NO. II. OF PART I.]
[PRICE
8s.
THE
ZOOLOGY
OF
THE VOYAGE OF H.M.S. BEAGLE,
UNDER THE COMMAND OF CAPTAIN FITZROY, R.N.,
DURING THE YEARS
1832 TO 1836.
————————
PUBLISHED WITH THE APPROVAL OF
THE
LORDS COMMISSIONERS
OF HER
MAJESTY'S TREASURY.
————————
Edited and Superintended by
CHARLES DARWIN, ESQ. M.A., F.R.S., Sec. G.S.
NATURALIST TO THE EXPEDITION.
————————————————
FOSSIL MAMMALIA,
BY RICHARD OWEN, ESQ., F.R.S.
PROFESSOR OF ANATOMY AND PHYSIOLOGY IN THE ROYAL COLLEGE OF SURGEONS, LONDON.
————————
LONDON :
SMITH, ELDER AND CO. 65, CORNHILL.
MDCCCXXXIX.
VII.
STEWART AND MOWBRAY, OLD BAILEY.
Preparing for Publication, in One Vol. 8vo.
WITH THE APPROVAL OF THE LORDS COMMISSIONERS OF HER MAJESTY'S TREASURY.
GEOLOGICAL OBSERVATIONS
(with numerous maps and sections)
MADE DURING THE VOYAGE OF H. M. SHIP BEAGLE,
UNDER THE COMMAND OF CAPT. FITZROY, R.N.
ON THE
VOLCANIC ISLANDS
OF THE ATLANTIC AND PACIFIC OCEANS;
AND ON
CORAL FORMATIONS;
Together with a Brief Notice of the Geology of the Cape of Good Hope, and of Parts of Australia.
By CHARLES DARWIN, ESQ., M.A.
SECRETARY TO THE GEOLOGICAL SOCIETY OF LONDON.
————————
SMITH, ELDER AND CO., CORNHILL.
last lumbar vertebra, which presents in addition, the following characters; each transverse process sends off from its posterior margin a thickened and transversely elongated protuberance, which supports a flattened articular surface adapted to a corresponding surface on the anterior part of the transverse process of the first sacral vertebra: it likewise presents on its anterior edge a flattened and rough surface, which is closely attached by ligamentous substance to the opposite part of the transverse process of the penultimate lumbar vertebra. A similar structure exists in the last two lumbar vertebræ of the Rhinoceros, Tapir, and Horse. In the latter animal, anchylosis of these articulating surfaces of the lumbar and sacral vertebræ generally takes place with age, and, judging from the character of the same surfaces in the Hippopotamus, the motion of its lumbar vertebræ upon the sacrum may in like manner become ultimately arrested.
Now in the Macrauchenia, as in the Pachyderms above cited, the transverse processes of the last lumbar vertebræ are of considerable thickness and extent, and are joined by enarthrosis to the transverse processes of the sacrum; but the bony structure of these joints would indicate that they were not subject to be obliterated by anchylosis. The articular surfaces which project from the posterior part of the transverse processes of the last lumbar vertebræ present a regular and smooth concavity, adapted to a corresponding convexity in the transverse processes of the first sacral vertebra. These articulating surfaces have evidently been covered with smooth cartilage; they present a pretty regular transverse ellipsoid form. A view of the three joints by which, independently of the two oblique processes, the last lumbar vertebra of the Macrauchenia was articulated with the sacrum, is given in Plate VIII. fig. 1. The transverse processes of the posterior lumbar vertebra, besides their agreement with those of the Horse and Hippopotamus in the structure just described, also correspond with them in general form, and deviate remarkably from those of the Camelidæ in their great breadth.
It will be seen that the articulations on the body and transverse processes of the last lumbar vertebra of the Macrauchenia differ from the corresponding articular surfaces of the Horse, inasmuch as the middle surface is convex, while the two lateral ones are concave, and these are moreover relatively larger than either in the Horse or Hippopotamus: by this structure the trunk was more firmly locked to that segment of the vertebral column, which receives and transmits to the rest of the body the motive impetus derived from the hinder extremities, which are in all quadrupeds the chief powers in progression; while at the same time the shock must have been diminished by the great extent of interposed elastic cartilages; and a certain yielding or sliding motion would be allowed between the lumbar vertebræ and sacrum.
The anterior oblique processes of the lumbar vertebræ of the Macrauchenia
G
[page] 42 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
(fig. 4, Pl. VIII.) have concave articular facets turned towards, and nearly continued into, each other at their lower extremities; so as to form together a deep semilunar notch, into which the corresponding convex articular surfaces of the posterior oblique processes of the adjoining vertebra (fig. 3, Pl. VIII.) are firmly locked. In the close approximation of the two anterior concave articular facets, which are separated from each other only by a vertical ridge, and a rough surface of about three or four lines in breadth, the lumbar vertebræ of the Macrauchene resemble those of the Horse, and differ from those of the Camel-tribe and Ruminants generally, in which those surfaces are wider apart. In the hook-like form, however, of these articular processes the lumbar vertebræ of the Macrauchene differ from those of the Horse; and resemble those of many Ruminant species, and of the Anoplothere;* but the degree of concavity of the articulating surface is not so great in the Macrauchene. It would be interesting to determine the relations which the lumbar vertebræ of the Macrauchene bear to those of the Palæothere; but the indication which Cuvier gives of the single lumbar vertebra, of which he had cognizance in the latter genus† is too slight to enable me to enter upon the comparison.
The whole length of the lumbar region in the Macrauchene is twenty inches. When the bodies of these vertebræ are naturally adapted together, they form a slight curve, indicating that the loins of the Macrauchene were arched, or bent downwards towards the sacrum. That the lumbar vertebræ were rigidly connected together, or but slightly flexible, is evident from the flatness of the articular surfaces of the vertebral body, and by the circumstance of ossification having extended along the anterior vertebral ligaments, and produced an anchylosis between the fourth and fifth lumbar vertebræ; (fig. 2, c, Pl. VIII.) This kind of ossification is frequent in aged horses, and I have seen an example of a similar anchylosis of the lumbar vertebræ, by abnormal deposition of bone in their anterior ligaments, in the skeleton of a Hippopotamus preserved in the Senkenbergian Museum, at Frankfort.
In preparing the preceding account of the cervical and lumbar regions of the vertebral column of the Macrauchene, I have felt frequently a strong desire to enter into a comparison between them and the corresponding vertebræ of the extinct Pachyderms of the Paris Basin. Some of these, as the Anoplotherium gracile, in the length and slenderness of the cervical vertebræ, resemble both Auchenia and Macrauchenia; others, as the Palæotherium minus, and probably the rest of the genus, resemble the Camelidæ and Macrauchenia in having seven lumbar vertebræ. Cuvier points out the resemblance which the atlas of the Anoplothere bears to that of the Camel, and especially of the Llama;‡ but he
* Cuvier, Ossemens Fossiles, iii. p. 238.
† Loc. cit. p. 234.
‡ Loc. cit. p. 235.
expressly notices the existence of the canals for the vertebral artery in the fifth or sixth cervical vertebra of the Anoplotherium commune.* Do the cervical vertebræ—say from the third to the sixth inclusive—of the Palæotherium present an imperforate condition of their transverse processes, or exterior part of their sides? Cuvier, who seems not to have been aware of this peculiarity in the Camelidæ, merely notices the absence of these arterial foramina in the last cervical vertebra of the Palæotherium minus,† which, unfortunately for the comparison I am desirous of establishing, is that which most commonly presents this imperforate condition in the Mammalia generally. As, however, the cervical vertebræ of the Palæothere had the anterior articular surface of the body convex, and the transverse processes produced into descending laminæ, it is most probable that they corresponded with the cervical vertebræ of the typical Pachyderms in the condition of their arterial foramina.
The sacrum and ossa innominata in the present specimen of Macrauchenia are very imperfect; but sufficient is preserved to show that the sacrum was anchylosed to the ilia: the lower boundary of this anchylosis is marked below by an external ridge, and by vascular canals and grooves in the substance of the bone, as in the Hippopotamus. The body of the sacrum is lost, but the smooth articular convexities upon the transverse processes adapted to the articular depressions of the last lumbar vertebra are fortunately preserved.
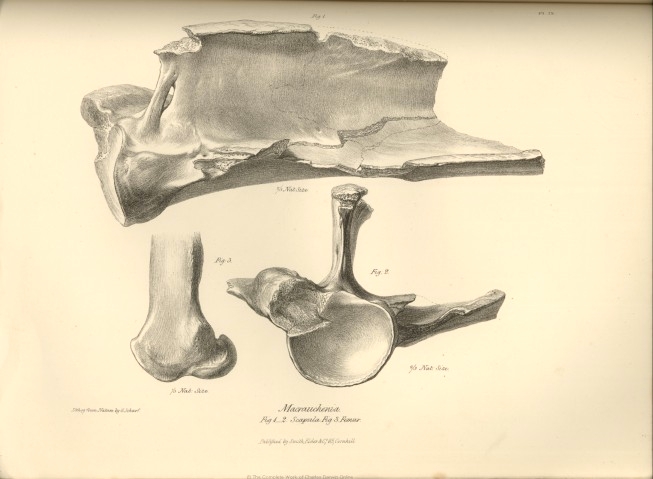
The remains of the anterior extremity of our Macrauchenia include fragments of a left scapula; the proximal extremities of the anchylosed bones of the right antibrachium; the metacarpal and most of the phalangeal bones of the right fore-foot. The first-mentioned fragments, include the head and neck of the scapula, a small part of its body with the beginning of the spine, the coracoid process, and the nearly entire glenoid cavity. This articular surface (fig. 2, Pl. IX.) resembles in its general form, and degree of concavity, that of the Camel and Rhinoceros, and is deeper than in the Hippopotamus. The coracoid process is represented by a slightly produced rough, thick, and obtuse tuberosity, situated closer to the glenoid cavity than in the Camelidæ or Rhinoceros, and having almost the same relative position and size, as in the Palæotherium crassum. The superior border or costa of the scapula presents much variety in the Ungulate quadrupeds with which we have to compare the Macrauchenia. In the Ruminants its contour forms behind the coracoid a concave sweep, which advances close to the spine of the scapula. In the Camel and Horse the marginal concavity is shallower, and the distance of the superior costa from the spine of the scapula is greater; the extent of the supra-spinal fossa increases in the true Pachyderms, and the Macrauchene agrees with them in this structure. In the Tapir, how-
* Loc. cit. p. 237.
† Loc. cit. p. 232.
[page] 44 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
ever, the contour of the superior costa is broken by a deep round notch immediately behind the coracoid: in the Hippopotamus this process arches in a slight degree backward over a corresponding but wider and shallower notch. In the Palæotherium crassum the concavity of the superior costa, behind the coracoid, is as slight as in the Rhinoceros; but in the Macrauchenia the superior costa of the scapula begins to rise or stretch away from the parallel of the spine, immediately behind the coracoid process. The modifications of the spine of the scapula which characterize respectively the Ruminants and Pachyderms have been clearly and concisely set forth by Cuvier, who at the same time points out the exceptional condition which the Camelidæ present in the production of the acromial angle. It was with peculiar interest and care, therefore, that I reunited all the fragments of the scapula of the Macrauchene, in the hope of gaining from this part of the skeleton as decisive evidence of an affinity to the Camel as the cervical vertebræ had afforded. It unfortunately happens, however, that the part of the scapula most important in this comparison is broken off; yet from this very circumstance, combined with a slight inclination forwards of the anterior margin of the spine immediately beneath the fractured acromion, and from the thickness of the fractured surface, we may infer that the acromial angle of the spine was more produced than in the ordinary Ruminants, although evidently in a less degree than in the Camel tribe. The Macrauchenia, however, surpasses these aberrant Ruminants, and equals the Pachyderms in the elevation and extent of its scapular spine: but this process commences about half an inch behind the glenoid cavity, and rises at once to the height of three inches above the plane of the scapula; in which structure we may trace the same tendency to the Ruminant type, as is manifested in the scapula of the Hippopotamus and Anoplotherium; for in most other Pachyderms the spine increases gradually from its extremities to the middle part. The anterior margin of the spine beneath the short acromion is perforated by an elliptical fissure measuring ten lines, by three lines. The extent of the spine which is preserved, measures eight inches and a half; it is a thin and nearly straight plate of bone, expanding into a thick and rugged upper margin, which slightly over-arches the inferior fossa, (fig. 1, Pl. IX.) In its general form and proportions the spine of the scapula in Macrauchenia presents the nearest resemblance to that of the Hippopotamus; but its origin is closer to the articular surface of the scapula than in this, or any other Pachydermal or Ruminant genus.
The portion of the antibrachium of the Macrauchenia which is preserved, presents a condition of the radius and ulna intermediate to those which respectively characterize the same bones in the Pachyderms and Camels. In the former the radius and ulna are separate bones, united in the prone position by ligament, yet so that the movement of supination cannot be performed; in the
ordinary Ruminants they are partially joined by bony confluence, which rarely extends to the proximal extremities; in the Camel and Llama the anchylosis of the radius and ulna is so complete, that no trace of their original separation can be perceived, and the olecranon appears but as a mere process of the radius.
In the Macrauchenia the anchylosis of the radius and ulna is also complete, but the boundary line of the two originally distinct bones is very manifest, and the proportion which each contributes to the great articulating surface for the distal end of the humerus is readily distinguishable. About a sixth part of this surface is due to the head of the radius, which enters into the composition of the anterior and outer part of the articulation, and its extent is defined by a depressed line describing a pretty regular curve, with the concavity directed forwards and a little outwards, (a, fig. 1, Pl. X.) Just below the articular surface a strong triangular rugged protuberance projects from the front of the head of the radius, for the attachment of the tendon of the biceps. The line of separation of the radius and ulna is indicated on the inner side of the head of the radius by a deep and narrow fissure extending downwards from below the anterior part of the articulating surface; and on the outer side by a broad groove leading upwards to a deep pit near the proximal end of the antibrachium. We may see by the direction of the head of the radius which is thus defined, that it crosses obliquely in front of the ulna, as in the Elephant, Hippopotamus, and other Pachyderms, and that the bones are anchylosed in the prone condition: below this fissure and groove, which mark the interosseous line, the radius and ulna become blended together into one compact bone, which is flattened from before backwards, with a well marked ridge on the outer side; and excavated by a single medullary cavity, the compact walls of which present a general thickness of one-third of an inch.
The proximal articular surface or sigmoid cavity of the antibrachium, constituted as above described, resembles that of the Palæothere, Tapir, and the generality of the Pachyderms in having two depressions, instead of three, as in the Anoplothere, and Ruminants. The Hippopotamus has a slight tendency to the latter structure, which is also less marked in the Camel than in the ordinary Ruminants. In its general form the sigmoid cavity of the Macrauchene resembles that of the Hippopotamus more than that of the Camel. In the Camel this articular surface is traversed transversely by a broad, shallow, and slightly roughened tract, which divides the smooth surface of the joint into two parts, one forming the anterior horizontal surface due to the conjoined radius and ulna, the other forming the vertical concave surface on the anterior part of the base of the olecranon. In the Hippopotamus there is, as it were, an attempt at a similar division of the articulating surface at the proximal end of the antibrachial bones ; a deeper and rougher depression encroaches upon the articulation from its outer side, but stops when it has reached half-way across. In the
[page] 46 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
Macrauchenia the roughened surface, (b. fig. 1, Pl. X.) commencing also at the outside, extends only one-third of the way across the articular surface: it is, however, as shallow as in the Camel. The articular surface on the anterior part of the base of the olecranon is broader in the Hippopotamus than in the Camel; but in the Macrauchene it is twice as broad as in the Hippopotamus. The size of the olecranon in the Macrauchene exceeds that of the Hippopotamus, and à fortiori that of the Camel: indeed in its general magnitude the Macrauchenia must have fully equalled the largest Hippopotamus; but it no doubt had a more shapely, and less broad and bulky trunk. The olecranon of the Macrauchenia differs in shape, both from that of the Camel and Hippopotamus; it terminates above in a three-sided cone with an obtuse apex; and presents a well marked protuberance at the outer side of the base, which is not present in either the Camel or Hippopotamus. There is also a strong rugged ridge on the back part of the olecranon which makes an angle before sinking into the level of the ulna below.
The confirmation of the close affinity of the Macrauchenia to the Pachydermatous Order, which the structure of the cervical vertebræ alone might have rendered very doubtful, is afforded by the bones of the right fore-foot (Pl. XI.); these are fortunately in so perfect a condition, as to make it certain that this interesting quadruped had three toes on the fore-feet, and not more; and that the fully developed metacarpal bones are distinct, and correspond in number with the toes, and are not anchylosed into a single cannon bone, as in the Ruminants. The bones preserved are the metacarpals, proximal phalanges, and middle phalanges of each of the three toes, and the distal phalanx of the innermost toe.
The proximal end of the innermost metacarpal bone presents three articular surfaces; the middle facet is the largest, and the two lateral ones slope away from it at an angle of 45°. The middle facet is broad and slightly convex in front, narrow and concave behind; the distal articular surface of the trapezoides must have corresponded with this surface; the outer facet is narrow, flat, extends from the fore to the back part of the head of the bone, and must have been adapted to a corresponding surface on the os magnum; the inner facet is the smallest, presents a triangular form, and is situated towards the back part of the head of the metacarpal bone; it indicates the existence of a rudimental metacarpal bone, or vestige of a pollex. Below the outermost of the lateral surfaces there is a crescentic articular surface with its concavity directed outwards and downwards (fig. 2, Pl. XV.), against which a corresponding convex articular surface of the middle metacarpal abuts, (fig. 3, Pl. XV.) External to this surface the proximal end of the middle metacarpal bone presents two articular surfaces for the carpus; the larger one, which was adapted to the os magnum, is hori-
zontal, broad and convex before, narrow and concave behind ; the outermost facet is a small triangular surface inclined downwards to the level of the articulating surface of the outermost metacarpal. It also presents a posterior vertical articular surface for a sesamoid bone. The proximal extremity of the outer metacarpal bone is joined to the middle metacarpal, not by one semilunar surface, but by two separate articulations of small size (fig. 4 and 5, Pl. XV.); it presents a single large slightly convex articular surface for the os magnum, of an irregular semicircular form, with the convexity of the curve turned outwards.
The metacarpus increases in breadth as it approaches the phalanges; the two lateral metacarpals bending slightly away from the middle one, and expanding towards their distal extremities: the middle bone presents a symmetrical figure except at its proximal extremity (fig. 2, Pl. XI.) The distal articulating facet of each of the metacarpal bones extends so far upon both the anterior and posterior surfaces as to describe more than a semi-circle (fig. 3, Pl. XI.); in the two lateral metacarpals it is traversed throughout by a longitudinal convex ridge dividing it into two equal lateral parts; the ridge is most produced on the posterior half of the joint (fig. 4, Pl. XI.): in the middle metacarpal this ridge subsides before it reaches the anterior part of the articular surface.
The proximal extremity of the middle proximal phalanx presents a posterior notch corresponding to the above partially developed ridge : the proximal extremities of the lateral phalanges are traversed by a middle longitudinal depression, and two lateral shallow concavities (fig. 6, Pl. XI.;) but these are of such an extent as to be in contact with only a part of the convexity above, which therefore was doubtless adapted to a sesamoid bone on each side of the longitudinal ridge. The structure of the above described joints proves that the motion of the toe upon the metacarpus was much freer and more extensive than in the Rhinoceros, which is the only existing Ungulate mammal which presents the tridactyle structure in the fore-foot. In this species the metacarpo-phalangeal articulations exhibit only a slight trace of the longitudinal ridges and grooves which are confined to the posterior part of the joint; these are more developed in the Camelidæ ; but the Hog and Horse in this respect approach nearer to the Macrauchene, though the structure of the metacarpo-phalangeal joints in the Hog falls far short of the compactness and strength combined with freedom of play in flexion and extension which distinguish those of the Macrauchene. The Palæotherium medium most resembles the Macrauchene in the structure of the trochlear metacarpo-phalangeal joints; but both in this species,* and the Pal. crassum† the articular surface at the distal end of the metacarpal is relatively narrower than in the Macrauchenia; moreover all the species of the extinct Palæothere differ from the Macrau-
* See Ossem. Fossiles, Pl. XX. fig. 3.
† Loc. cit. Pl. XXII. fig. 6.
[page] 48 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
chene in the greater size and strength of the middle as compared with the lateral metacarpals.
The articulation at the distal extremity of the proximal phalanges (fig. 5, Pl. XI.) is simple, and not divided into two pulleys by a longitudinal ridge; it is slightly concave from side to side; but in its extent upon the anterior and posterior surfaces of the bone indicates a freedom of flexion and extension of the toes, which harmonizes with the structure of the joint above.
The proximal articulating surfaces of the second phalanges (fig. 7, Pl. XI.) corresponds of course to those to which they are adapted; they are, however, characterized by sending upwards an obtuse process from the middle of their anterior margin. The distal articulating surfaces (fig. 8, Pl. XI.) resemble those of the proximal phalanges, but extend further upon the back part of the phalanx than the front, indicating the more horizontal position of the second phalanges.
The last phalanx, does not resemble the neatly defined ungulate phalanges of the Ruminantia, and Solipedia, but has the irregular form characteristic of those of the Pachydermata. It is wedge-shaped, broader than it is long, with a rugged surface, except where it plays upon the distal end of the second phalanx, where it is slightly concave in one direction, and convex in the other, (figs. 7 and 9, Pl. XI.) A portion of this phalanx extends backwards behind the articular surface, as in the corresponding bone of the Palæothere and Rhinoceros.
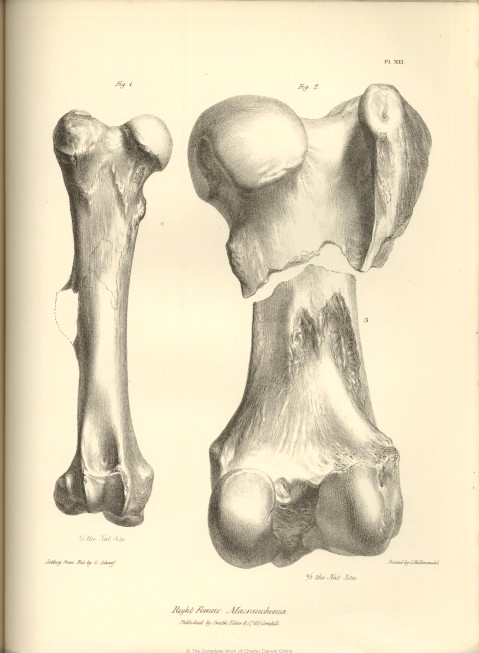
The femur of the Macrauchenia (fig. 1, Pl. XII.) is full two feet in length, and consequently longer than in any known Camel or Rhinoceros; as compared with its transverse diameter it is much longer than the femur of the latter animal: in the proportion of its breadth to its length, and the expansion of its extremities as compared with the diameter of the shaft, it more resembles that of the Camel. The femur of the Giraffe deviates from that of the Macrauchenia in the excessive expansion of its distal extremity. But the most striking evidence deducible from this bone, of the affinity of the Macrauchenia to the true Pachydermatous type is afforded by the evident traces of a third trochanter, the outline of which is conjecturally restored in the figure. Of the Pachyderms which have this characteristic structure, the extinct Palæothere offers the nearest resemblance to the Macrauchene in the general form and structure of the femur.
The head of the femur in the Macrauchene (fig. 2, Pl. XII.) presents the form of a pretty regular hemisphere; it is less flattened above, and is directed more obliquely inwards than in the Palæothere : the neck supporting it does not project so far from the shaft as in the Palæothere or Tapir, but farther than in the Camel. The great trochanter rises above the level of the head; in which structure and in the depression between the head and trochanter, the femur of the Macrauchene offers a character intermediate between the Tapir or Palæothere, and the Camel. The lesser trochanter is a slight projection from a ridge
of bone which is continued from the under part of the head of the femur to the inner surface of the shaft. In the Palæothere the lesser trochanter is situated more towards the posterior surface of the femur ; so that, in this particular, the Macrauchene approaches nearer to the Camel. Cuvier makes no mention of the condition of the depression for the ligamentum teres in the Palæothere. Among existing ordinary Pachyderms the Hippopotamus presents no trace of the insertion of a ligamentum teres in the head of the femur; in the Camel the place of its insertion is indicated by a well-marked circumscribed pit; in the Tapir a similar circular depression is situated close to the inferior margin of the articular convexity. The ligament was undoubtedly present in Macrauchenia, but the place of its insertion is a broad and deep notch leading from the under and back part of the head of the bone a little way into its articular surface: this I regard as another of those interesting transitional structures with which the remains of the Macrauchenia, few and imperfect though they unfortunately are, so freely abound.
The femur of Macrauchenia, in the flatness of the back part of its neck, and the elongated form of the post-trochanterian depression, resembles that of the Camel rather than that of the Palæothere ; and the same resemblance is shown in the cylindrical figure, straightness, and length of the shaft. The depth of the trochanterian depression, and the incurvation of the strong ridge continued downwards from the great trochanter are individual peculiarities in the Macrauchenia.
A great part of the third trochanter is broken off; but from the remains of its base we see that it had the same relative size as in the Palæothere; but it is situated at the middle of the shaft of the femur, and consequently lower down than in the Palæotheres and Tapirs. In the general form and relative size of the condyles at the distal extremity of the femur (fig. 3, Pl. IX. and XII.) the Macrauchene is intermediate to the Camel and Palæothere, but resembles more the latter. In the articular surface for the patella, it deviates somewhat from the Palæothere, having this part longer in proportion to its breadth, more regularly and deeply concave from side to side, and with its lateral boundaries more sharply defined. In all these points the Macrauchene approaches the Camel: the same affinity is shown in the protuberance above the inner condyle; but in the extent of the posterior projection of this condyle (fig. 3, Pl. IX.) it exceeds the Camel and Palæothere, and displays an intermediate structure between these species and the Hippopotamus.
There is a rough crescentic depression above the outer condyle where the linea aspera begins to diverge; the corresponding depression is deeper in the Hippopotamus, while in the Camel it is represented by a roughened surface only, which is not depressed. In the fossa between the rotular articulation and the external condyle the Macrauchene resembles the Camel: the interspace of the
H
[page] 50 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
condyles is relatively wider than in the Camel, and the process above the inner condyle is more angular; in both these respects the Macrauchene inclines towards the Palæothere.
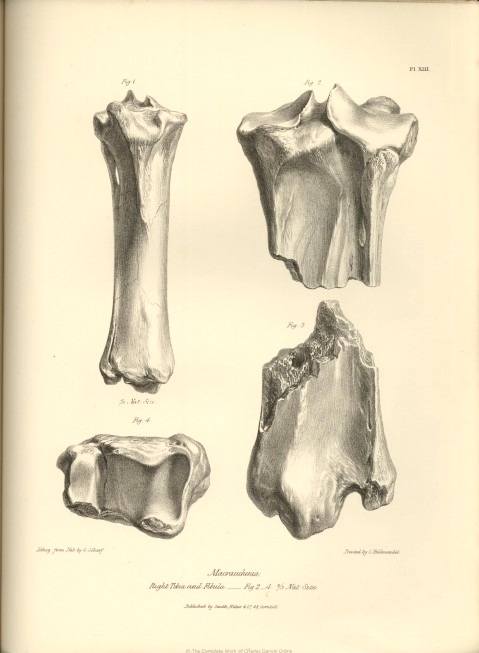
In the structure of the bones of the leg of the Macrauchenia we find the same transitional character which is afforded by the definable limits of the anchylosed bones of the fore-arm. In the Pachyderma the fibula is an entire and distinct bone. In the Ruminantia, with the exception of the small Musk-deer, and, in an inferior degree, the Elk, the fibula appears only as a short continuous process sent down from the under part of the external condyle of the tibia. In the Camel tribe the only trace of the fibula in the bones of the leg, is this process in a still more rudimental state. In the Macrauchenia the fibula is entire, but is confluent with the tibia through nearly its whole extent : the proximal part of the fibula is well defined; its head is anchylosed to the outer condyle of the tibia, but the shaft is continued free for the extent of nearly two inches, and then again becomes confluent with the tibia, forming apparently the outer ridge of that bone. About five inches from the distal end of the tibia this outer ridge becomes flattened by being, as it were, pressed against the tibia, and the anterior and posterior edges are raised above the level of the tibia; beyond this part the limits of the fibula begin again to be defined by deep vascular grooves. The outer side of the distal end of the fibula is excavated by a broad tendinous groove. The fibula and tibia are distinct bones in both the Palæothere and Anoplothere, as in the Pachyderms. It is to the former genus, however, especially Pal. magnum, that the Macrauchene presents the nearest approach in the general form of the tibia, the principal bone of its leg: but in the Macrauchene the tibia is relatively shorter, and thicker, and is straighter and less expanded at its extremities, especially the upper one, than in any of the Palæotheres.
The mesial boundaries of the two superior articulating surfaces of the tibia are raised in the form of ridges, which are separated by a deep groove; of these ridges the external is the highest, as in Pal. magnum : but the articular surfaces in the Macrauchene slope away from these ridges more than in the Palæotheres. The rotular or anterior tuberosity of the tibia is more produced, and rises higher than in the Palæotheres; the ridge continued downwards from this process is more marked in the Macrauchene, and its limits are better defined: the shaft of the tibia below the ridge is also more flattened in the antero-posterior direction than in the Palæothere. The configuration of the back part of both proximal and distal extremities of the tibia are so clearly and accurately given in figures 2 and 3, Pl. XIII., as to render verbal description unnecessary. Neither the text nor the figures in the 'Ossemens Fossiles' afford the means of pursuing the comparison between the Macrauchene and Palæothere in these particulars; and I proceed,
therefore, to the consideration of the inferior articulating surface of the bones of the leg (fig. 4, Pl. XIII.)
Since, of the hind foot, we possess in the present collection only a single tarsal and metatarsal bone, the structure of the distal articular surface of the tibia is attended with peculiar interest, because we are taught by Cuvier that it reveals to us in the Ungulate animals the didactyle or tridactyle structure of the foot. In the Ruminants this articular surface is nearly square, and extended transversely between two perpendicular malleoli, while in the Pachyderms with three toes to the hind-foot the articular surface of the tibia is oblique, and is divided into two facets between the perpendicular malleolar boundaries. Now in the Macrauchenia, although the two bones of the leg are anchylosed together, the extent of that part of the tarsal articular surface which is due to the tibia is indicated, as in the case of the radius in the joint of the fore-arm, by a groove; and we are thus able to compare this surface with the distal articular surface of the tibia in the Palæothere and Anoplothere. It presents in the Macrauchenia a very close resemblance with that of the Palæotherium magnum,* being divided into two facets by a convex rising, which traverses the joint from behind forwards; but the ridge is narrower, the internal facet somewhat deeper, and the external oblique surface rather flatter than in the three-toed Palæothere. In the portion of the tarsal articular surface due to the fibula, we find, however, a more marked deviation from the Palæothere, and an interesting correspondence with the Anoplothere, in the inferior truncation and horizontal articular surface which is continued upon the lower extremity of the fibula, at right angles with the vertical malleolar facet which forms the outer boundary of the trochlea of the astragalus: this articular surface unerringly indicates a corresponding articular projection in the calcaneum, which, therefore, although the bone itself does not form part of the present collection, we may conclude to differ from the calcaneum of the Palæotherium, and to resemble that of the Anoplotherium, in this particular at least.
The valuable indication which the distal articular surfaces of the anchylosed tibia and fibula have given of the correspondence of the hind-foot with the fore-foot of the Macrauchenia, in regard to the number of the toes, receives ample confirmation from the astragalus, which, of all the bones in the foot, is the one that an anatomist would have chosen, had his choice been so limited, and which most fortunately has been secured by Mr. Darwin, in a very perfect state, in the present instance. I have compared this astragalus with that of the Giraffe, and other Ruminants, the Camel, the Anoplothere, the Horse, the Hog, the Hippopotamus, Rhinoceros, Tapir, and Palæothere : it is with the Pachyderms having three toes to the hind-foot, that the Macrauchenia agrees in the main distinguishing
* See Ossem. Foss. iii. Pl. XXVI. fig. 5.
[page] 52 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
characters of this bone; its anterior articular surface, for example, is simple, and not divided into a double trochlea by a vertical ridge: lastly, it is with the astragalus of the Tapir and Palæothere that it presents the closest correspondence in the general form and the minor details of structure, and with these Pachyderms, therefore, I shall chiefly limit the comparison of the Macrauchenia, in regard to the bone in question. If the upper or tibial articular surface (fig. 5, Pl. XIV.) be compared with that in the Palæotherium magnum (Ossem. Foss. Pl. LIV. fig. 2,) it will be seen, that the general direction of that surface is more parallel with the axis of the bone in Macrauchenia. In the Palæotherium it is turned a little towards the outer or fibular side, and in the Tapir the general direction of the same surface is placed still more obliquely. The anterior border of this articulating surface is broken by a semicircular notch in the Palæothere ; in the Tapir it describes a gentle concave curve, and the Macrauchene resembles the Tapir in this respect. The chief difference between the astragalus of the Tapir and the Palæothere, when viewed from above, obtains in the relative length of the bone, anterior to the tibial articulating surface: the Macrauchene presents, in this respect, an intermediate structure, but differs from both in the greater extent of the tibial side of this part of the astragalus.
If we next direct attention to the anterior or scaphoid articular surface, (fig. 3, Pl. XIV.) and compare it with that of the Palæotherium magnum, (fig. 4, Pl. liv, Ossem. Foss.) it will be seen, that it presents in the Macrauchenia an oval, and in the Palæotherium an irregular quadrangular form: in the Macrauchenia, this surface is uniform or undivided, and is gently convex, except at its lower part; while in the Palæothere it is divided by an oblique ridge into a broad internal facet for the scaphoid bone, and a narrow internal surface for articulation with the os cuboides; the larger surface is also concave transversely, and slightly convex vertically: in the Tapir, the anterior surface of the astragalus deviates still further from that of the Macrauchenia, both in general form, and in the proportion of the cuboidal facet. In the didactyle Anoplotherium, Camel, and true Ruminants, where the cuboides presents a large relative size, a still greater proportion of the anterior surface of the astragalus is devoted to the articulation with this bone, and is separated from the scaphoid surface by a well-developed vertical ridge. The Macrauchenia presents, therefore, the extreme variation from this type;—and should the entire tarsus hereafter be discovered, it will doubtless be found, that the os cuboides is articulated posteriorly to the os calcis exclusively.
The external surface of the astragalus of the Macrauchene, (fig. 1. Pl. XIV,) is longer in proportion to its vertical extent than in the Tapir or Palæothere: the articular surface for the fibular malleolus is less curved. Between this surface and the anterior facet the bone is excavated by a deep notch, both in the Tapir and Palæothere ; but in the Macrauchenia by a gentle concavity. Beneath the
malleolar articular smooth surface in the Palæothere there is a deep pit; in the Tapir a shallow one; but in the Macrauchenia we observe only a smooth and slightly convex triangular surface. If we compare the inner surface of the astragalus in these three animals, we shall find the existing Tapir again forming a transition between the two extinct genera. In the Palæothere, a round protuberance projects from the anterior part of this surface: in the Tapir, we observe a gentle rising of the bone in the same part, while in the Macrauchene (fig. 2) the surface of the bone is level at this part. The margin of the tibial malleolar articular surface, which is very slightly raised in the Macrauchene, is more developed in the Tapir, and still more so in the Palæothere, where it forms a ridge, overhanging the rough outer side of the bone. Near the lower part of this surface we observe a small but deep depression in the Palæothere ; there is a shallower one in the corresponding part in the Tapir; and the depression is still wider and shallower in the Macrauchenia. In the Palæothere the astragalus articulates by three surfaces with the os calcis, posteriorly by a large concave surface, externally by a longitudinal sub-elliptic surface, and anteriorly by a thin transverse facet: in the Macrauchene (fig.4) two only of these surfaces are present, viz. the concave and the longitudinal one, the anterior transverse surface being wanting: in the Tapir, the transverse surface is present, but is confluent with the longitudinal one. The posterior surface is relatively larger and deeper in the Macrauchene than in the Palæothere, and approaches nearer to the triangular than the oval form : the longitudinal surface is placed more obliquely, and is truncated anteriorly. In the Tapir this surface is confluent with the scaphoid articular surface, but it is separated therefrom by a narrow strip of bone in both the Palæothere and Macrauchene. It is satisfactory to find in the bone, which marks most strongly the affinity of Macrauchenia to Palæotherium, so many easily recognizable differences, because the structure of the cervical vertebræ in the latter genus is too imperfectly known, to allow us to predicate confidently a distinction between it and Macrauchenia in that particular; the difference, however, which they present in the condition of the bones of the fore-arm and leg, forbids their being considered as generically related.
There remains to be noticed only a single fractured metatarsal bone (fig. 1. Pl. XV.) This, from its bent and unsymmetrical figure, is evidently not a middle one, and having the side of the proximal end, which was articulated to the adjoining metatarsal in a nearly perfect state, it enables us to refer it with certainty to the hind-foot, since it does not agree with any of the corresponding surfaces at the proximal extremities of the metacarpal bones. It remains then to be determined, whether it is an external metatarsal of the right-foot, or an internal one of the left-foot, the general curvature of these being in the same direction. With neither of these bones in the Tapir does our metatarsal agree, since it has but one articular facet on the lateral
[page] 54 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
surface of its proximal end, while the outer metatarsal of the right-foot of the Tapir, with which, in other respects, it most closely corresponds, has two articular surfaces. In the cast of a hind-foot of a Palæothere, I find that the outer metatarsal bone closely agrees with this metatarsal bone of the Macrauchene, in the structure just alluded to: the articulation with the middle metatarsal being by a single sub-oval facet, which stands out a little way from the surface of the bone: the articular surface in the Macrauchene presents a similar form and condition, and is similarly situated to that in the Palæothere, being at the posterior part of the lateral surface, and a little below the superior or tarsal articular surface. The bone expands towards its distal end, which corresponds in structure with those of the two lateral metatarsals in the fore-foot, in being completely divided into two trochlear surfaces by a well-developed median ridge, and in having the posterior half of this ridge suddenly produced, so as to project about two lines further from the trochlear surface than the anterior part of the same ridge. In both the Tapir and Palæothere this anterior part of the ridge is wholly suppressed, and the posterior is much more feebly developed than in the Macrauchenia. The metatarsal bone here described is of exactly the same length with the internal metacarpal bone, and proves, in conjunction with the proportions of the astralagus, that the fore and hind feet of the Macrauchenia were of equal size.
Thus then we obtain evidence, from a few mutilated bones of the trunk and extremities of a single representative of its race, that there once existed in South America a Pachydermatous quadruped, not proboscidian, which equalled in stature the Rhinoceroses and Hippopotamuses of the old world. But this, though an interesting and hitherto unsuspected fact, is far from being the sum of the information which is yielded by these fossils. We have seen that the single ungueal phalanx bespeaks a quadruped of the great series of Ungulata, and this indication is corroborated by the condition of the radius and ulna, which are fixed immoveably in the prone position. Now in the Ungulated series there are but two known genera,—the Rhinoceros and Palæotherium,—which, like the quadruped in question, have only three toes on the fore-foot. Again, in referring the Macrauchenia to the Tridactyle family of Pachyderms, we find, towards the close of our analysis, and by a detailed comparison of individual bones, that the Macrauchenia has the closest affinity to the Palæotherium.
But the Palæotherium, like the Rhinoceros and Tapir, has the ulna distinct from the radius, and the fibula from the tibia; so that even if the Parisian Pachyderm had actually presented the same peculiarities of the cervical vertebræ as the Patagonian one, it would have been hazardous, to say the least, while ignorant of the dentition of the latter, to refer it to the genus Palæotherium.
Most interesting, indeed will be the knowledge, whenever the means of obtaining it may arrive, of the structure of the skull and teeth in the Macrauchenia.
Meanwhile, we cannot but recognise, in the anchylosed and confluent state of the bones of the fore-arm and leg, a marked tendency in it towards the Ruminant Order, and the singular modifications of the cervical vertebræ have enabled us to point out the precise family of that order, with which the Macrauchenia is more immediately allied.
In first demonstrating this relationship, it was shown in how many particulars the Camelidæ, without losing the essential characters of Ruminantia, manifested a tendency to the Pachydermatous type; and the evidence which the lost genera, Macrauchenia and Anoplotherium, bear to a reciprocal transition from the Pachyderms to the Ruminants, through the Camelidæ, cannot but be viewed with extreme interest by the Zoologist engaged in the study of the natural affinities of the Animal Kingdom.
The Macrauchenia is not less valuable to the Geologist, in reference to the geographical distribution of animal forms. It is well known how unlooked-for and unlikely was the announcement of the existence of an extinct quadruped entombed in the Paris Basin, whose closest affinities were to a genus, (Tapirus,) at that time, regarded as exclusively South American. Still greater surprise was excited when a species of the genus Didelphys was discovered to have co-existed in Europe with the Palæotherium.
Now, on the other hand, we find in South America, besides the Tapir, which is closely allied to the Palæothere, — and the Llama, to which the Anoplothere offers many traces of affinity, —the remains of an extinct Pachyderm, nearly akin to the European genus Palæotherium : and, lastly, this Macrauchenia is itself in a remarkable degree a transitional form, and manifests characters which connect it both with the Tapir and the Llama.
ADMEASUREMENTS OF THE BONES OF THE
MACRAUCHENIA.
| Inches. | Lines. | |
| Length of third (?) cervical vertebra . . . . . . . | 7 | 9 |
| Vertical diameter of ditto . . . . . . . . . | 4 | 0 |
| Do. do. of body of ditto . . . . . . . . | 2 |
3 |
| Transverse
diameter of ditto . . . . . . . . . |
3 |
3 |
| Vertical diameter of spinal canal . . . . . . . . | 1 |
... |
| Length of fourth lumbar vertebra . . . . . . . . | 5 |
5 |
| Vertical diameter of body of ditto . . . . . . . . | 2 |
9 |
| Transverse
diameter of ditto . . . . . . . . . |
2 |
10
|
| Vertical diameter of spinal canal . . . . . . . . | 1 |
1 |
| Transverse ditto ditto * . . . . . . . . | 1 |
6 |
* This diameter increases rapidly in the posterior lumbar vertebræ, in correspondence with the enlargement of the spinal chord, which gives off the great nerves of the hinder extremities.
[page] 56 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
| Inches. | Lines. | |
| Transverse diameter of last lumbar vertebra . . . . . . | 9 |
... |
| Ditto do. of body of ditto . . . . . . . | 2 |
2 |
| Vertical diameter of ditto . . . . . . . . . | 1 |
3 |
| Entire length of lumbar region of vertebral column . . . . . | 20 |
|
| Vertical diameter of glenoid cavity of scapula . . . . . . | 3 |
... |
| Transverse ditto ditto ditto . . . . . . | 2 |
10
|
| Elevation
of spine of
scapula . . . . . . . . . |
3 |
5 |
| Vertical diameter of proximal articular surface of fore-arm . . . . | 3 |
6 |
| Transverse ditto ditto ditto . . . . . . | 3 |
5 |
| Height of olecranon . . . . . . . . , . | 5 |
3 |
| Greatest diameter of its base . . . . . . . . . | 2 |
... |
| Circumference of proximal end of anchylosed radius and ulna . . . | 11 |
10 |
| Entire
length of inner toe of fore-foot, inclusive of metacarpal bone
. . |
13 |
... |
| Breadth of proximal end of metacarpus . . . . . . . | 3 |
8 |
| Do. distal end of ditto . . . . . . . . | 5 |
4 |
| Length of inner metacarpal bone . . . . . . . . | 7 |
6 |
| Do. middle ditto . . . . . . . . . . | 8 |
... |
| Do. outer ditto . . . . . . . . . . | 7 |
... |
| Do.
inner proximal
phalanx . . . . . . . . . |
3 |
6 |
| Do. middle ditto . . . . . . . . . . | 2 |
10 |
| Do. outer ditto . . . . . . . . . . | 3 |
4 |
| Do.
inner middle
phalanx
. . . . . . . . . |
2 |
... |
| Do. middle ditto . . . . . . . . . . | 2 |
3 |
| Do. inner distal phalanx * . . . . . . . . | 1 |
... |
| Do. the femur . . . . . . . . . . | 24 |
... |
| Diameter of base of articular surface of the head of ditto . . . . | 3 |
6 |
| Greatest diameter of proximal end . . . . . . . | 7 |
... |
| Do. of distal end . . . . . . . . | 6 |
3 |
| Circumference of middle of shaft . . . . . . . . | 8 |
... |
| Length of tibia . . . . . . . . . . . | 18 |
|
| Greatest diameter of proximal end . . . . . . . . | 5 |
7 |
| Do. of distal end, including fibula . . . . . | 4 |
4 |
| Circumference of middle of shaft . . . . . . . . | 9 |
... |
| Length
of metatarsal bone† . . . . . . . . . |
7 |
4 |
* The relative breadth of these bones is shown in the figures of the fore-foot, Pl. XI.
† The figures in Pl. XIV. preclude the necessity of giving the admeasurements of the astragalus.
ERRATA.—The reader is requested to substitute the word 'right' for 'left' in the last line of p. 35, before the words 'radius,' 'fore-foot,' and 'femur,' and in the first line of p, 36, before the words 'tibia,' and 'hind-foot.'
DESCRIPTION OF A FRAGMENT OF A CRANIUM OF
AN EXTINCT
MAMMAL,
INDICATIVE OF A NEW GENUS OF EDENTATA, AND FOR WHICH IS PROPOSED
THE NAME OF
GLOSSOTHERIUM.
" LA première chose à faire dans l'étude d'un animal fossile, est de reconnaitre la forme de ses dents molaires; on détermine par-là s'il est carnivore ou herbivore;" says Cuvier, at the commencement of that series of splendid chapters in which the restoration of the extinct Pachyderms of the Paris Basin is recorded. In the present case, however, as in that of the Mammiferous animal whose fossil remains we were last considering, the important organs, to which Cuvier directs our first attention, are wanting. Nor are there here, as in the Macrauchenia, any remains of the locomotive extremities to compensate for the deficiency of teeth, and guide us into the right track of investigation and comparison. The animal, the nature and affinities of which are the subject of the following pages, is, in fact, represented in Mr. Darwin's collection, by nothing more than a fragment of the cranium.
This fragment, which was found in the bed of the same river, (see p. 16,) in Banda Oriental, with the cranium of the Toxodon, includes the parietes of the left side of the cerebral cavity, the corresponding nervous and vascular foramina, the left occipital condyle, a portion of the left zygomatic process, and, fortunately also, the left articular surface for the lower jaw. The importance of this surface in the determination of the affinities of a fossil animal has been duly appreciated, since the relations of the motions of the lower jaw to the kind of life of each animal were pointed out by Cuvier; but yet we should be deceived were we to establish, in conformity with the generalization enunciated by Cuvier,* our conclusion, from this surface, of the nature of the food of the extinct species under con-
* "Comme le genre de vie de chaque animal est toujours en rapport avec les mouvements dont sa mâchoire est susceptible, on retrouve dans la conformation des surfaces destinées à l'articulation, les particularités qui semblent le déterminer d'avance. Ainsi dans les animaux qui vivent de chairs, substances filamenteuses qui ne peuvent être écrasées, mais seulement coupées et dechirées, le mouvement de la mâchoire inférieure ne peut s'exécuter que de haut én bas. Dans les herbivores, les frugivores et les granivores, comme le principal mouvement est celui de broiement pour écraser, comprimer les herbes et les fruits, pour briser les grains et les réduire, en pâte, le mouvement des mâchoires se fait encore de droite à gauche, et réciproquement, on en même temps, de devant en arrière, en un mot, dans un plan horizontal autant que dans un vertical: les uns représentent des ciseaux, les autres des meules de moulin."
I
[page] 58 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
sideration; for the glenoid cavity is so shaped as to allow the lower jaw free motion in a horizontal plane, from right to left, and forwards or backwards, like the movements of a mill-stone; and, nevertheless, I venture to affirm it to be most probable, that the food of Glossotherium was derived from the animal and not from the vegetable kingdom; and to predict, that when the bones of the extremities shall be discovered, they will prove the Glossothere to be not an ungulate but an unguiculate quadruped, with a fore-foot endowed with the movements of pronation and supination, and armed with claws, adapted to make a breach in the strong walls of the habitations of those insect-societies, upon which there is good evidence in other parts of the present cranial fragment, that the animal, though as large as an ox, was adapted to prey.
We perceive, in the first place, looking upon the base of this portion of skull, a remarkable cavity, situated immediately behind the tympanic bone, of nearly a regular hemispherical form, an inch in diameter (fig. 2, b, Pl. XVI). The superficies of this cavity appears not to have been covered with articular cartilage, for it is irregularly pitted with many deep impressions; and I conclude, therefore, that it served to afford a ligamentous attachment to the styloid element of a large os hyoides. With this indication of the size of the skeleton of the tongue, is combined a more certain proof of the extent of its soft, and especially its muscular parts, in the magnitude of the foramen, for the passage of the lingual or motor nerve (c. fig. 2 and 3). This foramen, (the anterior condyloid,) in the present specimen, is the largest of those which perforate the walls of the cranium, with the exception of the foramen magnum; it is fully twice the size of that which gives passage to the second division of the fifth nerve; its area is oval, and eight lines in the long diameter, so that it readily admits the passage of the little finger.
It is only in the Ant-eaters and Pangolins that we find an approximation to these proportions of the foramen for the passage of the muscular nerve of the tongue; and the existing Myrmecophagous species even fall short of the larger fossil in this respect. Some idea of the size of the lingual nerve, and of the organ it was destined to put in motion, may be formed, when it is stated that the foramen giving passage to the corresponding nerve in the Giraffe,—the largest of the Ruminants, and having the longest and most muscular tongue in that order,—is scarcely more than one-fourth the size.
With these indications of the extraordinary development of the tongue, we are naturally led, in order to carry out a closer and more detailed comparison of the fossil in question, to that group of mammalia in which the tongue plays the chief part in the acquisition of the food. The size, form, and position of the occipital condyle,—the magnitude of the occipital foramen, (which must here have somewhat exceeded three inches in the transverse diameter,)—the slope of the occipital surface of the cranium from below, upwards and forwards, at an angle of 60°
with the base of the cranial cavity—each and all attest the close affinities of the present animal to the Edentata. More decisive evidence of the same relationship will be adduced from the organization of other parts of the cranium. The glenoid articular surface (a, fig. 2, Pl. XVI.) is an almost flattened plane, wider in the transverse than in the longitudinal direction; and, as in the genera Myrmecophaga and Manis, it is not defended behind by any descending process. In its general form it resembles the glenoid cavity of Orycteropus more than that of the preceding Edentates; but, in Orycteropus, the articulation is defended posteriorly by a descending process of the zygoma, and it is also situated relatively closer to the os tympanicum.
Had the Glossotherium teeth ? The extent of the temporal muscle, which is indicated by the rugged surface of the temporal fossa, and by the well-marked boundary, formed by a slightly elevated bony ridge, which extends to near the line of the sagittal suture, together with the size of the zygomatic portion of the temporal bone, and the remains of the oblique suture by which it was articulated to the malar bone, enables me to answer this question confidently in the affirmative. They will probably be found to be molar teeth of a simple structure, as in the Orycteropus.
The evidence just alluded to of the existence of an os malæ is interesting, because this bone is wanting in the Pangolins; and its rudimental representative in the true Ant-eaters does not reach the zygomatic process of the temporal bone, which consequently has no articular or sutural surface at its anterior extremity. In the presence, therefore, of the surface for the junction of the os malæ, and the consequent evidence of the completion of the zygomatic arch, we learn that the Glossothere was more nearly allied to the Armadillos and Orycterope. That its affinity to the latter genus was closer than to the Armadillos we have most interesting evidence in the form and loose condition of the tympanic bone: it is represented of the natural size at fig. 4, Pl. XVI. Through the care and attention devoted to his specimens by their gifted discoverer, this bone was preserved in situ, as represented at d, fig. 1; but it had no osseous connection with the petrous or other elements of the temporal bone, and could be displaced and replaced with the same ease as in the Orycterope. This bony frame of the membrana tympani, in the Glossothere, describes rather more than a semicircle, having the horns directed upwards; it has a groove, one line in breadth, along its concave margin, for the attachment of the ear-drum, and sends down a rugged process, half an inch long, from its lower margin. In the Dasypodes and Myrmecophagæ, the tympanic bone soon becomes anchylosed with the other parts of the temporal; it is only in Orycteropus, among the existing insectivorous Bruta or Edentata, that it manifests throughout life the fœtal condition of a distinct bony hoop, deficient at the upper part. The os tympanicum of Orycteropus, however, differs from that of
[page] 60 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
Glossotherium, in forming part of the circumference of an ellipse, whose long axis is vertical; and in sending outwards, from its anterior part, a convex eminence, which terminates in a point directed downwards and forwards.
Such appear to be the most characteristic features of the cranial fragment under consideration, in which we have found, that the articular surface for the os hyoides throws more light upon the nature of the animal of which it is a part, than even the glenoid cavity itself. There now remains to be described as much of the individual characters of the constituent bones as the specimen exhibits.
The occipital bone, besides forming the posterior and part of the inferior parietes of the cranium, extends for about half an inch upon the sides, where the ex-occipital element is articulated by a vertical suture with the mastoid element of the temporal: this suture is situated in a deep and well-marked muscular depression (e, fig. 1), measuring three inches in the vertical, and upwards of one inch in the transverse direction. The other sutures, uniting the occipital to the adjoining bones, are obliterated. The breadth of the occipital region must have exceeded the height of the same by about one-third. The condyle extends nearly to the external boundary of the occipital aspect of the cranium; there is situated, external to it, only a small ovate, rounded and smooth protuberance. The slightly concave surface of the occipital plane of the cranium is bounded above by a thick obtuse ridge, the muscular impressions are well sculptured upon it. It is traversed transversely at its upper third by a slightly elevated bony crest; and the surface below this ridge is again divided by a narrower intermuscular crest, which runs nearly vertically, at about an inch and a half from the external boundary of the occipital plane. As a similar crest must have existed on the opposite side, the general character of the occipital surface in the Glossothere would resemble that of the Toxodon. A similar correspondence may be noticed in the terminal position of the condyle, and the slope of the occipital plane.
Above the transverse ridge, the rough surface of the occipital plane slopes forward, at a less obtuse angle with the basal plane, to the first named ridge which separates the occipital from the coronal or superior surface of the skull. The contour of this surface runs forwards, as far as the fragment extends, in an almost straight line : the extent of surface between the temporal muscular ridges must have been about five inches posteriorly, but it decreases gradually as it extends forwards: all that part which is preserved is quite smooth. The attachment of the fasciculi of the temporal muscle, and the convergence of their fibres as they passed through the zygoma are well marked on the sculptured surface of the bone. The zygomatic process is relatively stouter than in Orycteropus : it is prismatic : the external facet is nearly plane: the superior is concave, and increases in breadth anteriorly: the inferior surface offers a slight convexity behind the flattened articular surface for the lower jaw. The margin
of the zygoma formed by the meeting of the upper and lower facets presents a semicircular curve, extended transversely from the cranium, and directed forwards.
The anterior extremity is obliquely truncated from below upwards and forwards, and presents a flattened triangular surface indicative of its junction with an os malæ: the space between this extremity and the side of the cranium measures one inch and nine lines across, and thus gives us the thickness of the temporal muscle. The distance from the origin of the zygoma to the occipital plane is relatively greater than in Orycteropus; Glossotherium is in this respect more similar to Myrmecophaga and Manis.
The sphenoid bone forms a somewhat smooth protuberance below and behind the base of the zygoma. The tympanic bone is wedged in between this protuberance in front, and the mastoid process behind. The chief peculiarity of the broad mastoid is the regular semicircular cavity at its under part for the articulation of the styloid bone of the tongue. This depression is separated below by a broad rough protuberance from the foramen jugulare, (f, fig. 2, Pl. XVI,) which is immediately external to, and slightly in advance of the great foramen condyloideum, c. A small rugged portion of the os petrosum separates the jugular from the carotid canal, which arches upwards and directly inwards to the side of the shallow sella turcica, (the external and internal orifices of the carotid canal are shown at g, figs. 2 and 3). The chief protuberance on the basis cranii is a large and rugged one, serving for the attachment of muscles, and due chiefly to the expansion of a great sinus in the body of the sphenoid. This protuberance is separated from the smaller sphenoid protuberance before mentioned by a large groove continued downwards and forwards from the tympanic cavity, and containing the Eustachian tube, which does not traverse a complete osseous canal. Immediately internal to the glenoid cavity is the large orifice of the canal transmitting the third division of the fifth pair of nerves, the principal branch of which endows the tongue with sensibility; this foramen (h, fig. 2) is rather less than that for the muscular nerve of the tongue.
The internal surface of the present cranial fragment affords a very satisfactory idea of the size and shape of the brain of the extinct species to which it belongs. It is evident that, as in other Bruta, the cerebellum must have been almost entirely exposed behind the cerebrum; and that the latter was of small relative size, not exceeding that of the Ass; and chiefly remarkable, as in the Orycterope, Ant-eater, and Armadillo for the great development of the olfactory ganglia. The antero-posterior extent of the cribriform plate, as exposed in this fragment, is three inches, and the complication of the œthmoid olfactory lamellæ which radiate from it into the nasal cavity is equal to that which exists in the smaller Edentata (fig. 3, Pl. XVI). The nasal cavity is complicated in Glosso-
[page] 62 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
therium by the great number and capacious size of the air-cells which are in communication with it: these extend over all the upper, lateral, and back parts of the cranial cavity, as far even as the upper boundary of the foramen magnum: they also occupy the anterior two-thirds of the basis cranii. The external configuration of the skull would, therefore, afford a very inadequate or rather deceptive notion of the capacity of the cerebral cavity, were not the existence and magnitude of these sinuses known. The interspace of the outer and inner tables of the cranium are separated above the origins of the olfactory ganglia for the extent of three inches: above the middle of the cerebrum they are an inch and a half apart; at the sides of the cranium the interposed air-cells are from one to two inches across; at the back part of the cranium about one inch. The sinuses have generally a rounded form.
The foramen rotundum, (through which in figure 3 a probe is represented as passing), and the foramen ovale are situated close together, within a common transversely oblong depression (i). The carotid canal (g) opens into the outer side of the commencement of this wide channel, which conducts the great fifth pair of nerves to the outlets of its two chief divisions.
The petrous bone projects into the cranial cavity, in the form of an angular process with three facets: the foramen auditorium internum (k), and the aqueductus vestibuli, are situated on the posterior facet. Immediately behind the os petrosum is the foramen lacerum jugulare (l), situated at the point of convergence of the vertical groove of the lateral sinus, with a groove of similar size continued forwards from above the anterior condyloid canal. The plane of the internal opening of this canal (c, fig. 3) is directed obliquely inwards and backwards, and the lateral wall of the foramen magnum behind the foramen condyloideum slopes outwards to the edge of the condyle. Immediately internal to the foramen condyloideum is a small vascular foramen conducting a branch of the basilar artery into the condyloid canal, for the nourishment, doubtless, of the great lingual nerve.
In the relations of the plane of the internal orifice of the anterior condyloid foramen with that of the foramen magnum, we search in vain for a corresponding structure in any of the Mammiferous orders, save the Edentata :* and among these the Orycterope comes nearest the Glossothere in this respect. In the degree of development of the internal osseous ridge giving attachment to the tentorium cerebelli, the Ant-eaters and Armadillos more resemble the Glossothere than does the Orycterope; in which a continuous bony plate arches across the cranial cavity: in the Manis a still greater proportion of the tentorium is ossified,
* In the monotrematous Echidna, the large canal for the lingual nerve has a widely different direction and course from that in the placental Edentata.
and it consequently recedes the furthest amongst the Edentata, in this, as in most other particulars of the cranial organization, from the Glossothere. The chief distinctive peculiarity in the cranium of the Glossothere, so far as it can be studied in the present fragment, and compared with that of other Edentata, is the deep, well-marked, semicircular styloid depression, above described.
A question may arise after perusing the preceding evidence, upon which the present fossil is referred to a great Edentate species nearly allied to the Orycteropus, whether one or other of the lower jaws, subsequently to be described, and in like manner referable, from their dentition, either to the Orycteropodoid or Dasypodoid families of Edentata, may not have belonged to the same species as does the present mutilated cranium. I can only answer, that those jaws were discovered by Mr. Darwin in a different and very remote locality,—that no fragments or teeth referable to them were found associated with the present fossil; and that, as it would be, therefore, impossible to determine from the evidence we have now before us, which of the two lower jaws should be associated with Glossotherium ; and as both may with equal if not greater probability belong to a totally distinct genus, it appears to me to be preferable, both in regard to the advancement of our knowledge of these most interesting Edentata of an ancient world, as well as for the convenience of their description, to assign to them, for the present, distinct generic appellations.
The figures in Plate XVI. preclude the necessity of a table of admeasurements of the cranial fragment of Glossotherium.
~~~~~~~~~~~~~~~~
DESCRIPTION OF A MUTILATED LOWER JAW AND
TEETH, ON WHICH
IS FOUNDED
A SUBGENUS OF MEGATHERIOID EDENTATA, UNDER THE NAME OF
MYLODON.
The genus Megalonyx, as is well known, owes its name and the discovery of the fossil remains on which it was founded, to the celebrated Jefferson,* formerly President of the United States. Cuvier, from an examination of a single tooth, and the casts of certain bones of the extremities, especially the terminal ones, determined the ordinal affinities of this remarkable extinct quadruped.† But while he
* Transactions of the Philosophical Society of Philadelphia, vol. iv. p. 246.
† Its relations to the Edentata, previously conjectured by Dr. Wistar, are proved in the Annales du Muséum, tom. v. p. 358; its more immediate affinities as an annectant form in that group are discussed in the edition of the Ossem. Fossiles, of 1833, tom. v. pt. 1. p. 160.
[page] 64 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
retained the name of Megalonyx, and used it in a generic sense, Cuvier offered no characters whereby other fossil remains might be generically either distinguished from, or identified with the Megalonyx Jeffersonii, unless, among such remains there happened to be a tooth, or a claw exactly corresponding with the descriptions and figures in the Ossemens Fossiles; and when, of course, a specific identity, and not merely a generic relationship would be established.
The greater part of Cuvier's chapter on Megalonyx is devoted to the beautiful and justly celebrated reasoning on the ungueal phalanx, whereby it is proved to belong, not to a gigantic Carnivore of the Lion-kind, as Jefferson supposed, but to the less formidable order of Edentate quadrupeds; and Cuvier, in reference to the tooth,—the part on which alone a generic character could have been founded,—merely observes that it resembles at least as much the teeth of one of the great Armadillos, as it does those of the Sloths.*
In the last edition of the Rêgne Animal, Cuvier introduces the Megatherium and Megalonyx, between the Sloths and Armadillos; but alludes to no other difference between the two genera than that of size,—"l'autre, le Megalonyx, est un peu moindre." (p. 226.) Some systematic naturalists, as Desmarest, and Fischer, have, therefore, suppressed the genus, and made the Megalonyx a species of Megatherium under the name of Megatherium Jeffersonii. The dental characters of the genus Megatherium are laid down by Fischer† as follows: —"Dent. prim. et lan. 0/0. molares 4/4-4/4, obducti, tritores, coronide nunc planâ transversim sulcatâ nunc medio excavatâ marginibus prominulis." That Megalonyx had the same number of molares as Megatherium, (supposing that number in the Megathere to be correctly stated, which it is not,) is here assumed from analogy, for neither Jefferson, Wistar, nor Cuvier, — the authorities for Megalonyx quoted by Fischer — possessed other means of knowing the dentition of that animal than were afforded by the fragment of a single tooth.
Now the almost entire lower jaw about to be described offers, in so far as respects the general form and structure of the teeth, the same kind and degree of correspondence with the Megatherium, as does the Megalonyx Jeffersonii of Cuvier: and, what is only probable in that species, is here certain, viz., an agreement with the Megatherium in the class, viz. molares, to which the teeth exclusively belong. The question, therefore, on which I find myself, in the outset, called upon to come to a decision is, as to the preference of the mode of viewing the subject of the generic relationship of the Megalonyx adopted by Desmarest, Fischer, &c., or of that, on which Cuvier, and after him Dr. Harlan, have practically acted: whether, in short, the genus Megatherium is to rest upon the more
* Speaking of this tooth, Cuvier observes, "Je l'avois cru d'abord nécessairement de paresseux; mais aujourd-hui que je connois mieux l'ostéologie des divers tatous, je trouve qu'elle ressemble au moins autant à une dent de l'un des grands tatous.—Loc. cit. p. 172.
† Synopsis Mammalium.
Pl. VIII.

G. Scharf del et lithog.
Lumbar Vertebræ; Macrauchenia.
Fig. 1. Posterior View of last lumbar. Fig: 2. 3 & 4. Fourth lumbar Vertebra.
Nat: Size.
Published by Smith, Elder & Co,, 65, Cornhill London.
Pl. IX.
Lithog from Nature by G. Scharf.
Macrauchenia.
Fig. 1-2. Scapula Fig. 3. Femur.
Published by Smith, Elder & Co,, 65, Cornhill.
Pl. X.

G. Scharf del et lithog.
Proximal Extremity of anchylosed Ulna and Radius, Macrauchenia.
2/3 Nat. Size.
London Published by Smith, Elder & Co,, 65, Cornhill.
Pl. XI.

G. Scharf del et lithog.
Bones of the right fore-foot, Macrauchenia.
Fig 1, 2/3. 2-9., Nat. Size.
Published by Smith, Elder & Co,, 65, Cornhill.
Pl. XII.
Lithog from Nature by G. Scharf.
Right Femur. Macrauchenia.
Published by Smith, Elder & Co,, 65, Cornhill.
Pl. XIII.
Lithog from Nat. by G. Scharf.
Macrauchenia.
Right Tibia and Fibula. - Fig. 2-4 2/3 Nat. Size.
Published by Smith, Elder & Co,, 65, Cornhill.
Pl. XIV.

Lithog from Nature by G. Scharf.
Right Astragulas. Macrauchenia.
Nat. Size.
Published by Smith, Elder & Co,, 65, Cornhill.
Pl. XV.

Lithog from Nature by G. Scharf.
Macrauchenia.
Fig: 1 Metatarsal. 2-5, Matacarpals. Nat. Size.
Published by Smith, Elder & Co,, 65, Cornhill.
Pl. XVI.

Lithog from Nature by G. Scharf.
Fragment of the Cranium of the Glossotherium.
1/2 Nat. Size.
Published by Smith, Elder & Co,, 65, Cornhill.
Pl. XVII.
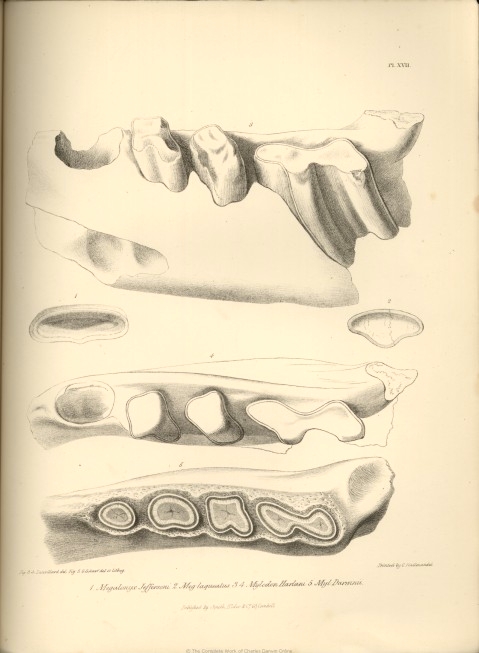
Fig. 3,4 Laurillard del. Fig 5 G. Scharf del et lithog.
1. Megalonyx Jeffersoni. 2. Meg laqueatus. 3,4 Mylodon Harlani. 5. Myl Darwinii.
1/2 Nat. Size.
Published by Smith, Elder & Co,, 65, Cornhill.
Now Publishing in Parts, Royal 4to.
UNDER THEY AUTHORITY OF THE LORDS COMMISSIONERS OF HER MAJESTY'S TREASURY
ILLUSTRATIONS OF
THE ZOOLOGY OF SOUTH AFRICA:
Consisting chiefly of Figures and Descriptions of the Objects of Natural History collected during an Expedition
into the Interior of South Africa, in the years 1834, 1835, and 1836, fitted out by
"The Cape of Good Hope Association for Exploring Central Africa."
TOGETHER WITH
A SUMMARY OF AFRICAN ZOOLOGY,
AND AN INQUIRY INTO THE GEOGRAPHICAL RANGES OF SPECIES IN THAT QUARTER OF THE GLOBS.
BY ANDREW SMITH, M.D.
SURGEON TO THE FORCES, AND DIRECTOR OP THE EXPEDITION.
PROSPECTUS
The Cape of Good Hope is now acknowledged to be one of the greatest avenues as yet opened for the researches of the Naturalist. Our Colony in that part of Southern Africa is the key to a large portion of an extensive continent which is still but very partially explored; and the field to which it admits the scientific traveller is rich to exuberance in the variety and novelty, both of animal and vegetable life. Stimulated by the prospect of Discovery in a quarter so fertile in interest, "The Cape of Good Hope Association for Exploring Central Africa" was established in 1833; and in 1836, an Expedition fitted out by that body, consisting of thirty-four persons, and directed by Dr. Smith, after an absence of nineteen months, and penetrating as far as 23° 28' South latitude, returned to Cape Town laden with a variety of curious and important specimens in Natural History, &c.
Previously to this period little information has been furnished, in a shape calculated to enable the public to form accurate ideas of the various animated beings by which these regions are inhabited. The splendid publication of Le Vaillant, no doubt, should be mentioned as forming an exception, pro tanto; but this includes only a portion of the Birds of the most southern extremity of the country, and a work therefore extensive enough to comprehend the various departments of Zoology is still a desideratum.
The Members of The Cape of Good Hope Association for Exploring Central Africa found themselves, on the return of the recent Expedition, in a situation to supply at least some portion of the existing deficiencies; but their funds, even if it had been possible to divert them to such an object, were altogether inadequate to defray the expense of laying the result of their labours before the world. Under such circumstances, it was decided that Dr. Smith, the director of the Expedition, should be authorised, on his arrival in England, to wait upon Lord Glehelg, for the purpose of making him acquainted with the position and views of the Society, in the hope that Government might be induced to assist in the publication of their materials.
This hope has not been disappointed. At the recommendation of the Secretary of State for the Colonial Department, the Lords Commissioners of Her Majesty's Treasury have been pleased, by a pecuniary grant, to enable the Society to publish the result of its labours, without infringing upon the funds raised solely for the purposes of discovery; and in a form which, while it places the work within reach of most of the friends and promoters of science, will not, it is hoped, be found inconsistent with the interest and importance of the subject.
The materials for the work now offered, under such patronage, to the public, will consist of pictorial illustrations of between three and four hundred subjects of the animal kingdom, all of which have been collected to the south of 23° 28' South latitude; and will comprise,
First, and principally, unknown animals;
Secondly, animals known, but not yet figured; and
Lastly, such as have been imperfectly figured; but of which the Association is in possession of accurate drawings.
The Entomological portion of the work will be from the pen of W. S. Macleay, Esq., who has kindly undertaken that department. The rest of the descriptions will be furnished by Dr. Smith, who will add a summary of African Zoology, and an inquiry into the Geographical ranges of species in that quarter of the Globe.
Conditions of Publication.
The Work will appear periodically; and it is estimated that the whole will be completed in about thirty-four parts, price, on an average, Ten Shillings each. As it will be necessary that the plates be published promiscuously, they will be arranged in five divisions, viz. Mammalia, Aves, Pisces, Reptilia, and Invertebrate. The plates of each of these divisions will be numbered independently, and the letter-press descriptions left unpaged, so that on the work being completed, they may be arranged either agreeably to the general classified order which will accompany the last number, or according to the particular views of the purchasers.
The whole of the Plates will be engraved in the highest style of Art, from Drawings taken expressly for this Work,
and beautifully coloured after Nature.
THE FIRST FIVE PARTS ARE NOW READY.
[Continued in Fossil Mammalia Part 1 No. 3 ]