearance (though not in essence) from the wall or parietal portion, owing to the direction of the lines of growth and the state of its usually depressed surface. In the upper part, the radii overlie the alæ of the adjoining compartments: in outline
2
(r, fig. 2, 3), they are wedge-formed, with their points downwards; their summits (and this is often a useful specific character) are either parallel to the basis, or as in fig. 1, 2, oblique. The radii are sometimes not developed.1
The alæ (so called by Dr. Gray) are overlapped by the radii and by part of the walls; they usually extend only about half way down the compartment (a, fig. 3, and 4); their summits are either parallel to the basis or oblique. The alæ of the several compartments, together with the internal, upper, thickened surfaces of the walls, against a shoulder of which the sutural edges of the alæ abut, have been called (by Dr. Gray) the sheath (vagina). The upper and greater portion of the sheath is marked by transverse lines, caused by the exuviation of the opercular membrane, as that membrane may be called, which unites the operculum all round to the sheath, or upper internal surface of the shell.
The carina has always two alæ, as in fig. 4.
The carino-lateral and lateral compartments have always an ala on one (the rostral) side, and a radius on the other (the carinal) side, as in fig. 3.
The rostro-lateral compartment (not at present known to occur in any fossil) has always radii on both sides, as in fig. 2.
The rostrum has normally alæ on both sides, as in fig. 4, but in many recent, and all the fossil species yet known, it has radii on both sides, as in fig. 2, owing to its fusion with the rostro-lateral compartments on both sides.
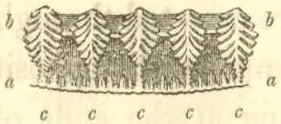
Basal edge of wall of compartment in Balanus tintinnabulum. a, a, outer lamina; b, b, inner lamina; c, c, longitudinal septa, uniting the inner and outer laminae, with their ends denticulated.
The walls of the shell, the basis, and the radii, are in very many cases composed of an outer and inner lamina, united together by septa; a set of pores or tubes being thus formed. The points of the septa generally project beyond the laminæ, and are denticulated on both sides as shown in the accompanying woodcut.
Operculum, or opercular valves.—These consist of a pair of scuta and a pair of terga. They are joined to the sheath of the shell by the opercular membrane.
Scutum (fig. 5): this valve is generally sub-triangular, and its three margins are the basal, the tergal, so called from being articulated with the tergum, and the occludent, so called from opening and shutting against the opposed valve. The angles are named, from the adjoining margins, as basi-tergal, &c.; the upper angle being the apex. The scutum is ordinarily articulated to the tergum by an articular ridge (crista articularis), running up to the apex of the valve, and by an articular furrow, which latter receives the
1 The radii have been called by Ranzani and De Blainville "areæ depressæ" (the parietal portions of the compartments being the "areæ prominentes"); by Poli, "areæ interjectæ;" by Gray, "sutures;" by Coldstream, "compartments of the second order," (the parietal portions being those of the first order); by some authors, "intersticia." I may here add that the scuta are the "ventral valves" of Gray, the "anterior" of Ranzani, and the "inferior opercular" of De Blainville: the terga are the "posterior valves" of Gray and Ranzani, but the "superior opercular" of De Blainville: the rostrum, on the other hand, is the "anterior valve" of Ferussac, and the "ventral" of De Blainville; the carina being the "dorsal valve" of the latter author.
scutal margin of the tergum. The articular ridge, instead of projecting straight up from the valve, when the latter is laid flat on its external surface, often bends over to the tergal side, and is then said to be reflexed. On the internal surface of the valve there is almost always an adductor pit or cavity (fossa adductoris), for the attachment of the adductor scutorum muscle: this pit is often bounded on its tergal and basal sides, by a ridge, called the adductor ridge (crista adductoris), which, in its upper part, is often confluent with the articular ridge. Beneath the adductor ridge, in the basi-tergal corner of the valve, there is often a lateral-depressor pit (fossa musculi lateralis depressoris), for the attachment of the so-called muscle; and this pit is sometimes furnished with crests.
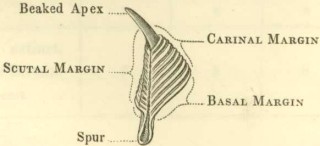
Tergum, (figs. 6 and 7):—this valve, also, has three margins, the scutal, basal, and carinal; its upper end, or apex, is sometimes beaked; on the basal margin a spur (calcar) depends; the outer surface of the valve is depressed or longitudinally furrowed (sulcus longitudinalis) in the line of the spur. The part called the spur is often so broad, that the name becomes not very appropriate. The angles are denominated, from the adjoining margins, as basi-carinal, or basi-scutal angle, &c. On the under side, in the upper part, there is an articular ridge, and on its scutal side, an articular furrow, receiving the articular ridge of the scutum. In the basi-carinal corner of the valve there are often crests for the attachment of the tergal depressor muscle.
Relative position of parts.—The centre of the generally flat basis, which is cemented to the supporting surface, is properly the anterior end, and the tips of the terga, often hidden within the shell, are properly the posterior end of the external covering; but I have found it more convenient to speak of the upper and basal surfaces and aspects, which hardly admit of any mistake. A line drawn from the centre of the basis, along the middle of the rostrum to the tips of the scula, shows the strictly medio-ventral surface of the shell; and another line drawn from the centre of the basis, along the carina, to the tips of the terga, shows the strictly medio-dorsal line; but from the crooked course of these lines, I have found it far more convenient to speak of the rostral and carinal end or aspect of the different parts of the shell. There has, moreover, been great confusion in these relative terms, as applied by different authors.
When a sessile Cirripede is held in the position in which they have generally been figured, namely with the basis downwards and the scuta towards the beholder, then the right and left sides of the Cirripede correspond with those of the holder.
SUB-CLASS—CIRRIPEDIA. ORDER—THORACICA.
Family—BALANIDÆ.
Cirripedia sine pedunculo; scuta et terga musculis depressoribus instructa; reliquæ testæ valvæ inter se immobiliter conjunctæ.
Cirripedia without a peduncle; scuta and terga furnished with depressor muscles; other valves united immoveably together.
This family, which includes all true Sessile Cirripedes, may be divided into two very natural sub-families; namely the Balaninæ and Chthamalinæ; but as not one member of the latter has been found fossil in Great Britain, and indeed only one, the Pachylasma giganteum, in any part of the world, viz., in the recent beds of Sicily, this sub-family of the Chthamalinæ may be here passed over in silence.
Sub-Family—BALANINÆ.
Rostrum cum radiis, sed sine alis: valvæ testæ laterales omnes, ex uno latere alis, ex altero radiis instructæ: parietes ferè aut porosi, aut ad interiorem superficiem longitudinaliter costati.
Shell with the rostrum having radii, but without alæ; lateral compartments all having alæ on one side and radii on the other side; parietes generally either porose, or longitudinally ribbed on their inner surfaces.
Genus—BALANUS, Auct.
CONOPEA (pars generis). Say. Journal Nat. Sc. Philadelphia, vol. ii, Part 2, 1822.
MESSULA (do.) Leach. Zoological Journal, vol. ii, 1825.
CHIRONA (do.) J. E. Gray. Philosoph. Transacts., 1835, p. 37.
Valvæ operculares inter se articulatæ, subtriangulares; valvæ testæ 6; basis calcarea aut membranacea.
Scutum and tergum articulated together, sub-triangular; compartments six; basis calcareous or membranous.
The genus Balanus already includes 45 species, recent and fossil, and consequently in my volume published by the Ray Society, I have divided the genus into sections, on characters derived from the porosity of the parietes, radii, and basis; and on whether the basis be membranous or calcareous. But as here we have to describe or notice only eleven species, I have thought it more convenient to drop the sections, and in their place add a few words to each of the diagnostic characters. The genus is quite distinct from all the other genera of Sessile Cirripedes, with the exception of the sub-genus Acasta, from which its separation, it must be confessed, is in one sense artificial; for the species of this sub-genus graduate into those of Balanus (such as B. calceolus and its allies), which have their shells elongated in the rostro-carinal axis, and which live attached to Gorgoniæ. These latter species have been generically separated by some authors from true Balanus; but I have found it impossible to effect this; and even the section of the genus, including these species, is hardly distinct enough from the adjoining sections. On the other hand, the sub-genus Acasta, in another sense, is a very natural one, inasmuch as all its species are closely allied together in essential structure, in general appearance, and in habit; and as the genus Balanus is already large, I have thought it best to adopt Acasta, which has been already admitted by many authors as a sub-genus. I need only further remark, that from reasons already assigned, I have thought it useless to give in this work long generic descriptions.
1. BALANUS TINTINNABULUM. Tab. I, fig. 1a-1d.
LEPAS TINTINNABULUM. Linn. Syst. Naturæ, 1767.
— — Ellis. Phil. Transact. vol. 50, 1758, Tab. 34, figs. 8 and 9.
— — Chemnitz. Neues Syst. Conch., 8 B. (1785), Tab. 97, figs. 828-831.
BALANUS TULIPA. Bruguière. Encyclop. Meth. 1789; sed non B. tulipa alba, in Chemnitz; nec non B. tulipa, O. F. Müller, Zoolog. Dan.; nec non B. tulipa, Poli, Test. ut Siciliæ.
— — G. B. Sowerby. Genera of Recent and Fossil Shells, Tab. Genus Balanus.
LEPAS CRISPATA (var.) Schröter. Einleitung Conch., vol. iii, Tab. 9, fig. 21.
— SPINOSA (var.) Gmelin. Linn. Syst. Nat.
— TINTINNABULUM, SPINOSA, CRISPATA ET PORCATA. W. Wood. General Conchology, 1815, Pl. 6, figs. 1, 2. Pl. 7, figs. 4, 5. Pl. 8, figs. 1-5.
BALANUS TINTINNABULUM. Chenu. Illust. Conch.
— D'ORBIGNII (var.) Chenu. Illust. Conch., Tab. 6, fig. 10, sed non Tab. 4, fig. 13.
—CRASSUS. Sowerby (!) Min. Conch., 1818, Tab. 84.
B. parietibus et basi et radiis poris perforatis: testâ a roseâ ad atro-purpuream variante, sæpe longitudinaliter virgatâ et costata: orificio plerumque integro, interdum dentato. Scuti cristâ articulari latâ et reflexâ. Tergi margine basali plerumque in contrariis calcaris partibus rectam lineam formante.
Walls, basis, and radii permeated by pores; shell varying from pink to blackish purple, often striped and ribbed longitudinally; orifice generally entire, sometimes toothed. Scutum with the articular ridge broad and reflexed. Tergum with the basal margin generally forming a straight line on opposite sides of the spur.
Fossil in the Red Crag (Sutton). Mus. S. Wood, J. de C. Sowerby. Touraine (?) Mus. Lyell.
Recent, on West Coast of Africa; Madeira; West Indies; Cape of Good Hope; Mouth of the Indus; East Indian Archipelago; Sydney, Australia; Peru; Galapagos Islands; West Mexico; California.
Of this species I have seen several specimens, and fragments. Three of these are the original specimens figured in the Mineral Conchology, as B. crassus, an examination of which I owe to the great kindness of Mr. J. de C. Sowerby. Some specimens equally or more perfect are in Mr. S. Wood's collection. I have further seen a specimen from Touraine, which was presented to Sir C. Lyell by M. Dujardin, under the name of B. fasciatus, which I fully believe to be B. tintinnabulum. None of these specimens had opercular valves, and therefore it is perhaps rash to assert quite positively that they are identical with B. tintinnabulum; but, extraordinarily variable as this latter species is, yet, after having examined so many hundreds of recent specimens from all quarters of the globe, a sort of instinctive knowledge of general aspect is acquired, which makes me feel convinced that the fossils in question do really belong to this species. Moreover, the large shell, with its trigonal orifice passing into rhomboidal,—the smooth, broad, finely porose radii, with their summits not oblique,—the rather large parietal pores,—and the cancellated basis, are characters which hardly concur in any other species; and those with which these fossils might be confounded, are inhabitants of distant quarters of the world. Most of the recent varieties of B. tintinnabulum, and all the fossil specimens from the Crag, can be at once discriminated from B. tulipiformis (with which, at least in the Mediterranean deposits, it is likely to be confounded) by the summits of the radii extending from tip to tip of the adjoining compartments, and therefore not being oblique, as is always the case with the radii of B. tulipiformis. The largest fossil specimen which I have seen is nearly two inches in basal diameter, and nearly the same in height, and therefore about two thirds of the size of the largest living specimens.
I have had engraved, from recent specimens, an internal view of the scutum and tergum, as these are likely hereafter to be found by searchers in the Crag deposits; and I may refer to my Monograph on the Balanidæ for their full description. It may be observed in the habitats given of the living specimens, that Madeira is the nearest point where the species now lives and propagates; but specimens in full vigour are often brought to the British shores, attached to the bottoms of vessels.
2. BALANUS CALCEOLUS,* Tab. I, fig. 2a-2d.
BALANUS CALCEOLUS KERATOPHYTO INVOLUTUS (?) Ellis. Phil. Trans., vol. 50 (1758), Tab. 34, fig. 19.
LEPAS CALCEOLUS (?) Pallas. Elench. Zooph., p. 198, (sine descript.) (1766).
CONOPEA OVATA (?) J. E. Gray. Annals of Philosophy, vol. 10, 1825.
B. testæ axe rostro-carinali elongato; basi cymbiformi; parietibus et basi, sed non radiis, poris perforatis. Scuto musculi depressoris lateralis fossâ parvâ, profundâ.
Shell with its rostro-carinal axis much elongated; basis boat-shaped; walls and basis porose, but not the radii. Scutum with the pit for the lateral depressor muscle small and deep.
Fossil in Coralline Crag, attached to a Gorgonia; Sutton; Mus. S. Wood.
Recent, attached to Gorgoniæ, West Coast of Africa. Tubicoreen, near Madras. Mediterranean (?).
I have seen only a single fossil specimen of this species, nearly half an inch in length. The shell was perfect, and a small portion of the Gorgonia yet remained attached to the grooved and boat-shaped basis. The opercular valves had been lost, but the shell in this instance is so peculiar, that it could only be confounded with the recent B. galeatus, cymbiformis, or navicula, and from all these it is easily distinguished by the parietes being permeated by pores. It is, of course, possible, that the opercular valves might present some new character, showing that this fossil, though agreeing with Bal. calceolus in its shell, yet was specifically distinct. I have given a drawing of the opercular valves from recent specimens, which have been fully described in my Monograph on the Balanidæ. In regard to the shell, the fossil specimen could not be distinguished from the recent; and as it had to be broken, in order that its internal structure might be examined, I have thought it best to give a drawing from a perfect recent specimen. The spur of the tergum, in recent specimens, sometimes presents a singular character, in being irregularly toothed, and I have given a drawing (fig. 2d) of this variety, as it might perplex a collector.
1 With respect to the nomenclature of this and three allied recent species, I must remark that in the published descriptions no allusion is made to any one of the characters by which alone they can be distinguished: hence I have been guided by geographical probabilities in assigning the specific name of calceolus to the present species, as Ellis's specimens came from the Mediterranean; and that of galeatus to the North American and West Indian specimens, as Linnæus' original specimens (according to a statement by Spengler) came from the West Indies. I have assigned new names to the two remaining East Indian species.
3. BALANUS SPONGICOLA, Tab. I, fig. 3a-3e.
BALANUS SPONGICOLA. Browne's Illustrations of the Conchology of Great Britain (1827), pl. 7, fig. 6: 2d edit. (1844), pl. 53, figs. 14-16.
B. parietibus et basi, sed non radiis poris perforatis; parietibus plerumque lævibus, roseis; orificio dentato; scuto longitudinaliter striato; tergum, apice producto, sine sulco longitudinali, calcare truncato, 1/3 valvæ latitudine.
Parietes and basis, but not the radii, permeated by pores; parietes generally smooth; shell pink; orifice toothed; scutum longitudinally striated; tergum, with the apex produced, without a longitudinal furrow; spur truncated, about one third of width of valve.
Fossil in Coralline Crag; Sutton; Mus. S. Wood.
Recent on the South coast of England, and Tenby in South Wales; Algiers; Madeira; Lagulhas Bank, Cape of Good Hope.
I have seen only a single specimen of this species, which I picked out of a mass of specimens of the extinct Bal. inclusus, collected by Mr. Wood, in the Coralline Crag at Sutton. This one specimen was perfect, and included the opercular valves; it even partially retained its rosy colour: it was .3 of an inch in basal diameter, and therefore exactly half the size of the largest recent specimen which I have seen. It was in every respect perfectly characterised. I have given drawings, external and internal, of the scutum and tergum from the fossil specimens. In the scutum, the adductor ridge is, perhaps, rather more prominent, and the pit for the lateral depressor muscle rather deeper than in recent specimens; but these points are extremely variable. The tergum, in its outline, strictly agrees with the European recent specimens, and not with those varieties from the Cape of Good Hope and West Indies; indeed, in the degree in which the basal margin on the carinal side of the spur slopes towards the spur, it even, perhaps, exceeds the European variety. These valves are fully described in my Monograph on the Balanidæ. From the shell alone, as viewed externally, Bal. spongicola, even in its recent state, can hardly be distinguished from Bal. tulipiformis, or from some varieties of Bal. Capensis: I doubt whether this species could anyhow be distinguished in its fossil condition from the young of the fossil Bal. concavus, without the aid of the opercular valves. But in order to give an idea of its general appearance, and as I was compelled to disarticulate the compartments of the one fossil shell, I have had a fine recent specimen from the Mediterranean engraved on an enlarged scale.
4. BALANUS CONCAVUS, Tab. I, fig. 4a-4p.
BALANUS CONCAVUS. Bronn. Italiens Tertiar-Gebilde (1831) et Lethæa Geognostica, b. ii, s. 1155 (1838), Tab. 36, fig. 12.1
— CYLINDRACEUS, var. c. Lamarck. Animaux sans Vertèbres (1818).
LEPAS TINTINNABULUM. Brocchi. Conchologia Sub-Appen., t. ii, p. 597, (1814).
B. parietibus et basi, sed non radiis poris perforatis; testâ albo cum roseo aut obscurè purpureo longitudinaliter pictâ, interdum purè albâ. Scuto longitudinaliter tenuiter striato: internè, adductoris cristâ admodum aut modicè prominente.
Parietes and basis, but not the radii, permeated by pores; shell longitudinally striped with white and pink, or dull purple; sometimes wholly white; scutum finally striated longitudinally; internally, adductor ridge very or moderately prominent.
Fossil in Coralline Crag, (Ramsholt and Sudborne) rarely in the Red Crag (Sutton); Mus. S. Wood, Bowerbank, Lyell, J. de C. Sowerby, Tennant. Sub-Appenine formations, near Turin, Asti, and Colle in Tuscany, Mus. Greenough, &c. Tertiary bed, near Lisbon, Mus. D. Sharpe and Smith. Bordeaux (?) Mus. Lyell. Tertiary beds, Williamsburg; and Evergreen, Virginia, Mus. Lyell. Maryland, Mus. Krantz. Pleiocene formations2 near Callao, Peru, Mus. Darwin.
Recent at Panama; Peru; S. Pedro, California; Philippine Arch.; Australia.
This species has caused me much trouble. It will be convenient first to make a few remarks on the recent specimens; I examined several from Panama and California, which, though differing greatly in colour, resembled each other in their scuta having the adductor ridge extremely prominent, and in having (Tab. I, fig. 4n) an almost tubular cavity for the attachment of the lateral depressor muscle,—characters which at first appeared of high specific value; but I soon found other specimens from Panama in which these peculiarities were barely developed. I then examined a single specimen from the Philippine Archipelago, resembling in external appearance one of the Panama varieties, but differing in the scuta being externally strongly denticulated in lines instead of being merely striated,—in the adductor ridge being far less prominent,—and in the spur of the tergum being broader and more truncated; I therefore considered this as a distinct species. I then examined a single white rugged specimen from the coast of Peru, which differed from the Philippine specimen in the shape of the well-defined denticulations on the scuta, and in some other trifling respects, and in the segments of the posterior cirri bearing a greater number of spines; with considerable doubt, I also named this as distinct. But when I came to
1 I suspect that B. pustularis, miser, and zonarius, all figured by Münster in his 'Beiträge,' b. iii, Tab. 6, may be this species.
2 I procured this specimen from the Island of S. Lorenzo, off Callao; it was imbedded, together with seventeen species of recent shells and with human remains, at the height of eighty-five feet.
3
examine a large series of fossil specimens from the Coralline Crag of England, and others from northern Italy, from Portugal, and from the southern United States, I at once discovered that the form of the denticuli on the striæ of the scuta was quite a worthless character,—that in young specimens the scuta were simply striated,—that the prominence of the adductor scutorum ridge and the depth of the cavity for the lateral depressor muscle varied much (as in the case of the recent specimens), owing apparently to the varying thickness of the valve,—that in the terga the spur varied considerably in length and breadth, the latter character being in part determined by the varying extent to which the edges of the longitudinal furrow are folded in,—and lastly, that in young specimens the basal end of the spur is much more abruptly truncated than in the old. Hence I was led to throw the three recent forms, originally considered by me as specifically distinct, into one species; but I may repeat that this considerable variation in the prominence of the adductor ridge, and in the depth of the pit for the lateral depressor muscle—the pit in some cases becoming even tubular—is a very unusual circumstance.
With respect to the fossil specimens from the above stated distant localities, I consider them as belonging to one species, though they vary considerably in several points of structure. When compared with the recent specimens, they differ from them in often attaining a considerably larger size; in the parieties being generally longitudinally ribbed, as in the case of the Coralline Crag specimen (Tab. I, fig. 4a), and in the radii often having more oblique summits. Some of the specimens from the United States, have strong rugged, depressed shells, frequently resembling, to a curious degree, Bal. porcatus. On the other hand, considering the many points of identity between the fossil and the recent specimens, I have concluded, without much doubt, that they ought all to be classed together. In the Coralline Crag specimens, the spur of the tergum (Tab. I, fig. 4g) is unusually long and narrow; it is broader in the Italian specimens (4o), and either short (4k) or long in the United States specimens. The scuta of the Lisbon specimens are remarkable for the great prominence of the adductor ridge, and for the depth of the lateral depressor cavity, as in most, but not in all, of the Panama specimens. The opercular valves, however, of some of the specimens from all these several distant localities are identical with the recent ones from the coast of America. I have entered into the above particulars, on account of, in the first place, its offering an excellent example how hopeless it is in most cases to make out the species of this difficult genus without a large series of specimens; secondly, as showing how the characters alter with age; and thirdly, as a good instance of the amount of variation which seems especially to occur in most of the species which have very extensive ranges.
Some of the pink-striped Panama varieties, though having a somewhat different aspect, can be distinguished from certain varieties of B. amphitrite only by their scuta being longitudinally striated,—a character in this species variable in degree, and in most cases of very little value. Some of the other recent varieties, however, are sufficiently distinct from B. amphitrite; and the great fossil Coralline Crag specimens, which stand at the opposite
end of the series of varieties, with their ribbed walls, very oblique radii, and coarsely striated scuta, are extremely unlike B. amphitrite.
With respect to the nomenclature of the present species, I have little doubt that I have properly identified the Italian fossil specimens with B. concavus of Bronn, who has given a very good figure of this species in his 'Lethæa Geognostica;' but it must be confessed that the longitudinal striæ on the scuta are not there represented. Considering the large size and frequency of this species in Europe and in the United States, it has probably received several other names besides the two synonyms, quoted at the head of this description. I should add that the true B. cylindraceus (not var. C) of Lamarck, according to the plate given by Chenu in his 'Illust. Conch.' is the B. psittacus of South America. I have seen in collections specimens of B. concavus labelled as B. tulipa of Poli (B. tulipiformis of my Monograph),—a very natural mistake, without the opercular valves be carefully examined.
General Appearance.—Shell conical (fig. 4a), often steeply conical (fig. 4c), but sometimes depressed and smooth (fig. 4d); orifice generally rather small, varying from rhomboidal to trigonal, with the radii narrow, and generally in the fossil specimens very oblique; surface generally smooth, sometimes rugged, and in the Coralline Crag specimens commonly ribbed longitudinally, the ribs being narrow. In the recent specimens the colour is various, either dull reddish-purple with narrow nearly white, or wider dark longitudinal bands; or, again, pale rosy-pink with broad white bands; or lastly, wholly white. The radii are either darker or paler than the parietes. The opercular valves are either dark purple or nearly white. Pale pink and white stripes are visible on some of the Italian and Portuguese tertiary specimens; and in most of the fossils the sheath is tinged dull red.
Dimensions.—The largest actually recent specimen which I have seen, from the Philippine Archipelago, had a basal diameter of 1.2 of an inch; the Peruvian pleistocene specimen is 1.7 in diameter; specimens from the crag and from the Italian deposits, however, sometimes slightly exceed two inches in basal diameter, and three in height.
Scuta: these in young and moderately-sized specimens are striated longitudinally (fig. 4l), sometimes faintly, but generally plainly, causing the lines of growth to be beaded; but in large and half-grown specimens, the lines of growth are often extremely prominent, and being intersected by the radiating striæ, are converted into little teeth or denticuli. As the striæ often run in pairs, the little teeth frequently stand in pairs, or broader teeth have a little notch on their summits, bearing a minute tuft of spines. In very old and large specimens, the prominent lines of growth are generally simply intersected by deep and narrow radiating striæ (Tab. I, fig. 4p). In one case, a single zone of growth in one valve was quite smooth, whilst the zones above and below were denticulated. The valve varies in thickness, which I think influences the prominence of the lines of growth and the depth of the striæ. These striæ often affect the internal surface (fig. 4h) of the basal margin, making it bluntly toothed. The articular ridge (fig. 4n), is rather small, and moderately reflexed. The adductor ridge (as already stated) varies remarkably; in most of the recent Panama specimens (fig. 4n),
and in the fossils from Portugal, it is extremely prominent, and extends down to near the basal margin; in other specimens it is but slightly prominent, as in those from the Crag (4f); it is short, but rather prominent in the specimens (4h) from Maryland; whereas it is very slightly prominent in the specimens from Virginia. The cavity for the lateral depressor, also, varies greatly; it is often, as in the recent specimens, bounded on the side towards the occludent margin by a very slight strait ridge, which occasionally folds a little over, making almost a tube; this, at first, I thought an excellent specific character, but far from this being the case, the cavity often becomes, in recent specimens as well as in the crag specimens (4f), wide, quite open, and shallow. The whole valve in the Crag specimens (fig. 4e) is apt to be more elongated than in the recent or Portuguese specimens (fig. 4l), and especially than in the Maryland (fig. 4h) specimens.
Terga very slightly beaked; the surface towards the carinal end of the valve, in some of the fossil specimens, is feebly striated longitudinally. There is either a slight depression (fig. 4k), or more commonly a deep longitudinal furrow (fig. 4g, 4o) with the edges folded in and touching each other, extending down the valve to the spur, and causing the latter to vary in width relatively to its length. When the furrow is closed in, the spur is about one fourth of the entire width of the valve, and has its lower end obliquely rounded, and stands at about its own width from the basi-scutal angle: when there is only a slight depression and no furrow (as is always the case with young specimens, and in the specimens (4k) from Maryland), the spur is broader, equalling one third of the width of the valve, with its lower end almost truncated, and standing at about half its own width from the basi-scutal angle. But the absolute length of the spur, also, varies considerably in the Coralline Crag specimens; it is often very long, (fig. 4g) compared to the whole valve. In many Italian specimens (4o) it is long and broad. The basal margin of the valve on the carinal side of the spur is sometimes slightly hollowed out; and when the longitudinal furrow is closed, this side slopes considerably towards the spur. Internally, the articular ridge and the crests for the tergal depressor muscles are moderately prominent.
Parietes, the longitudinal septa sometimes stand near each other, making the parietal pores small. The radii have oblique summits, but to a variable degree; their septa are unusually fine, and are denticulated on their lower sides; the interspaces are filled up solidly. The alæ have their summits very oblique, with their sutural edges nearly or quite smooth. In most of the fossil specimens (Tab. I, fig. 4b, r), and slightly in some of the recent specimens, the surface of the sheath presents an unusual character, in a narrow, longitudinal, slightly raised border, running along the sutures, on the rostral side of each suture.
Basis thin, porose; sometimes with an underlying cancellated layer.
All the recent specimens which I have seen, were, with one exception, attached to various shells and crabs, and to each other. The tertiary specimens are often congregated together into great masses. Including the recent and fossil specimens, this species encircles the globe. During the miocene period it seems to have been the commonest existing Sessile Cirripede; now, it does not appear to be common, excepting, perhaps, at Panama.
5. BALANUS PORCATUS. Tab. I, fig. 5a-5g.
BALANUS PORCATUS. Emanuel da Costa. Hist. Nat. Test. Brit., p. 249, (1778).
LEPAS BALANUS. Linn. Syst. Naturæ, (1767).
— — Born. Testacea Mus. Cæs. Desc., Tab. 1, fig. 4, (1780).
— — Chemnitz. Syst. Conch., 8 Band., Tab. 97, fig. 820, (1785).
BALANUS ARCTICA PATELLIFORMIS. Ellis. Philosoph. Transact., vol. 50, Tab. 34, fig. 18, (1758).
— SULCATUS. Bruguière. Encyclop. method., Tab. 164, fig. 1, (1789).
LEPAS COSTATA and BALANUS. Donovan. British Shells, 1802-1804, Tab. 30, fig. 1, 2.
LEPAS SCOTICA. W. Wood. General Conchology, Pl. 6, fig. 3, sed non Lepas balanus, Pl. 7, fig. 3, (1815).
BALANUS ANGULOSUS. Lamarck (1818), in Chenu, Illust. Conch., Tab. 11, fig. 11.
— TESSELATUS. Sowerby (!) Mineral Conchology, Tab. 84, (1818).
— SCOTICUS. Brown. Illust. Conch. Great Britain, Pl. 7, fig. 2, sed non Pl. 6, fig. 9 et 10 (1827): 2d edit., Pl. 53, fig. 1-3, 22, 23 et Pl. 54, fig. 1-3.
— GENICULATUS. Conrad. Journal Acad. Philadelphia, vol. vi, part 2, p. 265 (1830), Tab. 11, fig. 16.
— — Aug. Gould (!) Report on the Invertebrata of Massachussetts, fig. 9 (1841).
B. parietibus, sed non basi, poris perforatis; testâ albâ, plerumque longitudinaliter acutè costatâ; radiorum marginibus superioribus pæne basi parallelis: scuto longitudinaliter striato; tergi apice producto, purpureo.
Parietes, but not the basis, permeated by pores; shell white, generally sharply ribbed longitudinally; radii with their summits almost parallel to the basis. Scutum longitudinally striated; tergum with the apex produced and purple.
Fossil in the Glacial deposits of Scotland (Isle of Bute), of Uddevalla, and (Beaufort) Canada. In the Mammaliferous Crag (Bramerton, Thorpe) and Red Crag (Sutton); Mus. Lyell, J. de C. Sowerby, S. Wood, Bowerbank, &c.
Recent, England, Ireland, Scotland, Shetland Islands, Iceland, Davis's Straits, 66° 30' N.; Lancaster Sound, 74° 48' N. Maine and Massachussetts, United States. China (?) In deep water, commonly adherent on shells, crustacea, and rocks.
This species can be at once distinguished from all the foregoing by the basis being solid or not perforated by pores; and from all the following species, with the exception of B. crenatus, by the parieties having large square pores or tubes. From B. crenatus, this species can be distinguished by its longitudinally striated scuta, purple-beaked terga, and by the peculiar structure, immediately to be described, of its parietal pores; and in most cases even by its general aspect, larger size, and ribbed walls. When, however, B. porcatus and crenatus have grown together on the same irregular surface, for instance, on a Pecten, they sometimes resemble each other in a very deceptive manner. The opercular valves have not certainly been found fossil, but I have given drawings from recent specimens.
The parietes, (the basal margin of a small portion is represented at Tab. I, fig. 5b) are perforated by large square longitudinal tubes: in the upper part these are filled up solidly without transverse septa; the longitudinal septa between the tubes are finely denticulated at their bases, and the denticuli extend unusually close to the outer lamina. In very young specimens the inner lamina of the parietes is ribbed, in lines corresponding with the longitudinal septa, as in the case of other species of the genus; but in medium and large-sized specimens, there are between such ribs from one to four smaller ribs, which do not correspond with any longitudinal septa; these are finely denticulated at their bases, and may be considered as the representatives of longitudinal septa which have not been developed and reached the outer lamina. I have seen no other instance of this structure, namely, the presence of a greater number of ribs on the inner (upper in fig. 5b) lamina of the walls than there are longitudinal septa. As the parietes rest on the basis, the circumference of the latter becomes marked in a very peculiar manner (fig. 5c), by the basal edges of the parietal septa. Hence the basis of this species can be distinguished from that of every other sessile cirripede: its circumference is plainly impressed by the main parietal septa which connect the inner and outer laminæ of the walls; and between these marks there are two or three smaller impressions of the so-called representative septa, which do not extend beyond the impression of the basal edge of the inner lamina. The upper surface of the middle part of the basis (more especially when slightly disintegrated) is faintly striated in radiating lines, of which the stronger lines are prolonged from the circumferential marks left by the main parietal septa, and the weaker lines from the marks left by the representative septa.
Dimensions.—The largest recent specimens which I have seen from Great Britain or Ireland, have been 1.3 of an inch in basal diameter: in Mr. Cuming's collection, however, there is one much depressed specimen from the Shetland Islands, 2.1 in basal diameter; a regularly conical specimen from the coast of Massachussetts attains a nearly equal diameter. But out of the glacial deposits in the Isle of Bute, several specimens have this same diameter, namely, two inches, and are even more steeply conical, being 1.85 in height; some glacial specimens from Uddevalla and Canada, in Sir C. Lyell's collection, are 1.7 in basal diameter. Hence it appears, as we shall presently see is likewise the case with B. crenatus and Hameri, that northern specimens, and those from the United States and from the Glacial deposits, often exceed in dimensions those now living on the coasts of Great Britain and Ireland, or those found in the Crag.
This species is very common in the glacial deposits of Uddevalla, of Skien in Norway, and of Canada, and is associated with the same species, namely, B. crenatus and Hameri, as in the living state: I have seen, also, as just stated, specimens from the same formation in the Island of Bute, Scotland. I have examined numerous specimens from the Mammaliferous Crag, and a few from the Red Crag of England. I owe to the kindness of Mr. J. de C. Sowerby an inspection of the original specimens of B. tesselatus of the Mineral Conchology, which is certainly the present species.
6. BALANUS CRENATUS. Tab. I, fig. 6a-6g.
B. CRENATUS. Bruguière. Encyclop. Method. (des Vers) 1789.
LEPAS FOLIACEA, var. a. Spengler. Skrifter af Naturhist. Selskabet, b. i, 1790.
— BOREALIS. Donovan. British Shells, Pl. 163, (1802-1804).
B. RUGOSUS. Pulteney (?) Catalogue of Shells of Dorsetshire, 1799.
— Montagu (?) Test. Brit., 1803.
— Gould (!) Report on Invertebrata of Massachussetts (1841), fig. 10.
B. GLACIALIS (?) J. E. Gray. Suppl. Parry's Voyage, 1819.
B. ELONGATUS (!) CLAVATUS (!), auctorum variorum.
B. parietibus, sed non basi poris perforatis; testâ albâ; radiorum marginibus superioribus obliquis, asperis, rectis; scuto sine adductoris cristâ; tergi calcare rotundato.
Parietes but not basis permeated by pores; shell white; radii with their oblique summits rough and straight; scutum without an adductor ridge; tergum with the spur rounded.
Fossil in glacial deposits of Scandinavia and Canada, Mus. Lyell; in the mammaliferous and Red (Sutton) and Coralline Crags; Mus. S. Wood, J. de C. Sowerby, Bowerbank, &c. Miocene formation, Germany, Mus. Krantz.
Recent in Great Britain, Scandinavia, Arctic Regions as far as Lancaster Sound, in 74° 48' N.; Behring's Straits; United States; Mediterranean; West Indies; Cape of Good Hope. Generally attached to shells and crustacea in deep water.
Under the last species I have shown that the porose parieties, but solid basis, distinguish this species easily from all the others, with the exception of B. porcatus, from which it can readily be known by the characters of its opercular valves, as already thereunder stated. Judging by external appearances alone, which ought never to be trusted to in the identification of any sessile cirripede, this species might easily be confounded with Bal. dolosus, found fossil in the same deposits.
This species presents a great diversity of external aspect: I have had figured (Tab. I, fig. 6a) one of the commonest appearances presented by it; but frequently the shell is quite smooth and depressed, or extremely much elongated and cylindrical, or even club-shaped. The basis is generally thin and slightly furrowed in lines radiating from the centre, but it is not permeated by pores; when, however, in large and old specimens it becomes thicker, as in Tab. I, fig. 6c, its edge is very distinctly pitted by little hollows, which might sometimes be easily mistaken for the orifices of pores: the absence of pores is a very important character in the diagnosis of B. crenatus. The basis is less firmly attached to the supporting surface than is usual with most cirripedes, and consequently it often separates from it together with the parietes. With regard to the opercular valves (6d-6g) drawn from recent specimens, I need here only state that the most conspicuous
characters are the large articular ridge to the scutum, and the reflexed apices of all four valves, though this latter character is highly variable. I must refer to my Monograph on the Balanidæ for a full description of these valves.
The largest recent British specimen which I have seen was only .55 of an inch in basal diameter: specimens from Greenland and the northern United States, frequently attain a diameter of three-quarters of an inch, and I have seen one single somewhat distorted specimen actually 1.6 of an inch in basal diameter. Where individuals have grown crowded together, their length is often twice, and even occasionally thrice as great as their diameter; thus I have seen a recent Greenland specimen 1.6 of an inch in length, and only .75 in diameter. This species, in its recent state, as may be seen under the habitats, has an enormous range. I have felt myself unwillingly compelled to admit that it ranges from the Arctic Regions in 74° 48' N. to the Mediterranean, the West Indies, and Cape of Good Hope. That this species should live in the tropical seas is the more surprising, as the large size of the specimens in the northern seas and in the glacial deposits, might fairly have been supposed to have indicated special adaptation for a cold climate. This great geographical range, however, of the species accords with its range in time from the present day to the Coralline Crag period. The specimens from the glacial deposits which I have examined, chiefly in Sir C. Lyell's collection, are very fine and large, and appear, on an average, to attain as large or larger dimensions than the recent specimens from the United States; they are often associated, like the now living individuals, with B. porcatus and Hameri: they come from the well-known formation of Uddevalla, and from Canada. There are well-characterised specimens in the mammaliferous Crag, at Bramerton and near Norwich, in Sir C. Lyell's and Mr. Wood's collections, and from Sutton and other places in the Red Crag of the eastern shores of England: these specimens are not only smaller than the glacial, but than the recent English specimens; for the largest Crag specimens which I have seen had a basal diameter .5 of an inch, .3 to .4 being their ordinary size. The specimens which I have seen from the Coralline Crag, and some others sent me by Krantz from the miocene formation of Flonheim bei Abzei, in Germany, had not their opercular valves, yet I cannot doubt, considering how few species there are having porose walls and a solid basis, that I have rightly identified these specimens as belonging to B. crenatus.
7. BALANUS HAMERI, Tab. I, fig. 7a-7d., Tab. II, fig. 1a, 1b.
LEPAS HAMERI. Ascanius. Icones rerum naturalium, Tab. 10, 1767.
— TULIPA. O. F. Müller. Prodromus. Zoolog. Dan., 1776; sed non L. tulipa, in Poli, Test. ut Siciliæ; necnon B. tulipa, in Bruguière, Encyclop. method.; necnon B. tulipa, in Sowerby, Genera of Shells.
— TULIPA ALBA. Chemnitz. Syst. Conch., Tab. 98, fig. 832.
— FOLIACEA. Spengler. Skrivter af Naturhist. Selskabet, 1 B. 1790.
BALANUS CANDIDUS. (Tab. emendata) Brown. Conch. Great Britain (1827), Tab. 6, figs. 9 and 10, and 2d edit. Tab. 54, figs. 9-12.
— TULIPA. Lyell.1 In Phil. Transact., 1835, p. 37, Tab. 2, figs. 34-39.
B. nec parietibus, nec basi, nec radiis poris perforatis; testâ albâ; radiorum marginibus superioribus obliquis, lævibus, arcuatis; aciebus suturalibus lævibus; scuto angusto longitudinaliter, debiliter striato; tergi calcari angusto, rotundato.
Parieties, and basis, and radii not permeated by pores; shell white; radii with their oblique summits smooth and arched; sutural edges smooth; scutum narrow, feebly striated longitudinally; tergum with the spur narrow, rounded.
Fossil in Red Crag (Sutton), Mus. S. Wood. Doubtfully in the Glacial beds of Scotland. In the Glacial deposits at Uddevalla, in Sweden; and Beaufort, Canada, Mus. Lyell. Banks of the Dwina, Russia, Mus. Murchison. Greenland, "in blue clay," according to Spengler.
Recent on the Coast of Yorkshire; Scotland; Galway, Ireland; Isle of Man, and Anglesey, twelve fathoms. Generally in deep water; not very common. George's Bank, Massachussetts, United States. Iceland, Finmark, and the Faroe Island, according to Spengler. Attached to crustacea, mollusca, stems of fuci, and stones; often associated with B. porcatus and crenatus.
I have seen, in Mr. Wood's collection, from the Red Crag, fine and perfectly preserved specimens of a rostrum, and of a lateral compartment. The latter was three inches in height, and, including the alæ, one inch in width. I have also seen a specimen said doubtfully to have come from the glacial beds of Scotland. As it is so very common in the deposits of this same age in other countries, no doubt it will hereafter be found more plentifully in Scotland, and probably in the mammaliferous Crag of England. Balanus Hameri is a very fine species; I have seen a recent specimen from the coast of Yorkshire, two inches in diameter, and one inch and three-quarters in height: another specimen was three inches in height. The specimens in the glacial deposits, seem to have acquired larger dimensions: a compartment from Uddevalla being nearly four inches in height. The white colour, smooth surface, and regularly arched radii, give to the shell a very elegant appearance, which has appropriately been compared to that of a white tulip. The diagnosis of this species is easy; the walls as well as the basis being solid or not porose, serve to distinguish it from all other forms except certain varieties of Bal. unguiformis, and by several minor characters, such as the finely striated and more elongated scuta, &c., Bal. Hameri, can be discriminated from B. unguiformis.
1 Sir C. Lyell remarks that this is apparently the B. Uddevallensis, (Linn.), of Swedish lists of fossils. Prof. E. Forbes has shown ('Mem. Geolog. Survey of England,' vol. i, p. 364) how this name arose, from a short description, prior to the introduction of the binomial system, "Lepas quæ Balanus Uddevallensis," given by Linnæus in his Wast-Gotha Resa, in 1747.
For the reference to Ascanius' work, which is on the binomial system, and subsequent to the 10th edit. of Linnæus in 1758, I am greatly indebted to Mr. Sylvanus Hanley. Had it not been for this gentleman, I should have used Müller's name of B. tulipa as the first name.
4
In the recent condition, the compartments of the dead shell fall apart with singular facility; and Sir Charles Lyell has remarked ('Philosophical Transactions,' 1835, p. 37) that in the glacial deposits of Scandinavia, the shell is never found whole, but the separated compartments in abundance: it appears, also, that the basis likewise easily separates from its support. The extreme edge of the basis is finely crenated, and not pitted as in Bal. crenatus; the crenations or teeth are produced by the edge of the basis fitting in between the longitudinal septa on the internal surface of the parietes. There is one peculiarity in the alæ of this species in its recent state, which I have observed in no other species, and which can be distinguished in some of the fossil specimens, as in Tab. II, fig. 1b,—namely, the presence of an excessively fine linear furrow running along the sutural edge, a little towards the inner side, and filled (in the recent state), with a yellow ligamentous substance.
In regard to the opercular valves, (Tab. I, fig. 7a-7d, drawn from recent and glacial specimens), I need here only mention, that in the scuta, their flatness, elongation, and delicate longitudinal striæ, are their chief characteristics. In very old and large specimens of the terga (as in the specimen, fig. 7d, figured from Uddevalla), the basal margin on the carinal side of the spur slopes down towards it in a remarkable manner.
8. BALANUS BISULCATUS, Tab. II, fig. 2a-2h.
BALANUS SULCATINUS (?) Nyst, apud D'Omalius (sine descript. aut tabulâ), Géologie de Belgique, 1853.1
B. nec parietibus, nec radiis poris perforatis; basi poris magnis perforatâ; radiorum marginibus superioribus obliquis, lævibus; aciebus suturalibus lævibus; scuto angusto, sulcis longitudinalibus 2 ad 4; tergi calcare brevissimo dimidiâ valvæ latitudine.
Neither walls nor radii permeated by pores; basis permeated by large pores; radii with their upper margins oblique and smooth; sutural edges smooth; scutum narrow, with from two to four longitudinal furrows; tergum with the spur very short, broad as half the valve.
Var. plicatus (fig. 2c), with the walls deeply folded; radii narrow, with their upper margins very oblique.
Fossil in Coralline Crag; Ramsholt, Gedgrave, Sutton; Mus. S. Wood, Bowerbank, J. de C. Sowerby. Rauville, dans le Cotantin, Mus. G. B. Sowerby. Var. plicatus, Coralline Crag, Sutton, Mus. S. Wood, Bowerbank. Bolderberg, near Hasselt, Belgium, Mus. Bosquet.
———————————————————————————————————————————
1 I am indebted to M. Bosquet for a specimen, bearing this name and reference, found in the 'Système Bolderien' of Dumont, (miocene according to Sir C. Lyell) at Bolderberg. The specimen consists of a rostrum, with a portion of the base attached; and as these parts are in some degree characteristic, I fully believe this specimen to be B. bisulcatus.
General Appearance.—Shell (fig. 2a) conical or tubulo-conical, often rather globose; walls frequently thin, either very smooth, or deeply plicated longitudinally: occasionally the same specimen is smooth in the upper part ( fig. 2b), and strongly plicated in the lower. The radii in the large specimens are wide, and with their upper margins only slightly oblique; in the smaller they are narrower, and much more oblique; but in each case their upper margins are smooth and slightly bowed. Colour apparently originally nearly white, but with the alæ generally, in the smaller specimens, clouded with a dark tint: the radii are usually striped feebly in longitudinal lines. Basal diameter of largest specimen .8 of an inch; but this seems to have been an unusual size.
Scuta: (fig. 2e) narrow, with the basal margin forming an unusually small angle with the occludent margin; surface slightly convex, with lines of growth approximate, moderately prominent; on the tergal half of the valve, two distinct rather broad furrows, with sometimes a third, and even a fourth, nearer to the occludent margin, extend from the apex down the valve, and give it a very peculiar appearance: the furrows near the tergal margin are the deepest. Internally (fig. 2g), the upper part of the valve is roughened with small points: the articular furrow is unusually wide: the articular ridge is very prominent and but little reflexed, with the lower end almost abruptly cut off: the adductor ridge is prominent, but short: there are small deepish pits for the rostral and lateral depressores.
Terga (fig. 2f), broad, flat, with a slight narrow prominent rim along the scutal margin, which margin is slightly bowed. The basal margin on the carinal side of the spur slopes so gradually towards the spur, that the latter is barely distinct, and is very short, not depending nearly half its own width beneath the basi-scutal angle: the spur, also, is broad, namely, measured across the upper part, as broad as half the valve; its basal end is obliquely rounded off on the carinal side; it is placed close to basi-scutal angle. The carinal margin of the valve is just perceptibly bowed, and is formed by rectangularly upturned lines of growth. Internally (fig. 2h), the upper part of the valve is rough; the articular ridge is prominent; the crests for the tergal depressores muscles are moderately well-developed.
Parietes, not porose; internally, the ribs are smooth, with their basal edges very finely or barely denticulated. The radii (as already stated) are of variable breadth; they have their upper margins either very slightly or highly oblique, but always smooth and rounded: their sutural edges are quite smooth, or sometimes, with a strong lens, traces of transverse striæ, representing septa, can just be discovered. The alæ have their upper margins very oblique; their sutural edges are, in the large specimens, quite smooth; in the younger ones, plainly crenated; the recipient furrow being clearly marked by the teeth. Basis plainly porose.
Varieties.—It is certain (fig. 2b) that there are longitudinally plicated specimens of this species, and that the obliquity of the upper margins of the radii also varies a little; nevertheless some of the deeply plicated specimens (fig. 2c) undoubtedly have a very
different aspect from the ordinary varieties, and do really differ in the sutural edges of the alæ being crenated, and in the greater narrowness and obliquity of the radii; but these points are all commonly variable. I have not seen any large specimens of the variety (fig. 2c), plicatus, so as to compare them with the large specimens of the normal form, yet I can hardly entertain any doubt, considering their agreement in so many important points, that I have rightly treated these forms as mere varieties; it is unfortunate that none of the specimens of the var. plicatus seen by me have had opercular valves, as their presence would have removed all shadow of doubt. I have given a drawing, enlarged seven times, of some very young shells (fig. 2d), adhering in numbers on Pecten Gerardii, which I believe belong to the plicated variety of our present species, but which are much too young to be identified with certainty.
Affinities: This is a strongly characterised species, and nearly allied only to the following species, B. dolosus. The furrows on the scuta in some degree resemble those on the recent B. lævis, but there is no alliance with that species. It is certain that amongst recent species, the chief affinity is with B. Hameri and amaryllis.
9. BALANUS DOLOSUS. Tab. II, fig. 3a-3d.
B. nec parietibus, nec radiis poris perforatis; basi poris magnis perforatâ, radiorum marginibus superioribus obliquis, lævibus; aciebus suturalibus item lævibus; tergi calcare non admodum brevi, 1/3 valvæ latitudine.
Neither walls nor radii permeated by pores; basis permeated by large pores; radii with their upper margins oblique and smooth; sutural edges smooth; tergum with the spur not very short, broad as one third of valve.
Fossil in Red (Sutton) and Mammaliferous Crag; Mus. S. Wood, Bowerbank, Lyell, J. de C. Sowerby, Henslow, &c. Mammaliferous Crag, Postwick, near Norwich, Mus. Lyell.
This species so closely resembles B. bisulcatus, both externally and in all the essential characters of the parietes, radii, and basis, that it is quite superfluous to describe again these parts. The specific characters are derived from the opercular valves, which present well defined distinctions, found by me constant in several specimens of both species. B. dolosus, like B. bisulcatus, has quite smooth and deeply plicated varieties, often adhering to the same univalve. The ribs on the inner surfaces of the parietes are remarkably prominent, as shown in the drawing (fig. 3a) of the inside of the rostrum. I think the upper margins of the radii are in this species rather more oblique than in B. bisulcatus. The sutural edges of the radii are marked by the finest striæ, representing septa. The sutural edges of the alæ are generally distinctly crenated. The basis is often slightly cup-formed, and very plainly porose (fig. 3b): its upper surface is marked by
radiating ridges: the septa between the radiating pores are themselves often in part porose, as was plainly the case in the specimen (fig. 3b) engraved. The orifice of the shell is large and elongated in its rostro-carinal axis, especially in young specimens. The basal diameter of the largest specimen is .4 of an inch.
The scuta (fig. 3c) have no trace of the two or three longitudinal furrows so conspicuous on these valves in B. bisulcatus, and which, in that species, run down to the basal margin from the apex of the valve, this fact showing that the furrows occur in quite young individuals. The whole valve is not quite so narrow as in B. bisulcatus, but otherwise agrees with it in shape: internally, there is hardly any difference; but the articular furrow is not quite so wide: the articular ridge is very prominent, and abruptly truncated at its lower end: the adductor ridge is also prominent; it here runs a little higher up the valve than in B. bisulcatus. The tergum (fig. 3d) differs more in the two species: the spur is not so broad; measured in its upper part, it is only about one third of the entire width of the valve, instead of being half as wide as the valve: it is considerably longer, depending beneath the basi-scutal angle more than half its own width; the basal margin of the valve on the carinal side, does not slope so gradually into the spur; the occludent and carinal margins are slightly arched, as in B. bisulcatus. Internally, the surface is rough, the articular ridge is prominent, and the crests for the tergal depressores are well developed,—all as in B. bisulcatus. It is remarkable how generally the opercular valves have been preserved in this species in its fossil condition, as compared with most other species of the genus.
It is not easy to distinguish, by external characters, the rugged varieties of this species from B. crenatus; indeed, the only difference is that the furrows receiving the edges of the radii, generally, exhibit in B. crenatus slight impressions of the septa, which are entirely absent in B. dolosus. By internal characters, such as the non-porose parietes, and porose basis, our present species widely differs from B. crenatus.
10. BALANUS UNGUIFORMIS. Tab. II, fig. 4a-4f.
BALANUS UNGUIFORMIS. J. de C. Sowerby (!) Mineral Conchology (sine descriptione), Tab. 648, fig. 1, (Jan. 1846.)
— ERISMA. J. de C. Sowerby (!) Ib., fig. 2.
— PERPLEXUS. Nyst, apud D'Omalius (sine descript. vel Tab.), Géologie de la Belgique, 1853.1
B. parietibus tenuibus, interdum poris perforatis: radiis sine poris, marginibus superioribus obliquis; aciebus suturalibus tenuissimè crenatis: basi sine poris: tergi calcare angusto, obtuso.
1 I am much indebted to M. Bosquet for specimens bearing this title, from Klein Spauwen, which certainly appear to me, as far as can be judged by the separated compartments, without the opercular valves, to belong to our present species.
Parietes thin, sometimes permeated by pores; radii without pores, with their upper margins oblique; sutural edges very finely crenated; basis without pores. Tergum with the spur narrow, bluntly pointed.
Var. erisma (fig. 4b), with the walls longitudinally folded or ribbed.
Fossil in the Eocene formations, Isle of Wight, Colwell Bay, Hordwell; Barton, (Chama Bed); Headon; Bembridge; Bergh, near Klein Spauwen, Belgium (?). Attached to various shells and wood. Mus. J. de C. Sowerby, E. Forbes, F. Edwards, Charlsworth, T. Wright, Bowerbank, Tennant, Bosquet.
This species, the most ancient one as yet well known in the genus, presents to the systematist a most unfortunate peculiarity, in the parietes being almost as often as not permeated by small pores: I have seen no other instance, except to a limited degree in the recent B. glandula, of this character being variable, and hence it must be still considered of high classificatory value, in so varying a genus as Balanus. Owing to the kindness of Mr. F. Edwards, I have seen the original specimens, excellently figured by Mr. J. de C. Sowerby, in the 'Mineral Conchology,' under the names of B. unguiformis and erisma, between which I can perceive no difference, excepting that the walls in the latter are longitudinally folded,—a character we know to be variable in many species. In both varieties, the parietes are sometimes porose and sometimes solid. The smaller specimens, however, figured in the 'Mineral Conchology' to the right hand of the Plate, may possibly be a distinct species, as I infer from the narrowness of their radii. This species is intimately allied to B. varians, a fossil from the ancient tertiary plains of Patagonia. It is also allied to the recent B. crenatus and glandula.
General appearance.—Shell (fig. 4a), tubulo-conical, sometimes even sub-cylindrical: surface either very smooth, or slightly folded, or deeply folded so as to be strongly ribbed longitudinally: orifice rather large, rhomboidal, narrow at the carinal end, toothed, but not deeply: walls rather thin and fragile: radii of moderate width, with their summits oblique, not quite smooth. Basal diameter of largest specimen about three quarters of an inch.
Scuta (fig. 4c), with the external surface smooth: there is a trace of a furrow running down the valve from the apex, near to the occludent margin, and this is only worth mentioning from the analogous furrows in B. bisculatus. Internally (fig. 4e), the upper surface of the valve is roughened: the articular ridge is very prominent, and slightly reflexed: there is no distinct adductor ridge; there is a slight but variable depression for the lateral depressor. Tergum ( fig. 4d), with the longitudinal furrow shallow; spur moderately long, about one fourth or one fifth of the width of the valve; placed at about its own width from the basi-scutal angle; basal end bluntly pointed; the basal margin on the opposite sides of the spur forms a nearly straight line; the carinal margin has an extremely narrow border formed by upturned lines of growth. Internally (fig. 4f), the surface is roughened with little points: the articular ridge is prominent: the crests for the tergal depressores moderately prominent.
Parietes: the longitudinal ribs on the internal surface are either feebly, or, in the lower part, strongly developed; their basal ends are only just perceptibly denticulated. As already stated, in about half the specimens, there were no traces of parietal pores; in the other half there were either distinct or obscure pores; the pores are circular, generally of unequal sizes, and never large; in the same individual they would sometimes be almost wholly absent in some of the compartments, and quite plain in the other compartments. The radii are either moderately wide or rather narrow, and have their upper margins very oblique, and not distinctly arched, and not quite smooth: their sutural edges are very finely crenated, the teeth or septa not being denticulated. The upper margins of the alæ are rather less oblique than those of the radii: their sutural edges are barely crenated. The basis is thin, and without any trace of pores; the upper surface is sometimes furrowed in radiating lines.
11. BALANUS INCLUSUS. Tab. II, fig. 5a-5g.
B. nec parietibus, nec radiis poris perforatis; basi poris perforatâ: testâ rufo-fuscâ: radiis latis, marginibus superioribus aut non obliquis aut modicè; aciebus suturalibus cum septis planè denticulatis: scuto sine adductoris cristâ: tergi calcare subangusto.
Neither walls nor radii permeated by pores; basis porose; shell reddish-brown; radii broad, with their upper margins not oblique, or only moderately oblique; sutural edges with plainly denticulated septa: scutum without an adductor ridge; tergum with the spur rather narrow.
Var. (a) (fig. 5c, 5d), with the shell elongated in its rostro-carinal axis; basis narrow, clasping the stem of a zoophyte; lateral compartments much broader than the almost linear rostrum, carina, and carino-lateral compartments.
Var. (b), with rough longitudinally folded walls, and with the summits of the radii forming an angle of about 45° with the basis.
Fossil in Coralline Crag; Sutton and Gedgrave; attached to foliaceous Bryozoa; Mus. S. Wood, Bowerbank. Var. a, Coralline Crag, Sutton, attached to cylindrical branches of corals; Mus. S. Wood, Bowerbank. Var. b, attached to shells, Osnabruck, Hanover, Mus. Lyell; Bunde, Westphalia, Mus. Krantz.
My materials consist of a beautiful series of specimens in Messrs. Wood and Bowerbank's collections; but unfortunately only a single young specimen had its opercular valves preserved. Not one specimen of the very curious variety (a) had opercular valves, yet I cannot feel any doubt about its being only a variety, caused by its attachment to a thin cylindrical branch of a coral, instead of to a foliaceous Bryozoon; it will, however, be convenient to give a separate description of this very remarkable form. With respect to var. (b), both sets of specimens came to me from the Continent, with the name of
B. stellaris, of Bronn; but as Bronn distinctly states, that in his species the parietes are porose, and as such is not here the case, this cannot possibly be that species: these specimens did not possess their opercular valves, and therefore cannot be identified with certainty.
General Appearance.—Shell conical (fig. 5a, 5b), with the orifice rather large, and rhomboidal. The surface is very smooth, except in var. (b) from the Continent, in which it is rugged and longitudinally folded. The colour is ochreous-brown (chiefly no doubt derived from the imbedding substance), tinged with red. The radii often have a much darker and more distinct red tint; they are sometimes longitudinally striped with dirty white. The radii are broad, with their summits straight, and very slightly oblique; in var. b, however, they slope at an angle of about 45°. Basal diameter of largest specimens .6 of an inch; but this is an unusual size.
Scuta, with the growth ridges little prominent. Internally (fig. 5f, from a young individual) the articular ridge is moderately prominent, with its lower end very obliquely rounded off; there is no adductor ridge; there is a minute pit for the lateral depressor muscle. Terga, with a slight longitudinal depression extending down to the spur; spur short, with its lower end almost square or truncated, about one fourth of width of valve, and placed at about half its own width from the basi-scutal angle. Internally (fig. 5g), the articular ridge is prominent; the crests for the tergal depressores are feebly developed.
Parietes, moderately thick and generally strongly ribbed internally, without parietal pores. Radii, wide, with their upper margins straight, not smooth or rounded, and very slightly (or, in var. b, moderately) oblique; their sutural edges have well-developed septa, which are denticulated: the interspaces between the septa are filled up solidly. The alæ have their upper margins oblique: they are only slightly, and sometimes not at all, added to above the level of the opercular membrane: their sutural edges are smooth. The basis is thin, but plainly porose.
Var. (a) (fig. 5c, 5d).—With respect to this remarkable variety, any one would at first think it specifically distinct. The shell is much compressed, or elongated in the rostro-carinal axis, sometimes to a great degree; I have seen a specimen .25 of an inch in this axis, and only .1 in its broadest part; but this is a very unusual degree of elongation. The most remarkable character is the extraordinary narrowness of the carina, of the carino-lateral compartments, and of the rostrum, compared with the great breadth, especially along the basal margin (fig. 5d), of the lateral compartments. The radii are of unusual breadth. The tips of the rostrum and of the lateral compartments are a little arched in, tending to make the shell somewhat globular. The true basis is extremely narrow (fig. 5d): it is deeply grooved, from clasping the thin, cylindrical stem of the coral to which it has adhered; and I have seen specimens in which the opposite edges of the groove had met, a tube having been thus actually formed. From the grooved basis, and from the elongation of the shell in the rostro-carinal axis, this variety presents so close a general resemblance to Balanus calceolus, and its allies, that I have seen it in a collection arranged on the same tablet with
a fossil specimen of B. calceolus. Notwithstanding the above several strongly-marked characters, by which this variety differs from the ordinary form, there is a resemblance in colour and aspect, which, though difficult to be described, made me from the first suspect that the two were specifically identical. In no point of real structure is there any difference, excepting that, perhaps, the pores in the basis are here rather smaller; but this might arise from the little development of the peculiar basis. Having come to this conclusion, I was interested by finding a specimen (fig. 5e) in Mr. Wood's collection, which had originally fixed itself (judging from the form of the basis) on a thick cylindrical stem, but which had subsequently grown on to an adjoining flat surface; consequently, one side of the shell presented all the peculiar characters of the present variety, but not strongly pronounced, whereas the other side, at the rostral end, was undistinguishable from the ordinary form. The unequal development of the rostrum on the two sides was very striking, and clearly showed how great an effect could be produced by the nature of the surface of attachment.
This singular variety cannot be considered accidental, in the sense in which this term may be applied in some cases: the pupa evidently fixes itself intentionally, in a certain definite position, on the branch of the coral (when a branch is chosen), exactly as in the case of Balanus calceolus, or Scalpellum vulgare,—species which always live attached to branches. But when other Balani occasionally fix themselves on branched corals, their position seems to be accidental and unsymmetrical; thus among the symmetrically elongated specimens of the present species, I found one specimen of Balanus bisulcatus, which had evidently been attached in an almost transverse position to a branch, and had thus become much distorted; so, again, I have seen specimens of the recent B. amaryllis attached irregularly to a Gorgonia, in the midst of the symmetrically elongated shells of Balanus navicula, an ally of B. calceolus.
This variety does not seem to attain so large a size as the ordinary form.
Affinities.—This species is allied to B. unguiformis and B. varians, but is perhaps more nearly related to the recent B. allium, an inhabitant of the Barrier Reef of Australia. The longitudinally folded variety (b) can hardly be distinguished by external aspect, or even by the opercular valves, from B. crenatus; but when the shell is disarticulated, the porose walls and non-porose basis of B. crenatus, allow of no mistake in the diagnosis of the two species.
Sub-Genus—ACASTA.
ACASTA. Leach. Journal de Physique, tom. lxxxv, 1817.
Valvæ testæ 6; parietes et basis non porosa; basis calcarea, cyathiformis, non elongata. Valvæ operculares inter se articulatæ, subtriangulares. Spongiis, aut rarò Isidis cortici, affixa.
Compartments six; parietes and basis non-porose: basis calcareous, cup-formed, not
5
elongated, attached to Sponges, or rarely to the bark of Isis: scutum and tergum articulated together, subtriangular.
Under the last genus, I have made a few remarks on the close affinities of this sub-genus to Balanus, and have given my reasons for retaining it, so that I need not here repeat them.
ACASTA UNDULATA. Tab. II, fig. 6a-6f.
A. testâ, ad speciem, ut in "A. spongites," sed majore: scuto externe striis longitudinalibus, sæpe binis, signato, sulcis intermediis latioribus: terge calcare, pæne 1/2 valvæ latitudine.
Shell, apparently, as in A. spongites, but larger: scutum marked by longitudinal ridges, often in pairs, with the intermediate furrows rather wide: spur of tergum nearly half as wide as valve.
Fossil in Coralline Crag (Sutton), Mus. S. Wood, Bowerbank.
I owe to Mr. Wood the inspection of a fine suite of valves, which, though separate, I have no reason to doubt have all been rightly attributed to the same species. Owing to the shell never having been found entire, its general shape is not known, and, what is of more consequence, the relative proportional width of the parietes of the carino-lateral compartment is unknown. I have (but with doubt) given it a distinct specific name, owing to the peculiar character of the furrows on the scuta, and to the large size of the whole shell. In its other characters it comes nearest to A. spongites, excepting in the spur of the tergum, which resembles that of A. sulcata.
The external surfaces of the compartments appear generally to have been smooth; but in several specimens they are studded with the sharp shelly points so common in the genus. A rostrum (Tab. II, fig. 6a), and lateral compartment (fig. 6b), have been figured. The radii are not wide. The basis (fig. 6c) is cup-formed: its edge is either quite smooth, or is very finely crenated. The basis is sometimes quite irregularly perforated, as in the case of several recent species, by numerous minute orifices, which, when the animal was alive, no doubt were covered by membrane. Internally the parietes are feebly ribbed, as in A. spongites. Judging from the dimensions of the separated valves, this species must have equalled and perhaps exceeded in size the largest living species, namely, A. glans, from Australia. Hence we may infer, that the basal diameter probably exceeded .55 of an inch: I may add, that the largest European specimens of A. spongites, from Naples and Portugal, are only .3 of an inch in basal diameter.
Scuta (fig. 6e).—These seem to resemble the scuta of A. spongites in all respects, except
in the external longitudinal ridges standing much further apart, and, consequently, in the furrows being much wider: each ridge is generally double. Although there is a good deal of variability in the character of these ridges in A. undulata, and likewise in A. spongites, I have not seen any form intermediate between them. It must, however, be confessed, that this is an extremely variable character in many sessile cirripedes. Internally the scutum (fig. 6d) is chiefly characterised by the absence of characters, that is, by the slightness of the pits for the muscles, and the little prominence of the articular ridge. In the tergum (fig. 6f), the spur is about half the width of the whole valve, and therefore rather wider than in A. spongites.
Genus—PYRGOMA.
PYRGOMA. Leach. Journal de Physique, tom. 85, 1817.
BOSCIA. Ferussac. Dict. Classique d'Hist. Naturelle, 1822.
SAVIGNIUM. Leach. Zoological Journal, vol. ii, July, 1825.
MEGATREMA. Ib. Ib.
ADNA. Ib. Ib.
DARACIA. J. E. Gray. Annals of Phil. (new series), August, 1825.
CREUSIA. De Blainville. Dict. Sc. Nat., Pl. 116, 1816-30.
NOBIA. G. B. Sowerby, junr. Conchological Manual,1 1839.
Valvæ testæ in unam confluente: basis cyathiformis aut subcylindrica, coraliis affixa: valvæ operculares inter se articulatæ.
Shell formed of a single piece: basis cup-formed, or subcylindrical, attached to corals: scutum and tergum articulated together.
This genus can at once be recognised by the shell consisting of a single piece without sutures, whether viewed externally or internally, and by the cup-shaped basis, attached and often imbedded in corals. The one species, P. Anglicum, found both recent and fossil, together with a closely allied recent species, P. Stokesii, in all the characters derived from the opercular valves, closely resemble Balanus and other ordinary forms, and for this very reason they have some slight claims to be generically separated from the other species of Pyrgoma; for in these latter, the opercular valves seem to have broken loose from all law, presenting a greater diversity in character than do all other species of Balaninæ and Chthamalinæ taken together.
1 The name, Nobia, is given in this work on the authority of Leach, but this must be a mistake, probably caused by some MS. name, (that fertile source of error in nomenclature), having been used.
PYRGOMA ANGLICUM. Tab. II, fig. 7a-7c.
PYRGOMA ANGLICA. G. B. Sowerby. Genera of Recent and Fossil Shells, fig. 7, No. 18, Sept. 1823 (sine descript.).
MEGATREMA (ADNA) ANGLICA. J. E. Gray. Annals of Philosoph. (new series), vol. x, Aug. 1825.
PYRGOMA SULCATUM. Philippi. Enumeratio Molluscorum Siciliæ, Tab. 12, fig. 24, (1836).
— ANGLICA. Brown. Illustrations of Conchology, (2d edit., 1844), Tab. 53, fig. 27-29.
P. testâ abruptè conicâ, purpureo-rubrâ; orificio ovato, angusto; basi porosâ, plerumque è coralio exserta: scuto et tergo subtriangularibus.
Shell steeply conical, purplish red: orifice oval, narrow: basis permeated by pores, generally exserted out of the coral: scutum and tergum sub-triangular.
Fossil in the Coralline Crag (Ramsholt) Mus. S. Wood.
Recent on the south coast of England and of Ireland, (12 to 45 fathoms, Forbes and MacAndrew); Sicily; Madeira; St. Jago, Cape de Verde Islands; generally attached to the edge of the cup of a Caryophyllia, in deep water, but at St. Jago within the tidal limits.
I have considered this fossil as identical with the recent species, but, as may be seen from the following description, it presents several slight differences; yet they are such that I dare not found a new species on only a few specimens thus characterised.
The shell is steeply conical, slightly compressed, with the lower part having rounded, approximate, radiating ribs; these ribs seem to be more prominent in the fossil than in the recent specimens. Colour dull purplish-red. Orifice oval, small, and narrow. The basis is not deeply conical, and occasionally is even flat: in the Crag specimens it is almost wholly imbedded in the coral to which it is attached; but in recent specimens it is generally exserted. Externally the basis is furnished with ribs corresponding with those on the shell. The largest recent specimens which I have seen, from St. Jago, was .22 of an inch in basal diameter; but some few of the British specimens are nearly as large, and one of the fossils from the Coralline Crag a very little larger.
The scuta and terga are of the ordinary shape of these valves in Balanus and its allies. Scutum (fig. 7b) triangular, with the basal margin a little curved and protuberant; adductor and articular ridges distinct from each other, moderately prominent; there is a small hollow for the lateral depressor muscle: in the fossils, the adductor ridge (as figured) is more distinct from the articular ridge, and consequently the cavity for the lateral depressor muscle is wider and less deep than in recent specimens. Tergum, I have not seen a fossil specimen, but have figured a recent valve (7c); it is triangular, with the spur rather narrow, moderately long, placed near, but not confluent with, the basi-scutal angle of the
valve. The basal margin forms an angle rather above a right angle with the spur. Internally the articular ridge and crests for the depressor muscles, feebly developed.
Internal Structure of the Shell and Basis.—Internally, the shell is ribbed, more or less prominently. The lower edge of the sheath, which is reddish, and extends far down the walls, seems always to project freely. In several specimens there were on each side, at the carinal end of the shell, a trace apparently of a suture, which could be perceived only on the sheath. The basis appears always to be permeated by minute tubes or pores, though these are sometimes rather difficult to be seen.
Michelotti, in the 'Bulletin Soc. Géolog.' Tom. 10, p. 141, has named, but not described, a species, viz., Pyrgoma undata, from the northern Italian Tertiary Strata.
Genus—CORONULA.
CORONULA. Lamarck. Annales du Muséum, tom. i, (1802).
DIADEMA. Schumacher. Essai d'un Nouveau Syst., &c., 1817.
CETOPIRUS (SED NON CORONULA). Ranzani. Memoire di Storia Naturale, (1820).
POLYLEPAS. J. E. Gray, (Klein). Annals of Philosophy, (new series), vol. 10, 1825.
Valvæ testæ 6, æquali latitudine; parietes tenues, profundè plicati, plicis cavitates infrà solùm apertas efficientibus; valvæ operculares non inter se articulatæ, orificio testæ multo minores: basis membranacea. Cetaceis affixa.
Compartments six, of equal sizes: walls thin, deeply folded, with the folds forming cavities, open only on the under side of the shell: opercular valves much smaller than the orifice of the shell; when both present not articulated together: basis membranous. Attached to Cetaceans.
The structure of the shell of Coronula is complicated, and has been generally quite misunderstood. Without a long description and several figures it would be impossible to give a true idea of its singular structure; but, in order to make the following description at all intelligible, I must make a few remarks. The wall of each compartment, and therefore of the whole shell, is extremely thin; but strength is gained by its being folded in a very complicated manner, as may be seen in the rostral compartment, Tab. II, fig. 8b, by tracing the wall e to e′, to e″; the folds at their outer ends are elongated into transverse loops, the extremities of which touch each other; consequently, what appears to be the outside of the shell consists only of a portion of the wall, namely, the outsides of the transverse circumferential loops, together with the radii. These loops appear externally like much flattened longitudinal broad ribs. On the other hand, the inside of the shell, in which the body is lodged, consists of the inner ends of the folded walls, lined by the sheath, and by the alæ. The basal edges of the folded walls, in the line of the ray of the circular shell, are oblique; the outer ends, or transverse circumferential loops, having grown downwards at a greater rate than the inner ends. Between each fold of the walls, there is a flattened
cavity, open at the bottom of the shell, and running up to the apex: these cavities are quite external to the cirripede, and are occupied by the epidermis of the whale to which the Coronula is attached: homologically they are only deep longitudinal furrows, and they would still have been furrows, had not the transversely elongated ends of the folds, i. e., the circumferential loops, in all cases, after early growth, grown into close contact. The ends of these loops are generally locked together by rows of minute teeth. In all the species, when young, the wall of each compartment is folded three times, and therefore the whole shell has eighteen folds.
The radii, normally, are only part of the wall, modified by growing against an opposed compartment; and hence the radius in Coronula would have been extremely thin, like the wall, and the sutures between the six compartments excessively weak, had not the radii been specially thickened by numerous sinuous denticulated plates, springing from the inner lamina of the true radius, and running downwards, attached to the folded wall of the compartment to which the radius belongs, and with their free edges pressed against the folded wall of the opposed compartment. Hence the radii may be said to be compound. For the sake of strengthening the sutures, the alæ, also, are very unusually thick: but, notwithstanding their thickness and the thickness of the compound radii, owing to the depth of the folds of wall, they are separated from each other by a considerable space, and the alæ, instead of resting in chief part, as they should do, on the inner lamina of the radius, have to rest on special plates, developed apparently from the sheath. In the upper part of the shell, between the special plates on which the alæ rest, and the compound radii, there are in two of the three recent species, open chambers, six in number, occupied by the ovarian cæca; but in the fossil C. barbara these chambers are almost filled up solidly by shell. I hope that the terms used in the following description may be now in some partial degree rendered intelligible.
CORONULA BARBARA. Tab. II, fig. 8a-8e.
CORONULITES DIADEMA (?) Parkinson. Organic Remains (1811) vol. iii, p. 240, pl. 16, fig. 19.
C. testâ (probabiliter) coroniformi, costis longitudinalibus convexis, aciebus earum crenatis, superficie internâ et externâ cristis transversis asperâ; radiis modicè crassis; spatio inter radios et alas solidè impleto.
Shell (probably) crown-shaped, with longitudinal convex ribs, having their edges crenated, and their surfaces rugged, both externally and internally, with transverse ridges: radii moderately thick; the space between the radii and the alæ solidly filled up.
Fossil in Red Crag, (Bawdsey and Sutton); Mus. S. Wood and Geological Society.
This species, though closely allied to C. diadema and easily confounded with it, I have no doubt is distinct. I owe to the kindness of the Rev. Mr. Image an examination of the original specimen figured by Parkinson; and in Mr. Stuchbury's collection there is a similar and more perfect specimen; both of these resemble C. diadema in general form, but have been too much worn to be positively identified. The following description is drawn up from some compartments collected by Mr. Searles Wood, belonging certainly to three and probably to four individuals, one of which was young; as these specimens agree in all essential respects, I feel pretty confident that the characters, by which the present species differ from C. diadema, are of specific value.
Structure of Shell.—The longitudinal ribs on each compartment (i. e. the circumferential transverse loops), are convex and prominent, as in C. diadema, but they are crossed by more prominent ridges of growth (fig. 8a, 8e) than even in the roughest varieties of that species, so that the surface of the shell is more rugged. In the three recent species—viz., C. diadema, balænaris, and reginæ, the surface of the wall all round the cavities occupied by the whale's skin, is striated only by very fine longitudinal lines; but here, the outer portion, or that (fig. 8d) formed by the transverse loops, is crossed by transverse ridges of growth, like, but less prominent, than those on the external surface of the shell. The minute teeth, along the lines of junction between the transverse loops, are here less regular, and can hardly be said to exist; for the two edges are locked together by what may be more strictly called minute zig-zag ridges (fig. 8d, 8e), than teeth. The exact number of the circumferential plications (fig. 8b) in the wall of the shell is variable, in the same manner as in the three recent species. In the rostrum which has been figured (8b, enlarged twice its natural size), there is a peculiarity, probably accidental, which I have seen in no other specimen—namely, that one of the transverse circumferential loops at the end of one of the original folds of the wall, has ceased to be added to, and therefore may be seen (rather on the right hand of the middle of the figure) to terminate in one of the cavities between two adjoining folds. The sutural edges of the compound radii (d, fig. 8b) are about as thick as, or rather thicker than, in C. diadema; for in the middle part they do not reach to the sheath by about half the thickness of the compartment. In the same manner as in C. diadema and reginæ, each ala here rests, not on the internal surface (as in C. balænaris, and in all other Balanidæ) of the radius, but on a special plate (c, fig. 8b, 8c); but in C. barbara, instead of there being a deep chamber, running up to the apex of the compartment, between the radius and the special plate, this part is filled up almost entirely by solid shell. Although the extent to which this chamber is filled up varies a little, and although its depth varies a little in C. diadema, yet there is a marked difference between the specimens of this latter species, in which the chamber is most filled up, and those of C. barbara, in which it is least filled up. The alæ are thick, as in C. diadema, and their sutural edges have a central ridge, sending off on both sides sinuous crests. The basal margins of the alæ are not short compared with their upper margins, and therefore the whole ala is not wedge-formed (fig. 8c); and in this rather important respect C. barbara
resembles C. balænaris, and differs from C. diadema. The lower edge of the sheath does not seem to have projected freely,—in this respect, also, resembling C. balænaris. From the basal margin of the alæ not being narrow, and from the inner ends of the folded walls being, as it would appear, also broad, I have little doubt that the cavity in which the animal's body was lodged, resembled in shape that in C. balænaris, the membranous basis being larger than the orifice of the shell.
Opercular valves unknown.
Summary.—This species is more nearly related to C. diadema than to the others; but in some points, just specified, it resembles C. balænaris. The characters by which it differs from all the species are, firstly, the more prominent transverse ridges on the external surface of the shell, and more especially on the surfaces bounding the outer sides of the cavities occupied by the epidermis of the whale. Secondly, the character of the teeth, or rather ridges, along the lines of junction between the transverse loops. And, thirdly, the spaces between the radii and the special plates on which the alæ rest, being solidly filled up.
The Coronula bifida is an Italian tertiary species, so named by Bronn, in his "Italiens Tertiär-Gebilde" (1831), p. 126. It is very possible that this may be identical with C. barbara, but Bronn does not seem to have been aware of the absolute necessity of giving minute details in his descriptions of fossil cirripedes. The chief character of C. bifida is thus given:—"Eine tiefe Furche oder Spalte theilt die Längenrippe von oben herab bis zur Hälfte, welche der sonst ähnlichen C. diadema entweder ganz fehlt, oder nur zuweilen kurz angedeutet ist." Had it been stated that the longitudinal ribs were divided from the middle down to the base, instead of from the top to the middle, the description would have been intelligible to me, though the character thus afforded would not have been of specific value, as this dividing of the ribs occasionally occurs in all four species, and is produced by the formation of new folds in the walls.
Family—VERRUCIDÆ.
Cirripedia sine pedunculo: scuta et terga, musculis depressoribus non instructa, ex uno latere tantum mobilia, ex altero cum carinâ et rostro in testam asymmetricam immobiliter conjuncta.
Cirripedia without a peduncle: scuta and terga, not furnished with depressor muscles, moveable only on one side, on the other side united immoveably with the rostrum and carina into an unsymmetrical shell.
Genus—VERRUCA.
VERRUCA.1 Schumacher. Essai d'un Nouveau Syst. Class., 1817.
CLYSIA. Leach. Journal de Physique, tom. 85, July, 1817; Clisia, Leach, Encyclop. Brit. Suppl., vol. iii, 1824; Clitia, G. B. Sowerby, Genera of Recent and Fossil Shells.
CREUSIA. Lamarck. Animaux sans Vertèbres, 1818.
OCHTHOSIA. Ranzani. Memoire di Storia Nat., 1820.
LEPAS ET BALANUS AUCTORUM.
The family of Verrucidæ includes only the above single genus; but it has, I think, as good a claim to be considered a distinct Family as either the Balanidæ or Lepadidæ, that is, either the Sessile or Pedunculated Cirripedes. The two latter Families differ from each other almost exclusively in the nature of the shell or external covering, and in the muscles moving the different portions of it: now Verruca has a very peculiar shell, destitute of all muscles, excepting the adductor scutorum, and composed of only six valves, and these are so unequally developed, that the longitudinal dorso-ventral plane of the body comes to lie nearly parallel to the surface of attachment, instead of at right angles to it. Upon the whole, the Verrucidæ are nearly equally related to the Lepadidæ and Balanidæ; but certainly nearer to the Lepadidæ, than to the sub-family Balaninæ or typical sessile cirripedes; though, on the other hand, if compelled to place Verruca in one of these two Families, I should place it amongst the Chthamalinæ, the other sub-family of Balanidæ. The distinctness of Verruca, though in appearance a sessile cirripede, from the Balanidæ or true sessile cirripedes, is interesting, inasmuch as no member of this latter Family has hitherto been found fossil in any Secondary Deposit, whereas Verruca ranges from the present day to the upper beds of the Chalk near Norwich, and in Belgium; being likewise found in the Glacial Deposits, in the Red and Coralline Crags of England, and in an ancient tertiary formation of Patagonia.
The shell of Verruca has generally been quite misunderstood: it consists, as already stated, of six valves; and these can be proved (as I have shown in my volume published by the Ray Society), by tracing the development of the young shell, to consist of a rostrum and carina, unequally developed on their two sides,—of a scutum and tergum in their normal and moveable condition,—and, lastly, of the scutum and tergum on the opposite side, most singularly modified, immoveably articulated to the rostrum and carina, forming together with them a shell, which is firmly united to the basal membrane, and so to the surface of attachment. It can be shown that the very remarkable modification and
1 According to Bock, in the 'Naturforscher' of 1778, this term was used by Rumph for a Chelonobia, but as it was before the adoption of the binomial nomenclature, according to the Rules, it may be passed over, and does not interfere with the priority of Schumacher.
6
enlargement of the fixed scutum and tergum, is due to the development of a single small portion in each valve, namely, the lower ridge of the articular ridges by which these valves are united together. It is very remarkable that in all the species it seems to be a matter of chance, whether the right or left hand valves undergo this singular modification; consequently, of every valve it is equally likely to find a right-hand or left-hand specimen; and these, though exactly alike, except in being reversed, or in coming from opposite sides of the body, yet, from this very circumstance, and from the fixed valves being of very irregular shapes, are rather perplexing to identify. This short description will, I hope, suffice to make the following descriptions intelligible.
1. VERRUCA STRÖMIA. Tab. II, fig. 9a, 9b.
LEPAS STRÖMIA. O. Müller. Zool. Dan. Prod., No. 3025, 1776.
— — Ib. Zoolog. Dan., vol. iii, Tab. 94, 1789.
— STRIATA. Pennant. British Zoology, vol. iv, Tab. 38, fig. 7, 1777.
DIE WARZENFORMIGE MEEREICHEL. Spengler. Schriften der Berlin. Gesell., 1 B., Tab. 5, fig. 1-3, 1780.
LEPAS VERRUCA. Spengler. Skrifter af Naturhist. Selskabet, 1 B., 1790.
— — ET STRÖMIA. Gmelin. Syst. Nat., 1789.
BALANUS VERUCA. Bruguière. Encyclop. Meth., 1789; Clisia verrucosa, Deshayes, in Tab.
— INTERTEXTUS. Pulteney. Catalogue of Shells of Dorsetshire, 1799.
LEPAS STRIATUS. Montagu. Test. Brit., 1803.
— VERRUCA. Wood's General Conchology, Pl. 9, fig. 5, 1815.
VERRUCA STRÖMII. Schumacher. Essai d'un Nouveau Syst. Class., 1817.
CREUSIA STRÖMIA ET VERRUCA. Lamarck. Animaux sans Vertèbres, 1818.
OCHTHOSIA STROEMIA. Ranzani. Memoire di Storia Nat., 1820.
CLISIA STRIATA. Leach. Encyclop. Brit. Suppl., vol. iii (sine descript.), 1824.
CLITIA VERRUCA. G. B. Sowerby. Genera of Recent and Fossil Shells, Plate.
VERRUCA STRÖMII. J. E. Gray. Annals of Philosophy (new series), vol. x, Aug. 1825.
V. scuto mobili, cristâ articulari inferiore dimidiam brevis cristæ articularis superioris latitudinem non æquante: testâ plerumque longitudinaliter sulcatâ.
Moveable scutum, with the lower articular ridge not half as broad as the short upper articular ridge: shell generally ribbed longitudinally.
Fossil in Glacial deposits of Scotland, Mus. Lyell; Red Crag (Walton, Essex), Coralline Crag (Sutton), Mus. S. V. Wood.
Recent on the shores of Great Britain and Ireland; Shetland Islands; Denmark; Iceland; shores of northern Europe; Red Sea. Attached to shells, laminariæ, rocks, crabs, and floating bark, from low tidal mark to fifty or ninety fathoms.
I have seen a perfect specimen of this species from the Glacial deposits of Scotland, and
separated valves from the Red and Coralline Crags, collected by Mr. S. Wood. The moveable opercular valves have not been discovered; and these are certainly much the most important parts for the diagnosis of the species; but the other valves are tolerably perfect, and are undistinguishable from recent specimens of V. Strömia; therefore, I have ventured, with some hesitation, thus to name these specimens. The fossil specimens all belong to the common variety, having its shell longitudinally ribbed, a character not observed in the four other species of the genus. As an aid to collectors in the Crag, I have thought it would be more serviceable to give a drawing (fig. 9a), from a recent specimen, of all the six valves, separated, but in as nearly as possible their proper relative positions, and likewise of the under side of the fixed scutum and tergum, than to give fac-similes of such valves, in themselves not perfectly characteristic, which have as yet been discovered fossil.
It should be borne in mind, that of the six valves of which figures are here given, it is just as likely that reversed specimens from the opposite side of the body should be found, as these which represent valves taken from a specimen in which the left-hand scutum and tergum were fixed and formed part of the shell.
2. VERRUCA PRISCA. Tab. II, fig. 10a-10c.
VERRUCA PRISCA. Bosquet. Monographie des Crustacés fossiles du Terrain Crét. de Limbourg, Tab. 1, fig. 1-6; 1853.
V. testâ lævi; scuti mobilis cristâ articulari inferiore aliquanto latiore quam superior.
Shell smooth: moveable scutum, with the lower articular ridge somewhat broader than the upper articular ridge.
Fossil in Chalk, Norwich, Mus. J. de C. Sowerby: 'Système Senonien et Maestrichtien,' Belgium, Mus. Bosquet.
M. Bosquet has admirably figured and described the several separated valves belonging to this species, and I owe to his great kindness an examination of some of them. In Mr. J. de C. Sowerby's collection, also, there is a single specimen (fig. 10a), attached to a Mollusc, with the four valves of the shell united together, but without the two moveable opercular valves; it cannot be positively asserted that this is the same species with that of M. Bosquet, but such probably is the case. The opercular valves (fig. 10b, 10c) are necessarily figured from Belgian specimens. It is the English specimen to which I alluded in the Introduction to my 'Monograph on Fossil Lepadidæ.' This species of Verruca is interesting, from being the only known Secondary one, but in itself it is a
very poorly characterised form, and I can point out no important character in the shell by which it can be recognised. The rostrum and carina, which are of nearly equal sizes, are locked together, and likewise to the fixed scutum and tergum, by the usual interfolding plates; the plates between these latter valves seem to have been less developed in M. Bosquet's specimen than in the English. The fixed scutum has a large adductor plate, which seems to have been chipped in M. Bosquet's specimen; this valve and the fixed tergum in all essential respects resemble the same valves in V. Strömia. The surface of the shell is very smooth.
The moveable scutum has its occludent margin considerably arched: the lower articular ridge is broader than the upper ridge, in which respect it resembles the same valve in V. lævigata, a South American form, but the whole valve is not so broad as in that species. There is no adductor ridge on the under surface. The moveable tergum has its upper articular ridge narrow, and slightly produced into a point on the scutal margin: in this latter respect this species, also, resembles V. lævigata, but differs from it in the whole valve not being so broad in proportion to its height.
INDEX
TO
MONOGRAPH ON FOSSIL BALANIDÆ.
BY
C. DARWIN, M.A., F.R.S., &c.
—————
N.B.—The names in Italics are Synonyms.
—————
| PAGE | ||
| Acasta undulata . | 34 | |
| Adna |
35 | |
| Balanus | angulosus . | 21 |
| ,, |
arctica patelliformis . . | 21 |
| ,, | bisulcatus |
26 |
| ,, | calceolus . | 15 |
| ,, | candidus . | 25 |
| ,, | clavatus . . . | 23 |
| ,, | concavus . | 17 |
| ,, | crassus . | 13 |
| ,, | crenatus . | 23 |
| ,, | cylindraceus | 17 |
| ,, | dolosus | 28 |
| ,, | d'Orbignii | 13 |
| ,, | elongatus | 23 |
| ,, | erisma | 29 |
| ,, | geniculatus | 21 |
| ,, | glacialis | 23 |
| ,, | Hameri | 24 |
| ,, | inclusus | 31 |
| ,, | intertextus | 42 |
| ,, | miser | 17 |
| ,, | perplexus | 29 |
| ,, | porcatus | 21 |
| ,, | pustularis | 17 |
| ,, | rugosus | 23 |
| ,, | Scoticus | 21 |
| PAGE |
||
| Balanus | spongicola | 16 |
| ,, | sulcatinus | 26 |
| ,, | sulcatus | 21 |
| ,, | tesselatus | 21 |
| ,, | tintinnabulum | 13 |
| ,, | tulipa | 13, 25 |
| ,, | Uddevallensis | 25 |
| ,, | unguiformis | 29 |
| ,, | verruca | 42 |
| ,, | zonarius | 17 |
| Boscia |
35 | |
| Cetopirus |
37 | |
| Chirona |
12 | |
| Clitia
verruca |
42 | |
| Clysia |
41 | |
| Conopea
ovata |
15 | |
| Coronula
barbara |
38 | |
| Coronulites diadema | 38 | |
| Creusia |
35, 42 | |
| Daracia |
35 | |
| Diadema |
37 | |
| Lepas | balanus | 21 |
| ,, | borealis | 23 |
| ,, | calceolus | 15 |
| ,, | costata | 21 |
| ,, | crispata | 13 |
| ,, | foliacea | 23, 24 |
| PAGE | ||
| Lepas | Hameri | 24 |
| ,, | porcata | 13 |
| ,, | Scotica | 21 |
| ,, |
spinosa | 13 |
| ,, | striata | 42 |
| ,, | strömia | 42 |
| ,, | tintinnabulum | 13, 17 |
| ,, | tulipa | 24 |
| ,, | tulipa alba | 24 |
| ,, | verruca | 42 |
| PAGE |
||
| Megatrema | 35 | |
| Messula | 12 | |
| Nobia | 35 | |
| Ochthosia | 41 | |
| Polylepas | 37 | |
| Pyrgoma Anglicum | 36 | |
| Pyrgoma sulcatum | 36 | |
| Savignium | 35 | |
| Verruca | prisca | 43 |
| ,, | strömia | 42 |
| TAB. I. | ||||||
| Fig. | 1a, | Balanus | tintinnabulum, | nat. size; small specimen. | ||
| 1b, | ,, |
,, |
rostrum, internal view of, large specimen, nat. size. | |||
| 1c, 1d, | ,, | ,, | scutum and tergum, internal views of, from recent specimens, the opercular valves not having been found fossil. | |||
| Fig. | 2a, | Balanus | calceolus, | engraved from a recent specimen, the single fossil having been injured during examination. | ||
| 2b, 2c, | ,, | ,, | scutum and tergum, internal views. | |||
| 2d, | Spur of tergum, variety: all from recent specimens, the opercular valves not having been found fossil. | |||||
| Fig. | 3a, 3b, | Balanus | spongicola, | scutum and tergum, external views. | ||
| 3c, 3d, | ,, | ,, | ,, ,, internal views. | |||
| 3e, | ,, | ,, | shell, enlarged from a recent specimen, the single fossil being young, and having been injured by examination. | |||
| Fig. | 4a, | Balanus
|
concavus, | shell (Coralline Crag specimen). | ||
| 4b, | ,, | ,, | internal view of part of the carina (to the left hand), of the carino-lateral compartment, and of part of the lateral compartment, showing the raised borders (r) on the rostral sides of the sutures in the sheath (Italian Tertiary specimen). | |||
| 4c, | ,, | ,, | shell, var., with longitudinal ridges or ribs not prominent (Coralline Crag). | |||
| 4d, | ,, | ,, | smooth var. (Piedmont specimen). | |||
| 4e, | ,, | ,, | scutum, external view (Coralline Crag). | |||
| 4f, | ,, | ,, | ,, internal view do. | |||
| 4g, | ,, | ,, | tergum, external view do. | |||
| 4h, | ,, | ,, | scutum, internal view (Maryland, U.S.). | |||
| 4i, | ,, | ,, | tergum, internal view do. | |||
| 4k, | ,, | ,, | ,, external view do. | |||
| 4l, | ,, | ,, | scutum, external view (Portugal fossil, and recent). | |||
| 4m, | ,, | ,, | tergum, external view do. do. | |||
| 4n, | ,, | ,, | scutum, internal view do. do. | |||
| 4o, | ,, | ,, | tergum, external view, very large specimen (Turin). | |||
| 4p, | ,, | ,, | scutum, small portion, at the rostral corner, highly magnified, very large specimen (Turin). | |||
| Fig. | 5a, | Balanus porcatus, shell, nat. size (Red Crag). | ||||
| |
5b, | Balanus | porcatus, | small portion of basal margin of wall, much enlarged. | ||
| 5c, | ,, |
,, |
portion of basal plate, much enlarged. | |||
| 5d, 5e, 5f, 5g, | Balanus porcatus, scutum and tergum, external and internal views; engraved from recent specimens, the opercular valves not having been positively found fossil. | |||||
| Fig. | 6a, | Balanus | crenatus, | twice nat. size (Red Crag). | ||
| 6b, | ,, | ,, | small portion of basal margin of wall, much enlarged. | |||
| 6c, | ,, | ,, | portion of basal plate, of unusual thickness, much enlarged. | |||
| 6d, 6e, | ,, | ,, | scutum and tergum, external views from recent specimens. | |||
| 6f, 6g, | ,, | ,, | ,, ,, var., internal views from recent specimens. | |||
| Fig. | 7a, 7b, | Balanus
|
Hameri, | scutum and tergum, external views engraved from Canada glacial specimens, the opercular valves not having been found in Great Britain. | ||
| 7c, 7d, | ,, | ,, | scutum and tergum, internal views of very large and old specimens (Canada and Uddevalla). | |||
TAB. I

BALANUS.
George B. Sowerby
| TAB. II. | ||||||
| Fig. | 1a, | Balanus | Hameri, | lateral compartment, viewed externally. | ||
| 1b, | ,, |
,, | carino-lateral compartment, viewed internally. | |||
| Fig. | 2a, | Balanus
|
bisulcatus, | shell, 1½ nat. size. | ||
| 2b, | ,, | ,, | rostrum, viewed externally, to show the commencement of the folding of the walls, 1½ nat. size. | |||
| 2c, | ,, | ,, | var. plicatus, thrice nat. size. | |||
| 2d, | ,, | ,, | very young shell, probably belonging to this species, enlarged about seven times. | |||
| 2e, 2f, | ,, | ,, | scutum and tergum, external views. | |||
| 2g, 2h, | ,, | ,, | ,, ,, internal views. | |||
| Fig. | 3a, | Balanus | dolosus, | rostrum, much enlarged, viewed internally. | ||
| 3b, | ,, | ,, | portion of basal plate, much enlarged. | |||
| 3c, 3d, | ,, | ,, | scutum and tergum, external views. | |||
| Fig. | 4a, | Balanus | unguiformis, | shell, twice nat. size. | ||
| 4b, | ,, | ,, | shell, var. erisma, twice nat. size. | |||
| 4c, 4d, | ,, | ,, | scutum and tergum, external views. | |||
| 4e, 4f, | ,, | ,, | ,, ,, internal views. | |||
| Fig. | 5a, | Balanus | inclusus, | shell, nearly thrice nat. size. | ||
| 5b, | ,, | ,, | basis of ditto, showing basal edges of the compartments. | |||
| 5c, | ,, | ,, | var., with its rostro-carinal axis elongated. | |||
| 5d, | ,, | ,, | var., ,, ,, ,, showing the narrow furrowed basis and lower portions of the six compartments. | |||
| 5e, | ,, | ,, | var. intermediate between the last two varieties, basal view of, nearly twice nat. size. | |||
| 5f, 5g, | ,, | ,, | scutum and tergum, internal views from a very young specimen. | |||
| Fig. | 6a, | Acasta | undulata, | rostrum viewed externally; spinose var.; twice nat. size. | ||
| 6b, | ,, | ,, | lateral compartment, smooth var., twice nat. size. | |||
| 6c, | ,, | ,, | basal cup. | |||
| 6d, | ,, | ,, | scutum viewed internally. | |||
| 6e, 6f, | ,, | ,, | ,, and tergum, viewed externally. | |||
| Fig. | 7a, | Pyrgoma
|
Anglicum, | shell, viewed from above, four times nat. size. | ||
| 7b, | ,, | ,, | scutum, internal view. | |||
| 7c, | ,, | ,, | tergum, internal view, engraved from recent specimen, this valve not having been found fossil. | |||
| Fig. | 8a, | Coronula | barbara, | rostrum, external view; nat. size. | ||
| 8b, | ,, | ,, | ,, viewed internally; twice nat. size. | |||
| 8c, | ,, | ,, | lateral compartment, internal view. | |||
| 8d, | ,, | ,, | internal view, much enlarged, of small portion of basal margin of folded wall. | |||
| 8e, | ,, | ,, | external view, greatly enlarged, of small portion of surface of folded walls, near the basal margin; (r r) the transverse ridges of growth. | |||
| The following letters of reference apply to all the figures of Coronula: | ||||||
| a, | sheath marked transversely in the upper part by the attachment of the opercular membrane. | |||||
| a′, | ala. | |||||
| b, | furrow on each side of (a), receiving the edge of the thick ala of the adjoining lateral compartment. | |||||
| c, | special plate, on which the ala rests. | |||||
| d, | radius, on the edge it may be just seen to consist of an outer layer (the normal radius), and a much thicker inner part (the pseudo or complementary radius) formed of oblique denticulated septa. | |||||
| e e′ e′′, | basal edge of wall, which from its commencement at e, or e′′, can be followed, folding up to near the basal edge of the sheath, to its termination at e′′ or e. | |||||
| f f, | serrated lines of junction between the folds of the wall. | |||||
| Fig. | 9a, | Verruca | Strömia, | much enlarged, engraved from a recent specimen, only certain valves having been found fossil. | ||
| 9b, | ,, | ,, | fixed scutum and tergum, internal views. The following letters apply to both these figures: | |||
| A, rostrum. | ||||||
| B, carina. | ||||||
| S, moveable scutum, S′, scutum fixed and modified so as to form part of shell. | ||||||
| T, moveable tergum, T′, tergum fixed, forming part of shell. | ||||||
| In S and S′, a is the occludent margin; b, the basal margin; m, the plate to which the adductor muscle is fixed. | ||||||
| In S and S′, the tergal margin is marked by small dashes; (′) being the upper articular ridge, and (′′) the second or lower articular ridge: in S′ (′′) is called the parietal portion of the valve. | ||||||
| In T and T′, the scutal margin is marked by small dashes; (′) being the first and upper articular ridge, hardly distinct from the occludent margin, and called in T′ the occludent rim; (′′) is the second or middle, and (′′′) the lower or third articular ridge, called in T′ the parietal portion of the valve: x is the carinal margin, called in T′ the carinal rim, and z the basal margin. | ||||||
| Fig. | 10a, | Verruca | prisca, | five times nat. size. | ||
| |
10b, 10c, ,, | |
scutum and tergum, external views; engraved from a Belgian Cretacean specimen, the opercular valves not having been found in England. | |||
TAB. II
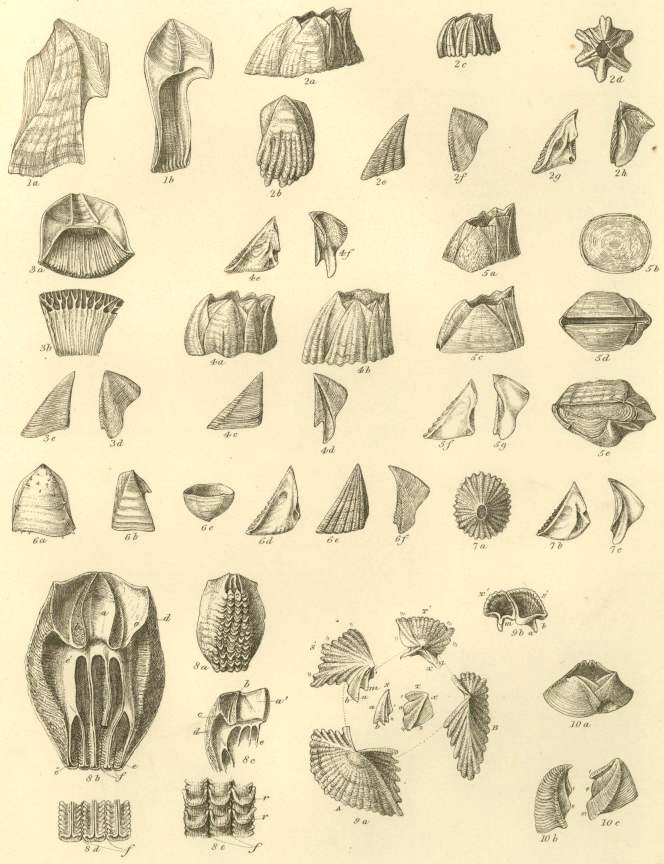
BALANUS, ACASTA, PYRGOMA, CORONULA, VERRUCA.
George B. Sowerby