Fertilisation of
Orchids
Darwin
LONDON
JOHN MURRAY
ON
THE VARIOUS CONTRIVANCES
BY WHICH
BRITISH AND FOREIGN ORCHIDS
ARE
FERTILISED BY INSECTS,
AND ON THE GOOD EFFECTS OF INTERCROSSING.
BY CHARLES DARWIN, M.A., F.R.S., &c.
WITH ILLUSTRATIONS.
LONDON:
JOHN MURRAY, ALBEMARLE STREET.
1862.
The right of Translation is reserved.
BY THE SAME AUTHOR.
THE ORIGIN OF SPECIES BY MEANS OF NATURAL SELECTION; or, the Preservation of Favoured Races in the Struggle for Life. Seventh Thousand. MURRAY.
A NATURALIST'S VOYAGE ROUND THE WORLD; or, a Journal of Researches into the Natural History and Geology of the Countries visited during the Voyage of H.M.S. Beagle, under the command of Capt. Fitz-Roy, R.N. Tenth Thousand. MURRAY.
ON THE STRUCTURE AND DISTRIBUTION OF CORAL REEFS. SMITH, ELDER, & Co.
GEOLOGICAL OBSERVATIONS ON VOLCANIC ISLANDS. SMITH, ELDER, & Co.
GEOLOGICAL OBSERVATIONS ON SOUTH AMERICA. SMITH, ELDER, & Co.
A MONOGRAPH OF THE CIRRIPEDIA; with numerous Illustrations. 2 vols. 8vo.HARDWICKE.
PREPARING FOR PUBLICATION.
ON THE VARIATION OP ANIMALS AND PLANTS UNDER DOMESTICATION.
_____________________________
LONDON: PRINTED BY W. CLOWES AND SONS, STAMFORD STREET, AND CHARING CROSS.
CONTENTS.
—°—
INTRODUCTION .. .. .. .. .. .. .. .. .. .. .. Page 1
CHAPTER I.
Structure of Orchis — Power of movement of the pollinia — Perfect adaptation of the parts in Orchis pyramidalis — On the insects which visit Orchids, and on the frequency of their visits — On the fertility and sterility of several Orchids — On the secretion of nectar, and on moths being purposely delayed in obtaining it .. .. .. .. .. .. .. .. .. 9-53
CHAPTER II.
Ophreæ continued — Fly and Spider Ophrys — Bee Ophrys, apparently adapted for perpetual self-fertilisation, but with paradoxical contrivances for intercrossing — The Frog Orchis; fertilisation effected by nectar secreted from two parts of the labellum — Gymnadenia conopsea — Greater and Lesser Butterfly Orchis; their differences and means of fertilisation — Summary on the powers of movement in the pollinia .. .. .. .. .. .. .. .. .. 54-92
CHAPTER III.
Epipactis palustris; curious shape of the labellum and its apparent importance in the fructification of the flower — Cephalanthera grandiflora; rostellum aborted; early penetration of the pollen-tubes; case of imperfect self-fertilisation; fertilisation aided by insects — Goodyera repens — Spiranthes autumnalis; perfect adaptation by which the pollen of a younger flower is carried to the stigma of an older flower on another plant .. .. .. .. .. .. .. .. .. 93-129
CHAPTER IV.
Malaxis paludosa; simple means of fertilisation — Listera ovata; sensitiveness of the rostellum; explosion of viscid matter; action of insects; perfect adaptation of the several oegans — Listera cordata — Neottia nidus-avis; its fertilisation effected in the same manner as in Listera .. .. 130-156
CHAPTER V.
Cattleya, simple manner of fertilisation — Masdevallia, curious closed flower — Dendrobium, contrivance for self-fertilisation — Vandeæ, diversified structure of the pollinia; importance of the elasticity of the pedicel; its power of movement — Elasticity and strength of the caudicle — Calanthe with lateral stigmas, manner of fertilisation — Angræcum sesquipedale, wonderful length of nectary — Acropera, perplexing case, a male Orchid .. .. .. .. .. .. .. .. ..Page 157-210
CHAPTER VI.
Catasetidæ, the most remarkable of all Orchids — The mechanism by which the pollinia of Catasetum are ejected to a distance, and are transported by insects — Sensitiveness of the horns of the rostellum — Extraordinary difference in the male, female, and hermaphrodite forms of Catasetum tridentatum — Mormodes ignea, curious structure of flower; ejection of its pollinia — Cypripedium, importance of the slipper-like form of the labellum — Secretion of nectar — Advantage derived from insects being delayed in sucking the nectar — Singular excrescences on the labellum, apparently attractive to insects .. .. .. .. .. .. 211-285
CHAPTER VII.
Homologies of Orchid-flowers — The great modification which they have undergone — Gradation of organs, of the rostellum, of the pollen-masses — Formation of the caudicle — Genealogical affinities — Mechanism of the movement of the pollinia — Uses of the petals — Production of seed — Importance of trifling details of structure — Cause of the vast diversity of structure for the same general purpose — Cause of the perfection of the contrivances in Orchids — Summary on insect-agency — Nature abhors perpetual self-fertilisation .. .. .. .. .. .. 286-360
INDEX .. .. .. .. .. .. 361-365
LIST OF WOODCUTS.
—°—
PAGE. |
||
I. ORCHIS MASCULA to face |
18 |
|
II. " " POLLINIA OF |
15 |
|
III. ORCHIS PYRAMIDALIS |
22 |
|
37 |
||
55 |
||
VI. " ARANIFERA |
61 |
|
VII. " APIFERA |
63 |
|
VIII. " ARACHNITES |
72 |
|
76 |
||
80 |
||
84 |
||
90 |
||
XIII. EPIPACTIS PALUSTRIS |
94 |
|
XIV. " LATIFOLIA |
103 |
|
105 |
||
117 |
||
XVII. MALAXIS PALUDOSA |
132 |
|
XVIII. LISTERA OVATA |
141 |
|
XIX. CATTLEYA |
161 |
|
168 |
||
173 |
||
179 |
||
XXIII. POLLINIA OF VANDEAE |
185 |
|
XXIV. CALANTHE MASUCA |
195 |
|
XXV. XXVI. |
216 217 |
|
|
|
PAGE. |
XXVII. CATASETUM TRIDENTATUM |
232 |
XXVIII. MONACHANTHUS |
239 |
XXIX. MORMODES IGNEA |
250 |
XXX. MORMODES |
267 |
XXXI. CYPRIPEDIUM |
272 |
292 |
|
XXIII. ROSTELLUM OF CATASETUM |
322 |
XXXIV. DISC OF GYMNADENIA |
336 |
P.S.—I am much indebted to Mr. G.B. Sowerby for the pains which he has taken in making the Diagrams as intelligible as possible.
__________
INSTRUCTION TO BINDER.
Let the Woodcut No I. face page 18.
__________
ON THE
FERTILISATION OF ORCHIDS
BY INSECTS,
&c. &c.
______
______
INTRODUCTION.
THE object of the following work is to show that the contrivances by which Orchids are fertilised, are as varied and almost as perfect as any of the most beautiful adaptations in the animal kingdom; and, secondly, to show that these contrivances have for their main object the fertilisation of each flower. In my volume 'On the Origin of Species' I have given only general reasons for my belief that it is apparently a universal law of nature that organic beings require an occasional cross with another individual; or, which is almost the same thing, that no hermaphrodite fertilises itself for a perpetuity of generations. Having been blamed for propounding this doctrine without giving
B
ample facts, for which I had not, in that work, sufficient space, I wish to show that I have not spoken without having gone into details.
I have been led to publish this little treatise separately, as it has become inconveniently large to be incorporated with the rest of the discussion on the same subject. And I have thought, that, as Orchids are universally acknowledged to rank amongst the most singular and most modified forms in the vegetable kingdon, the facts to be presently given might lead some observers to look more curiously into the habits of our several native species. An examination of their many beautiful contrivances will exalt the whole vegetable kingdom in most persons' estimation. I fear, however, that the necessary details will be too minute and complex for any one who has not a strong taste for Natural History. This treatise affords me also an opportunity of attempting to show that the study of organic beings may be as interesting to an observer who is fully convinced that the structure of each is due to secondary laws, as to one who views every trifling detail of structure as the result of the direct interposition of the Creator.
I must premise that Christian Konrad Sprengel, in his curious and valuable work, 'Das entdeckte Geheimniss der Natur,' published in 1793, gave an excellent outline of the action of the several parts in Orchids; for he well knew the position of the stigma; and he discovered that insects were necessary to remove the pollen-masses, by pushing open the pouch and coming into contact with the enclosed sticky glands. But he overlooked many curious contrivances,—a consequence, apparently, of his belief that the stigma generally receives the pollen of the same flower. Sprengel, likewise, has partially described the structure of Epipactis; but in the case of Listera he entirely misunderstood the remarkable phenomena characteristic of that genus, which has been so well described by Dr. Hooker in the 'Philosophical Transactions' for 1854. Dr. Hooker has given a full and accurate account, with drawings, of the structure of the parts, and of what takes place; but from not having attended to the agency of insects, he has not fully understood the object gained. Robert Brown,* in
* 'Linnæan Transactions,' 1833, vol. xvi. p. 704.
B2
his celebrated paper in the 'Linnæan Transactions,' expresses his belief that insects are necessary for the fructification of most Orchids; but adds, that the fact of all the capsules on a dense spike not infrequently producing seed, seems hardly reconcileable with this belief: we shall hereafter see that this doubt is groundless. Many other authors have given facts and expressed their belief, more or less fully, on the necessity of insect-agency in the fertilisation of Orchids.
In the course of the following work I shall have the pleasure of expressing my deep obligation to several gentlemen for their unremitting kindness in sending me fresh specimens, without which aid this work would have been impossible. The trouble which several of my kind assistants have taken has been extraordinary: I have never once expressed a wish for aid or for information which has not been granted, as far as possible, in the most liberal spirit.
____________
EXPLANATION OF TERMS.
In case any one should look at this treatise who has never attended to Botany, it may be convenient to explain the meaning of the common terms used. In most flowers the stamens, or male organs, surround in a ring the one or more female organs, called the pistils. In all common Orchids there is only one stamen, and this is confluent with the pistil forming the Column. The stamens consist of a filament, or supporting thread (rarely seen in British Orchids), which carries the anther; and within the anther the pollen, the male vivifying element, is included. The anther is divided into two cells, which are very distinct in most Orchids, so much so as to appear in some species like two separate anthers. The pollen in all common plants consists of fine granular powder: but in most Orchids the grains cohere in masses, which are often supported by a very curious appendage, called the Caudicle; as will hereafter be more fully explained. The pollen-masses, with their caudicles and other appendages, are called the Pollinia.
There are properly in most Orchids three united pistils, or female organs. The upper part of the pistil has its anterior surface soft and viscid, which forms the stigma. The two lower stigmas are often completely confluent, so as to appear as one. The stigma in the act of fertilisation is penetrated by long tubes emitted by the pollen-grains, which carry the contents of the grains down to the ovules, or young seeds in the ovarium.
Of the three pistils, which ought to be present, the stigma of the upper one has been modified into an extraordinary organ, called the Rostellum, which in many Orchids presents no resemblance to a true stigma. The rostellum either includes or is formed of viscid matter; and in very many Orchids the pollen-masses are firmly attached to a portion of its exterior membrane, which is removed, together with the pollen-masses, by insects. This removeable portion consists in most British Orchids of a small piece of membrane, with a layer or ball of viscid matter underneath, and I shall call it the "viscid disc;" but in many exotic Orchids the portion removed is so large and so import-
ant, that one part must be called, as before, the viscid disc, and the other part the pedicel of the rostellum, to the end of which pedicel the pollen-masses are attached. Authors have called that portion of the rostellum which is removed the "gland," or the "retinaculum," from its apparent function of retaining the pollen-masses in place. The pedicel, or prolongation of the rostellum, to which in many exotic Orchids the pollen-masses are attached, seems generally to have been confounded, under the name of caudicle, with the true caudicle of the pollen-masses, though their nature and origin are totally different. The part of the rostellum which is not removed, and which includes the viscid matter, is sometimes called the "bursicula," or "fovea," or "pouch." But it will be found most convenient to avoid all these terms, and to call the whole modified stigma the rostellum—sometimes adding an adjective to define its shape; and to call that portion of the rostellum which is attached to and removed with the pollen-masses the viscid disc, together in some cases with its pedicel.
Lastly, the three outer divisions of the
flower are called Sepals, and form the calyx; but, instead of being green, as in most common flowers, they are generally coloured, like the three inner divisions or Petals of the flower. The one petal which commonly stands lowest is larger than the others, and often assumes most singular shapes; it is called the lower lip, or Labellum. It secretes nectar, in order to attract insects, and is often produced into a long spur-like nectary.
CHAPTER I.
Structure of Orchis—Power of movement of the pollinia—Perfect adaptation of the parts in Orchis pyramidalis—On the insects which visit Orchids, and on the frequency of their visits—On the fertility and sterility of several Orchids—On the secretion of nectar, and on moths being purposely delayed in obtaining it.
FOR my purpose British Orchids may be divided into three groups, and the arrangement is, for the most part, a natural one. But I leave out of consideration the British species of Cypripedium, with its two anthers, of which I know nothing. Of these three groups the first consists of the Ophreæ, which have pollinia furnished at their lower ends with a caudicle, congenitally attached to a viscid disc. The anther stands above the rostellum. The Ophreæ include most of our common Orchids.
First, for the genus Orchis. The reader may find the following details rather difficult to understand; but I can assure him, if he will have patience to make out this first case, the succeeding cases will be easily intelligible.
B3
The accompanying diagrams (Fig. I., p. 18) show the relative position of the more important organs in the flower of the Early Orchis (O. mascula). The sepals and the petals have been removed, excepting the labellum with its nectary. The nectary is shown only in the side view (n Fig. A); for its enlarged orifice is almost hidden in shade in (B) the front view. The stigma (s) is bilobed, and consists of two almost confluent stigmas; it lies under the pouch-formed (r) rostellum. The anther (a in B and A) consists of two rather widely separated cells, which are longitudinally open in front: each cell includes a pollen-mass or pollinium.
A pollinium removed out of one of the two anther-cells is represented by Fig. C; it consists of a number of wedge-formed packets of pollen-grains (see Fig. F, in which the packets are forcibly separated), united together by excessively elastic, thin threads. These threads become confluent at the lower end of each pollen-mass, and compose the (c C) straight elastic caudicle. The end of the caudicle is firmly attached to the viscid disc (d C), which consists (as may have been seen in the section, Fig. E)
of a minute oval piece of membrane, with a ball of viscid matter on its under side. Each pollinium has its separate disc; and the two balls of viscid matter lie enclosed together (Fig. D) within the rostellum.
The rostellum is a nearly spherical, somewhat pointed projection (r Figs. A and B) overhanging the two almost confluent stigmas, and must be fully described, as every detail of its structure is full of signification. A section through one of the discs and balls of viscid matter is given (Fig. E); and a front view of both viscid discs within the rostellum (Fig. D) is likewise given. This latter figure (D) probably best serves to explain the structure of the rostellum; but it must be understood that the front lip is here considerably depressed. The lowest part of the anther is united to the back of the rostellum, as may be seen in Fig. B. At an early period of growth the rostellum consists of a mass of polygonal cells, full of brownish matter, which cells soon resolve themselves into two balls of an extremely viscid semi-fluid substance, void of structure. These viscid masses are slightly elongated, almost flat on the top, and convex
below. They lie quite free within the rostellum (being surrounded by fluid), except at the back, where each viscid ball firmly adheres to a small portion or disc of the exterior membrane of the rostellum. The ends of the two caudicles are strongly attached to these two little discs of membrane.
The membrane forming the whole exterior surface of the rostellum is at first continuous; but as soon as the flower opens the slightest touch causes it to rupture transversely in a sinuous line, in front of the anther-cells and of the little crest or fold of membrane (see Fig. D) between them. This act of rupturing makes no difference in the shape of the rostellum, but converts the front part into a lip, which can easily be depressed. This lip is represented considerably depressed in Fig. D, and its edge is seen, Fig. B, in the front view. When the lip is thoroughly depressed, the two balls of viscid matter are exposed. Owing to the elasticity of the hinder part or hinge, the lip or pouch, when not pressed down, springs up and again encloses the two viscid balls.
I will not affirm that the rupturing of the exterior membrane of the rostellum never
takes place spontaneously; and no doubt the membrane is prepared for the rupture by having become very weak along defined lines; but several times I saw the act ensue from an excessively slight touch—so slight that I conclude that the action is not simply mechanical, but, for the want of a better term, may be called vital. We shall hereafter meet with other cases, in which the slightest touch or the vapour of chloroform causes the exterior membrane of the rostellum to rupture along certain defined lines.
At the same time that the rostellum becomes transversely ruptured in front, it probably (for it was impossible to ascertain this fact from the position of the parts) ruptures behind in two oval lines, thus separating and freeing from the rest of the exterior surface of the rostellum the two little discs of membrane, to which externally the two caudicles are attached, and to which internally the two balls of viscid matter adhere. The line of rupture is thus very complex, but strictly defined.
As the two anther-cells open longtudinally in front from top to bottom, even before the flower expands, as soon as the rostellum is
properly ruptured from the effects of a slight touch, its lip can be easily depressed, and, the two little discs of membrane being already separate, the two pollinia now lie absolutely free, but are still embedded in their proper places. So that the packets of pollen and the caudicles lie within the anther-cells; the discs still form part of the posterior surface of the rostellum, but are separate: and the balls of visvid matter still lie concealed within the rostellum.
Now let us see how this complex mechanism acts. Let us suppose an insect to alight on the labellum, which forms a good landing-place, and to push its head into the chamber (see side view Fig. I., A, or front view, B) at the back of which lies the stigma (s), in order to reach with its proboscis the end of the nectary; or, which does equally well to show the action, push a sharply-pointed common pencil into the nectary. Owing to the pouch-formed rostellum projecting into the gangway of the nectary, it is scarcely possible that any object can be pushed into it without the rostellum being touched. The exterior membrane of the rostellum then ruptures in the proper lines,
and the lip or pouch is most easily depressed. When this is effected, one or both of the viscid balls will almost infallible touch the intruding body. So viscid are these balls that whatever they touch they firmly stick to. Moreover the viscid matter has the peculiar chemical quality of setting, like a cement, hard and dry in a few minutes' time. As the anther-cells are open in front, when the insect withdraws its head, or when the pencil is withdrawn, one pollinium, or both, will be withdrawn, firmly cemented to the object, projecting up like horns, as shown (Fig. II.) by the upper figure.
Fig. II.
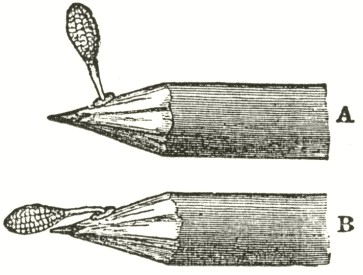
A. Pollen-mass of O. mascula, when first attached.
B. Pollen-mass of O. mascula after the act of depression.
The firmness of the attachment of the cement is very necessary, as we shall immediately see; for if the pollinia were to fall sideways or
backwards they could never fertilise the flower. From the position in which the two pollinia lie in their cells, they diverge a little when attached to any object. Now let us suppose our insect to fly to another flower, or insert the pencil (A, Fig. II.), with the attached pollinium, into the same or into another nectary: by looking at the diagram (Fig. I., A) it will be evident that the firmly attached pollinium will be simply pushed against or into its old position, namely, into its anther-cell. How then can the flower be fertilised? This is effected by a beautiful contrivance: though the viscid surface remains immoveably affixed, the apparently insignificant and minute disc of membrane to which the caudicle adheres is endowed with a remarkable power of contraction (as will hereafter be more minutely described), which causes the pollinium to sweep through about 90 degrees, always in one direction, viz., towards the apex of the proboscis or pencil, in the course, on an average, of thirty seconds. The position of the pollinium after the movement is shown in B in Fig. II. Now after this movement and interval of time (which would allow the insect
to fly to another flower), it will be seen, by turning to the diagram (Fig. I., A), that, if the pencil is inserted into the nectary, the thick end of the pollinium will exactly strike the stigmatic surface.
Here again comes into play another pretty adaptation, long ago noticed by Robert Brown.* The stigma is very viscid, but not so viscid as when touched to pull the whole pollinium off the insect's head or off the pencil, yet sufficiently viscid to break the elastic threads (Fig. I., F) by which the packets of pollen-grains are tied together, and leave some of them on the stigma. Hence a pollinium attached to an insect or to the pencil can be applied to many stigmas, and will fertilise all. I have seen the pollinia of Orchis pyramidalis adhering to the proboscis of a moth, with the stump-like caudicle alone left, all the packets of pollen having been left glued to the stigmas of the flowers successively visited.
One or two little points must still be noticed. The balls of viscid matter within the pouch-formed rostellum are surrounded with fluid; and this is very important, for,
* 'Transactions of the Linnæan Society,' vol. xvi. p. 731.
as already mentioned, the viscid matter sets hard when exposed to the air for a very short time. I have pulled the balls out of their pouches and have found that in a few minutes they entirely lost their power of adhesion. Again, the little discs of membrane, the movement of which, as causing the movement of the pollinium, is so absolutely indispensable for the fertilisation of the flower, lie at the upper and back surface of the rostellum, and are closely enfolded and thus kept damp within the bases of the anther-cells; and this is very necessary, as an exposure of about thirty seconds causes the movement of depression to take place; but as long as the disc is kept damp the pollinium remains ready for action whenever removed by an insect.
Lastly, as I have shown, the pouch, after being depressed, springs up to its former position; and this is of great service; for if this action did not take place, and an insect after depressing the lip failed to remove either viscid ball, or if it removed one alone, in the first case both, and in the second case one of the viscid balls would be left exposed to the air; consequently they would quickly lose all adhesive-
[To face page 18.
Fig. I.
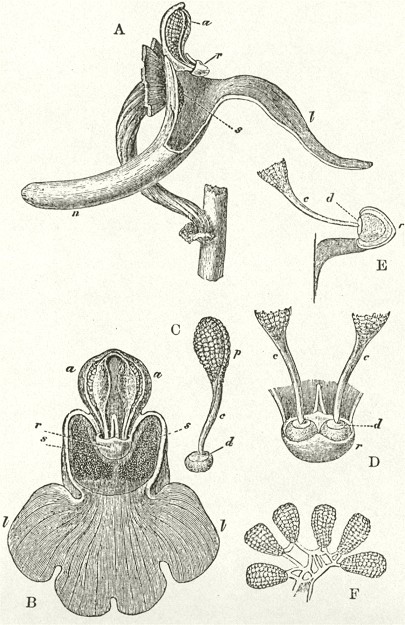
ORCHIS MASCULA.
Description of Fig. I.
a. anther.
r. rostellum.
s. stigma.
l. labellum.
n. nectary.
p. pollinium or pollen-mass.
c. caudicle of pollinium.
d. viscid disc of pollinium.
A. Side view of flower, with all the petals and sepals cut off except the labellum, of which the near half is cut away, as well as the upper portion of the near side of the nectary.
B. Front view of the flower, with all sepals and petals removed, except the labellum.
C. One pollinium or pollen-mass, showing the packets of pollen-grains, the caudicle, and viscid disc.
D. Front view of the discs and caudicles of both pollinia within the rostellum, with its lip depressed.
E. Section through one side of the rostellum, with the included disc and caudicle of one pollinium.
F. Packets of pollen-grains, tied together by elastic threads, here extended. (Copied from Bauer.)
ness, and the pollinia would be rendered absolutely useless. That insects often remove one alone of the two pollinia at a time in many kinds of Orchis is certain; it is even probable that they generally remove only one at a time, for the lower and older flowers almost always have both pollinia removed, and the younger flowers close beneath the buds, which will have been seldomer visited, have frequently only one pollinium removed. In a spike of Orchis maculata I found as many as ten flowers, chiefly the upper ones, which had only one pollinium removed; the other pollinium being in place, with the lip of the rostellum well closed up, and all the mechanism perfect for its subsequent removal by some insect.
The description now given of the action of the organs in Orchis mascula applies to O. morio, fusca, maculata, and latifolia, and to Aceras anthropomorpha.* These species pre-
* The separation of this genus is evidently artificial. It is a true Orchis, but with a very short nectary. Dr. Weddell has described ('Annales des Sci. Nat.,' 3° ser. 'Bot.,' tom. xviii. p. 6) the occurrence of numerous hybrids, naturally produced, between this Aceras and Orchis galeata.
sent slight and apparently co-ordinated differences in the length of the caudicle, in the direction of the nectary, in the shape and position of the stigma, but they are not worth detailing. In all, the pollinia undergo, after removal from the anther-cells, the curious movement of depression, which is so necessary to place them in a right position on the insect's head to strike the stigmatic surface of another flower. In Aceras the caudicle is unusually short; the nectary consists of two minute rounded depressions; the stigma is transversely elongated; the two viscid discs lie so close together within the rostellum that they affect each other's outline; this is worth notice, as a step towards the two becoming absolutely confluent, as in O. pyramidalis. Nevertheless, in Aceras a single pollinium is sometimes removed by insects, though more rarely than with the other species.
We now come to Orchis pyramidalis, one of the most highly organised species which I have examined, and which is ranked by several botanists in a distinct genus. The relative position of the parts (Fig. III.) is here consi-
derably different from what it is in O. mascula and its allies. There are two quite distinct rounded stigmatic surfaces (s, s, A) placed on each side of the pouch-formed rostellum. This latter organ, instead of standing some height above the nectary, is brought down (see side view B) so as to overhang and partially to close its orifice. The ante-chamber to the nectary formed by the union of the edges of the labellum to the column, which is large in O. mascula and its allies, is here small. The pouch-formed rostellum is hollowed out on the under side in the middle: it is filled with fluid. The viscid disc is single, of the shape of a saddle (Figs. C and E), carrying on its nearly flat top or seat the two caudicles of the pollinia; of which the two truncated ends firmly adhere to its upper surface. Before the membrane of the rostellum ruptures, it can be clearly seen that the saddle-formed disc forms part of the continuous surface of the rostellum. The disc is partially hidden and kept damp (which is of great importance) by the largely over-folded basal membranes of the two anther-cells. The upper membrane of the disc consists of several layers of minute
FIG. III.
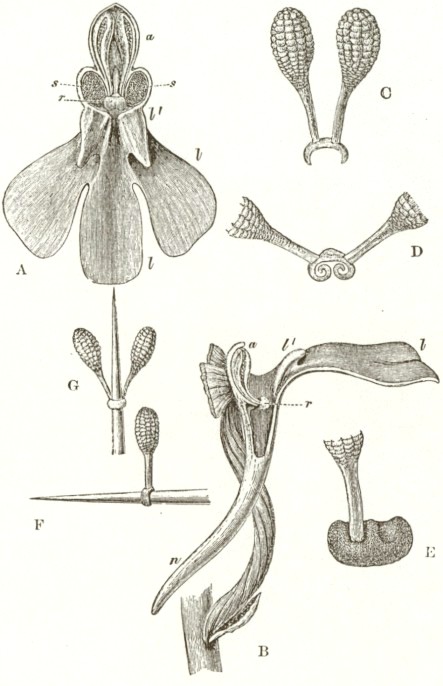
ORCHIS PYRAMIDALIS.
cells, and is therefore rather thick; it is lined beneath with a layer of highly adhesive matter, which is formed within the rostellum. The single saddle-formed disc strictly answers to the two minute, oval, separate discs of membrane to which the two caudicles of O. mascula and its allies are attached: two separate discs have here become completely confluent.
When the flower opens and the rostellum has become symmetrically ruptured, either from a touch or spontaneously (I know not
Description of Fig. III.
a. anther.
s. s. stigma.
r. rostellum.
l. labellum.
l'. guiding plate of the labellum.
n. nectary.
A. Front view, with all sepals and petals removed, except the labellum.
B. Side view, with all sepals and petals removed, with the labellum longitudunally bisected, and with the near side of the upper part of the nectary cut away.
C. The two pollinia attached to the saddle-shaped viscid disc.
D. The disc after the first act of contraction, with no object seized.
E. The disc seen from above, and flattened by force, with one pollinium removed; showing the depression, by which the second act of contraction is effected.
F. The pollinium removed by insertion of a needle into the nectary, after it has clasped the needle by the first act of contraction.
G. The same pollinium after the second act of contraction and depression.
which), the slightest touch depresses the lip, that is, the lower and bilobed portion of the exterior membrane of the rostellum, which projects into the mouth of the nectary. When the lip is depressed, the under and viscid surface of the disc, still remaining in its proper place, is uncovered, and is almost certain to adhere to the touching object. Even a human hair, when pushed into the nectary, is stiff enough to depress the lip or pouch; and the viscid surface of the saddle adheres to it. If, however, the lip be touched too slightly, it springs back and re-covers the under side of the saddle.
The perfect adaptation of the parts is well shown by cutting off the end of the nectary and inserting a bristle at that end; consequently in a reversed direction to that in which nature intended moths to insert their probosces, and it will be found that the rostellum may easily be torn or penetrated, but that the saddle is rarely or never caught. When the saddle sticking to a bristle together with its pollinia is removed, the under lip instantly curls closely inwards, and leaves the orifice of the nectary more open than it was before; but whether this is of any real use to
moths which so frequently visit the flowers, and consequently to the plant, I will not pretend to decide.
Lastly, the labellum is furnished with two prominent ridges (l', Fig. A, B), sloping down to the middle and expanding outwards like the mouth of a decoy; these ridges perfectly serve to guide any flexible body, like a fine bristle of hair, into the minute and rounded orifice of the nectary, which, small as it is, is partly choked up by the rostellum. This contrivance of the guiding ridges may be compared to the little instrument sometimes used for guiding a thread into the fine eye of a needle.
Now let us see how these parts act. Let a moth insert its proboscis (and we shall presently see how frequently the flowers are visited by Lepidoptera) between the guiding ridges of the labellum, or insert a fine bristle, and it is surely conducted to the minute orifice of the nectary, and can hardly fail to depress the lip of the rostellum; this being effected, the bristle comes into contact with the now naked and sticky under surface of the suspended saddle-formed disc. When the bristle is
C
removed, the saddle with the attached pollinia is removed. Almost instantly, as soon as the saddle is exposed to the air, a rapid movement takes place, and the two flaps curl inwards and embrace the bristle. When the pollinia are pulled out by their caudicles, by a pair of pincers, so that the saddle has nothing to clasp, I observed that the tips curled inwards so as to touch each other in nine seconds (see Fig. D), and in nine more seconds the saddle was converted by curling still more inwards into an apparently solid ball. The probosces of the many moths which I have examined, with the pollinia of this Orchis attached to them, were so thin that the tips of the saddle just met on the under side. Hence a naturalist, wo sent me a moth with several saddles attached to its proboscis, and who did not know of this movement, very naturally came to the extraordinary conclusion that the moth had cleverly bored through the exact centres of the so-called sticky glands of some Orchid.
Of course this rapid clasping movement helps to fix the saddle with its pollinia upright on the proboscis, which is very important; but the viscid matter rapidly setting hard would
probably suffice for this end, and the real object gained is the divergence of the pollinia. The pollinia, being attached to the flat top or seat of the saddle, project at first straight up and are nearly parallel to each other; but as the flat top curls round the cylindrical and thin proboscis, or round a bristle, the pollinia necessarily diverge. As soon as the saddle has clasped the bristle and the pollinia have diverged, a second movement commences, which action, like the last, is exclusively due to the contraction of the saddle-shaped disc of membrane, as will be more fully described in the seventh chapter. This second movement is the same as that in O. mascula and its allies, and causes the divergent pollinia, which at first projected at right angles to the needle or bristle (see Fig. F), to sweep through nearly 90 degrees towards the tip of the needle (see Fig. G), so as to become depressed and finally to lie in the same plane with the needle. In three specimens this second movement was effected in from 30 to 34 seconds after the removal of the pollinia from the anther-cells, and therefore in about 15 seconds after the saddle had clasped the bristle.
C2
The use of this double movement becomes evident if a bristle with pollinia attached to it, which have diverged and become depressed, be pushed between the guiding ridges of the labellum into the nectary of the same or another flower (compare Figs. A and G); for the two ends of the pollinia will be found to have acquired exactly such a position that the end of the one strikes against the stigma on the one side, and the end of the other at the same moment strikes against the stigma on the opposite side. These stigmas are so viscid that they rupture the elastic threads by which the packets of pollen are bound together; and some dark-green grains will be seen, even by the naked eye, remaining on the two white stigmatic surfaces. I have shown this little experiment to several persons, and all have expressed the liveliest admiration at the perfection of the contrivance by which this Orchid is fertilised.
As in no other plant, or indeed in hardly any animal, can adaptations of one part to another, and of the whole to other organized beings widely remote in the scale of nature, be named more perfect than those presented by this Orchis, it may be worth while briefly
to sum them up. As the flowers are visited both by day and night-flying Lepidoptera, I do not think that it is fanciful to believe that the bright-purple tint (whether or not specially developed for this purpose) attracts the day-fliers, and the strong foxy odour the night-fliers. The upper sepal and two upper petals form a hood protecting the anther and stigmatic surfaces from the weather. The labellum is developed into a long nectary in order to attract Lepidoptera, and we shall presently give reasons for suspecting that the nectar is purposely so lodged that it can be sucked only slowly (very differently from in most flowers of other families), in order to give time for the curious chemical quality of the viscid matter on the under side of the saddle setting hard and dry. He who will insert a fine and flexible bristle into the expanded mouth of the sloping ridges on the labellum will no doubt that they serve as guides; and that they effectually prevent the bristle or proboscis from being inserted obliquely into the nectary. This circumstance is of manifest importance, for, if the proboscis were inserted obliquely, the saddle-formed disc would become
attached obliquely, and after the compounded movement of the pollinia they could not strike the two lateral stigmatic surfaces.
Then we have the rostellum partially closing the mouth of the nectary, like a trap placed in a run for game; and the trap so complex and perfect, with its symmetrical lines of rupture forming the saddle-shaped disc above, and the lip of the pouch below; and, lastly, this lip so easily depressed that the proboscis of a moth could hardly fail to uncover the viscid disc and adhere to it. But if this did fail to occur, the elastic lip would rise again and re-cover and keep damp the viscid surface. We see the viscid matter within the rostellum attached to the saddle-shaped disc alone, and surrounded by fluid, so that the viscid matter does not set hard till the disc is withdrawn. Then we have the upper surface of the saddle, with its attached caudicles, also kept damp within the bases of the anther-cells, until withdrawn, when the curious clasping movement instantly commences, causing the pollinia to diverge, followed by the movement of depression, which compounded movements together are exactly fitted to cause the ends of the
two pollinia to strike the two stigmatic surfaces. These stigmatic surfaces are sticky enough not to tear off the whole pollinium from the proboscis of the moth, but by rupturing the elastic threads to secure a few packets of pollen, leaving plenty for other flowers.
But let it be observed that, although the moth probably takes a considerable time to suck the nectar of any one flower, yet the movement of depression in the pollinia does not commence (as I know by trial) until the pollinia are fairly withdrawn out of their cells; nor will the movement be completed, and the pollinia be fitted to strike the stigmatic surfaces, until about half a minute has elapsed, which will give ample time for the moth to fly to another plant, and thus effect a union between two distinct individuals. Lastly, we have the wonderful growth of the pollen-tubes and their penetration of the stigma, as well as the mysteries of germination, though these are common to all phanerogamic plants.
Orchis ustulata* resembles O. pyramidalis
* I am greatly indebted to Mr. G. Chichester Oxenden of Broome Park for fresh specimens of this Orchis, and for his
in some important respects, and differs in others. The labellum is deeply channelled; this channel, which replaces the guiding ridges of O. pyramidalis, leads to the small triangular orifice of the short nectary. The upper angle of the triangle is overhung by the rostellum, the pouch of which is rather pointed below. Owing to this position of the rostellum, close to the mouth of the nectary, the stigma is necessarily double and lateral; but we here have an interesting gradation, showing how easily the single and slightly lobed medial stigma of O. maculata would pass through the bilobed stigma of O. mascula into that of O. ustulata, and thence into the truly double stigma of O. pyramidalis; for in O. ustulata, directly under the rostellum, there is a narrow rim, in direct continuity with the two lateral stigmas, and which itself has the character of a true stigma, as it is formed of utriculi, or true stigmatic tissue, exactly like that of the lateral stigmas. The viscid discs are somewhat elongated. The pollinia undergo the
never-tiring kindness in supplying me with living plants, with numerous specimens, and information regarding many of the rarer British Orchids.
usual movement of depression, and in acquiring this position the two diverge slightly, so as to be ready to strike the lateral stigmas.
The divergence seemed due to the manner or direction in which the membrane forming the top of the disc contracted obliquely; but I am not sure of this observation.
I have now described the structure, as seen in fresh specimens, of most of the British species of the genus Orchis. All these species absolutely require the aid of insects for their fertilisation. This is obvious from the fact that the pollinia are so closely embedded in their anther-cells, and the disc with its ball of viscid matter in the pouch-formed rostellum, that they cannot be shaken out by violence. We have also seen numerous contrivances by which the pollinia assume, after an interval of time, a position adapted to strike the stigmatic surface; and this indicates that the pollinia are habitually carried from one flower to another. But to prove that insects are necessary I covered up a plant of Orchis morio under a bell-glass, before any of its pollinia had been removed, leaving three adjoining plants
C3
uncovered; I looked at the latter every morning, and daily found some of the pollinia removed, till all were removed with the exception of the pollinia in one flower low down on one spike, and with the exception of those in one or two flowers at the apex of each spike, which were never removed. I then looked at the perfectly healthy plant under the bell-glass, and it had, of course, all its pollinia in their cells. I tried an analogous experiment with specimens of O. mascula with exactly the same result. It deserves notice that the spikes which had been covered up, when subsequently left uncovered, had not their pollinia removed, and did not, of course, set any seed, whereas the adjoining plants produced plenty of seed; and from this fact I infer that probably there is a proper season for each kind of Orchis, and that insects cease their visits after the proper season has passed, and the regular secretion of nectar has ceased.
I have been in the habit for twenty years of watching Orchids, and have never seen an insect visit a flower, excepting butterflies twice sucking O. pyramidalis and Gymnadenia
conopsea. That bees sometimes visit Orchids* I have evidence in a humble and hive bee sent me by Professor Westwood, with pollinia attached to them; and Mr. F. Bond informs me that he has seen pollinia attached to other species of bees; but I feel almost certain that bees do not habitually visit the common British species of Orchis. On the other hand, I have met with several accounts in entomological works of pollinia having been observed attached to moths. Mr. F. Bond was so kind as to send me a large number of moths in this condition, with permission, at the risk of the destruction of the specimens, to remove the pollinia; and this is quite necessary, in order to ascertain to what species the pollinia belong. Singularly all the pollinia (with the exception of a few from Orchids of the genus Habenaria, pre-
* M. Ménière (in Bull. Bot. Soc. de France, to,. i. 1854, p. 370) says he saw, in Dr. Guépin's collection, bees collected at Saumur with the pollinia of Orchids attached to their heads; and he states that a person who kept bees near the Jardin de la Faculté (at Toulouse?) complained that his bees returned from the garden with their heads charged with yellow bodies, of which they could not free themselves. This is good evidence how firmly the pollinia become attached. There is nothing to show whether the pollinia in these cases belonged to the genus Orchis or to other genera of the family, some of which I know are visited by bees.
sently to be mentioned) belonged to O. pyramidalis. I here give the list of twenty-three species of Lepidoptera, with the pollinia of O. pyramidalis attached to their probosces.
Polyommatus alexis. Lycæna phlæas. Arge galathea. Hesperia sylvanus. Hesperia linea. Syrichthus alveolus. Anthrocera filipendulæ. Anthrocera trifolii.* Lithosia complanata. Leucania lithargyria (two specimens). Caradrina blanda. Caradrina alsines. Agrotis cataleuca. |
Eubolia mensuraria (two specimens). Hadenia dentina. Heliothis marginata (two specimens). Xylophasia sublustris (two specimens). Euclidia glyphica. Toxocampa pastinum. Melanippe rivaria. Spilodes palealis. Spilodes cinctalis. Acontia luctuosa. |
A large majority of these moths and butterflies had two or three pairs of pollinia attached to them, and invariably to the proboscis. The Acontia had seven pair, and the Caradrina no less than eleven pair! The probosces of these two latter moths presented an extraordinary
* I am indebted to Mr. Parfitt for an examination of this moth, which is mentioned in the 'Entomologist's Weekly Intelligencer,' vol. ii. p. 182, and vol. iii. p. 3, Oct. 3, 1857. The pollinia were erroneously thought to belong to Ophrys apifera. The pollen had changed from its natural green colour to yellow; on washing it, however, and drying it, the green tint returned.
arborescent appearance (Fig. IV). The saddle-formed discs adhered to the proboscis, one before the other, with perfect symmetry (as necessarily follows from its insertion having been guided by the ridges on the labellum), each saddle bearing its pair of pollinia. The unfortunate Caradrina, with its proboscis thus encumbered, could hardly have reached the extremity of the nectary, and would soon have been starved to death. These two moths must have sucked many more than the seven and eleven flowers, of which they bore the trophies, for the earlier attached pollinia had lost much of their pollen, showing that they had touched many viscid stigmas.
Fig. IV.
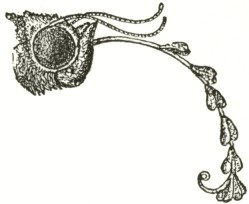
Head and proboscis of Acontia luctuosa with seven pair of the pollinia of Orchis pyramidalis attached to the proboscis.
The list shows, also, how many species of Lepidoptera visit the same kind of Orchis. The Hadena also frequents Habenaria. Probably all the Orchids provided with spur-like nectaries are visited indifferently by many kinds of moths. I have twice observed Gym-
nadenia conopsea, transplanted many miles from its native home, with nearly all its pollinia removed. Mr. Marshall of Ely* has made the same observation on transplanted specimens of O. maculata. I have not sufficient evidence, but I suspect that the Neotteæ and Malaxeæ, which have not tubular nectaries, are frequented by other orders of insects. Listera is generally fertilised by small Hymenoptera; Spiranthes by humble-bees. Mr. Marshall found that fifteen plants of Ophrys muscifera, transplanted to Ely, had not one pollen-mass removed; so it was during the first summer with Epipactis latifolia planted in my own garden; during the following summer six flowers out of ten had their pollinia removed by some insect. These facts possibly indicate that certain Orchids require special insects for their fertilisation. On the other hand, Malaxis paludosa, placed in a bog about two miles from that in which it grew, had most of its pollinia immediately removed.
The list which follows serves to show that
* 'Gardener's Chronicle,' 1861, p. 73. Mr. Marshall's communication was in answer to some remarks of mine on the subject previously published in the 'Gardener's Chronicle,' 1860, p. 528.
in most cases moths perform the work of fertilisation effectually. But the list by no means gives a fair idea how effectually it is done; for I have often found nearly all the pollinia removed; but generally I kept an exact record in exceptional cases only, as may be seen by the appended remarks. Moreover, in most cases, the pollinia which had not been removed were in the upper flowers beneath the buds, and many of these would probably have been subsequently removed. I have often found abundance of pollen on the stigmas of flowers which had not their own pollinia removed, showing that they had been visited by insects; in many other flowers the pollinia had been removed, but no pollen had, as yet, been left on their stigmas.
In the second lot of O. morio, given in the list, we see the injurious effects of the extraordinary cold and wet season of 1860 in the infrequency of the visits of insects, and, consequently, on the fertilisation of this Orchid. Very few seed-capsules were produced this year.
In O. pyramidalis I have examined spikes in which every single expanded flower had its
Number of flowers with both or one pollinium removed. Flowers lately open excluded. |
Number of flowers with only one pollinium removed. These flowers are included in the column to the left. |
Number of flowers with neither pollinium removed. |
|
Orchis morio—Three small plants. N. Kent |
22 |
2 |
6 |
Orchis morio—Thirty-eight plants. N. Kent. These plants were examined after nearly four weeks of extraordinary cold and wet weather in 1860; and therefore under the most unfavourable circumstances |
110 |
23 |
193 |
Orchis pyramidalis—Two plants. N. Kent and Devonshire |
39 |
. . |
8 |
Orchis pyramidalis—Six plants from two protected valleys. Devon |
102 |
. . |
66 |
Orchis pyramidalis—Six plants from a much exposed bank. Devon |
57 |
. . |
166 |
Orchis maculata—One plant. Staffordshire. Of the twelve flowers which had not their pollinia removed, the greater number were young flowers under the buds |
32 |
6 |
12 |
Orchis maculata—One plant. Surrey |
21 |
5 |
7 |
Orchis maculata—Two plants. N. and S. Kent |
28 |
17 |
50 |
Orchis latifolia—Nine plants from S. Kent, sent me by the Rev. B. S. Malden. The flowers were all mature |
50 |
27 |
119 |
Orchis fusca—Two plants. S. Kent. Flowers quite mature, and even withered |
8 |
5 |
54 |
Aceras anthropomorpha—Four plants. S. Kent |
63 |
6 |
34 |
pollinia removed. The forty-nine lower flowers of a spike from Folkestone (sent me by Sir Charles Lyell) actually produced forty-eigth fine seed-capsules; and of the sixty-nine flowers in three other spikes, seven alone had failed to produce capsules. These facts show conclusively how well moths had performed their office of marriage-priests.
In the list, the third lot of O. pyramidalis grew on a steep grassy bank, overhanging the sea near Torquay, and where there were no bushes or other shelter for moths; being surprised how few pollinia had been removed, though the spikes were old, and very many of the lower flowers had withered, I gathered, for comparison, six other spikes from two bushy and sheltered valleys, half a mile on each side of the exposed bank; these spikes were certainly younger, and would probably have had several more of their pollinia removed; but in their present condition we see how much more frequently they had been visited by moths, and consequently fertilised, than those growing on the much exposed bank. The Bee Ophrys and O. pyramidalis, in many parts of England, grow mingled together; and they did
so here, but the Bee Ophrys, instead of being, as usual, the rarer species, was here much more abundant than O. pyramidalis; no one would readily have suspected that probably one chief reason of this difference was, that the exposed situation was unfavourable to moths, and therefore to the seeding of O. pyramidalis; whereas, as we shall hereafter see, the Bee Ophrys is independent of insects.
I counted many spikes of O. latifolia, because, being familiar with the usual state of the closely-allied O. maculata, I was surprised to observe in nine nearly withered spikes how few pollinia had been removed. In one instance, however, I found O. maculata even worse fertilised; for seven spikes, which had borne 315 flowers, produced only forty-nine seed-capsules—that is, each plant on an average produced only seven capsules: in this case the plants had grown in greater numbers close together, forming large beds, than I had ever before observed; and I imagined that there were too many plants for the moths to suck and fertilise. On some other plants, growing at no great distance, I found above thirty capsules on each spike.
Orchis fusca offers a more curious case of imperfect fertilisation. I examined ten fine spikes from two localities in South Kent, sent to me by Mr. Oxenden and Mr. Malden: most of the flowers on these spikes were partly withered, with the pollen mouldy even in the uppermost flowers; hence we may safely infer that no more pollinia would have been removed. I examined all the flowers only in two spikes, on account of the trouble from their withered condition, and the result may be seen in the list, namely, fifty-four flowers with both pollinia in place, and only eight with one or both removed. We see in this Orchid, and in O. latifolia, neither of which had been sufficiently visited by moths, that there are more flowers with one pollinium than with both removed. I casually examined many flowers in the other spikes of O. fusca, and the proportion of pollinia removed was evidently not greater than in the two given in the list. The ten spikes had borne 358 flowers, but, in accordance with the few pollinia removed, only eleven capsules had been formed: five of the ten spikes bore not a single capsule; two spikes had only one, and one bore as many as
four capsules. As corroborating what I have previously said on pollen being often found on the stigmas of flowers which have their own pollinia in place, I may add that, of the eleven flowers which had produced capsules, five had both pollinia still within their now withered anther-cells.
From these facts the suspicion naturally arises that O. fusca is so rare a species in Britain from not being sufficiently attractive to our moths, and consequently not producing a sufficiency of seed. C. K. Sprengel* noticed, that in Germany O. militaris (ranked by Bentham as the same species with O. fusca) is likewise imperfectly fertilised, but more perfectly than our O. fusca; for he found five old spikes bearing 138 flowers, which had set thirty-one capsules; and he contrasts the state of these flowers with those of Gymnadenia conopsea, in which almost every flower produces a capsule.
An allied curious subject remains to be discussed. The existence of a well-developed spur-like nectary seems almost to imply the
* 'Das entdeckte Geheimniss,' etc. s. 404.
secretion of nectar. But Sprengel, a most careful observer, thoroughly searched many flowers of O. latifolia and of O. morio, and could never find a drop of nectar; nor could Krünitz* find nectar either in the nectary or on the labellum of O. morio, fusca, militaris, maculata, and latifolia. I have looked to all the species hitherto mentioned in this work, and could find no signs of nectar; I examined, for instance, eleven flowers of O. maculata, taken from different plants growing in different districts, and taken from the most favourable position on each spike, and could not find under the microscope the smallest bead of nectar. Sprengel calls these flowers "Scheinsaftblumen," or sham-nectar-producers; that is, he believes, for he well knew that the visits of insects were indispensable for their fertilisation, that these plants exist by an organized system of deception. But when we reflect on the incalculable number of plants which have existed for enormous periods of time, all absolutely requiring for each genera-
* Quoted by J. G. Kurr in his 'Untersuchungen über die Bedeutung der Nektarien,' 1833, s. 28. See also 'Das entdeckte Geheimnis, ' s. 403.'
tion insect-agency; when we think of the special contrivances clearly showing that, after an insect has visited one flower and has been cheated, it must almost immediately go to a second flower, in order that impregnation may be effected (of which fact we have the plainest evidence in the large number of pollinia attached to the probosces of those moths which had visited O. pyramidalis), we cannot believe in so gigantic an imposture. He who believes in this doctrine must rank very low the instinctive knowledge of many kinds of moths.
To test the intellect of moths I tried the following little experiment, which ought to have been tried on a larger scale. I removed a few already-opened flowers on a spike of O. pyramidalis, and then cut off about half the length of the nectaries of the six next not-expanded flowers. When all the flowers were nearly withered, I found that thirteen of the fifteen upper flowers with perfect nectaries had their pollinia removed, and two alone had their pollinia still in their anther-cells; of the six flowers with their nectaries cut off, three had their pollinia removed, and
three were still in place; and this seems to indicate that moths do not go to work in a quite senseless manner.
Nature may be said to have tried, but not quite fairly, this same experiment; for Orchis pyramidalis, as shown by Mr. Bentham,* often produces monstrous flowers without a nectary, or with a short and imperfect one. Sir C. Lyell sent me several spikes from Folkestone with many flowers in this condition: I found six without a vestige of a nectary, and their pollinia had not been removed. In about a dozen other flowers, having either short nectaries, or with the labellum imperfect, with the guiding ridges either absent or developed in excess and rendered foliaceous, the pollinia in one alone had been removed, and the ovarium of another flower was swelling. Yet I found that the saddle-formed discs in the first six and in the dozen other flowers were perfect, and that they readily clasped a needle when inserted in the proper place. Moths had removed the pollinia, and had thoroughly well fertilised the perfect flowers on the same spike; so that they must have
* 'Handbook of the British Flora,' 1858, p. 501.
neglected the monstrous flowers, or, if visiting them, the derangement in the complex mechanism had hindered the removement of the pollinia, and prevented their fertilisation.
From these several facts I still suspected that nectar must be secreted by our common Orchids, and I determined to examine O. morio rigorously. As soon as many flowers were open, I began to examine them for twenty-three consecutive days: I looked at them after hot sunshine, after rain, and at all hours: I kept the spikes in water, and examined them at midnight, and early the next morning: I irritated the nectaries with a bristle, and exposed them to irritating vapours: I took flowers which had quite lately had their pollinia removed by insects, of which I had independent proof on one occasion by finding within the nectary grains of some foreign pollen;* and I took other flowers which from their position on the spike would soon have had their pollinia removed; but the nectary was invariably quite dry.
* I may mention that on soaking and separating the laminæ of the proboscis of a moth, which had the pollinia of a Habenaria attached to its head, a surprising number of pollen-grains of some other plant were seen in the water.
I still thought that the secretion might perhaps take place at the earliest dawn, as I have found that the secretion of nectar in flowers of other orders ceases and commences in the most rapid manner. Consequently, as O. pyramidalis is visited (as may be seen in the foregoing list) by butterflies and by several day-flying moths (such as Anthrocera and Acontia), I carefully examined its nectary, taking plants from several localities and the most likely flowers, as just explained; but the glittering points within the nectary were absolutely dry. Hence we may safely conclude that the nectaries of the above-named Orchids neither in this country nor in Germany ever contain nectar.
In examining the nectaries of O. morio and maculata, and especially of O. pyramidalis, I was surprised at the degree to which the inner and outer membranes forming the tube or spur were separated from each other,—also at the delicate nature of the inner membrane, which could be most easily penetrated,—and, lastly, at the quantity of fluid contained between these two membranes. So copious is this fluid, that, having at first merely cut off the
D
ends of the nectaries of O. pyramidalis, and gently squeezing them on glass under the microscope, such large drops of fluid exuded from the cut ends that I concluded that the nectaries certainly did contain nectar; but when I carefully made, without any pressure, a slit along the upper surface, and looked into the tube, I found that the inner surface was quite dry.
I then examined the nectaries of Gymnadenia conopsea (a plant ranked by some botanists as a true Orchis) and of Habenaria bifolia, which are always one-third or two-thirds full of nectar: the inner membrane presented the same structure in being covered with papillæ, but there was a plain difference in the inner and outer membranes being closely united, instead of being, as with the above-named species of Orchis, in some degree separated from each other and charged with fluid. Hence I am led to suspect that moths penetrate the lax inner membrane of the nectaries of these Orchids, and suck the copious fluid between the
two membranes. I am aware that this is a bold hypothesis; for no case is recorded of nectar being contained between the two membranes of a nectary,* or of Lepidoptera penetrating with their delicate probosces even the laxest membrane.
We have seen how numerous and beautifully adapted the contrivances are for the fertilisation of Orchids. We know that it is of the highest importance that the pollinia, when attached to the head or proboscis of an insect, should not fall sideways or backwards. We know that the ball of viscid matter at the extremity of the pollinium rapidly becomes more and more viscid, and sets hard in a few minutes' time: therefore we can see that it would be an advantage to the plant if the moth were delayed in sucking the nectar, so as to give time for the viscid disc to become immoveably affixed. Assuredly moths would be delayed if they had to bore through several points of the inner membrane of the nectary, and to suck the nectar from the intercellular spaces. This explanation of the good thus gained in some degree corroborates the hypo-
* The nearest approach to this supposed case, yet really distinct, is the secretion of nectar in several monocotyledonous plants (as described by Ad. Brongniart in Bull. Soc. Bot. de France, tom. i. 1854, p. 75) from between the two walls (feuillets) which form the divisions of the ovarium. But the nectar in this case is conducted to the outside by a channel; and the secreting surface is homologically an exterior surface.
D2
thesis that the nectaries of the above-named species of Orchis do not secrete nectar externally, but into internal cavities.
The following singular relation supports this view more strongly. I have found nectar within the nectaries of only five British species of Ophreæ, namely, in Gymnadenia conopsea and albida, in Habenaria bifolia and chlorantha, and in Peristylus (or Habenaria) viridis. The first four of these species have the viscid surface of the discs of their pollinia, not enclosed within the pouch, but naked, which by itself shows that the viscid matter has a different chemical nature from that in the species of true Orchis, and does not rapidly set hard when exposed to the air. But to make sure of this I removed the pollinia from their anther-cells, so that the upper as well as the under surfaces of the viscid discs were freely exposed to the air; in Gymnadenia conopsea the disc remained sticky for two hours, and in Habenaria chlorantha for more than twenty-four hours. In Peristylus viridis the viscid disc is covered by a pouch-formed membrane, but this is so minute that botanists have overlooked it. I did not, when examining this species, see the importance of exactly
ascertaining how rapidly the viscid matter set hard; but I copy from my notes the words written at the time: "Disc remains sticky for some time when removed from its little pouch."
Now the bearing of these facts is clear: if, as is certainly the case, the viscid matter of the discs of these five latter species is so viscid as to serve at once for the firm attachment of the pollinia to insects, and does not quickly become more and more viscid and set hard, there could be no use in moths being delayed in sucking the nectar by having to bore through the inner membrane of the nectaries at several points; and in these five species, and in these alone, we find copious nectar ready stored for their use in the open tubular nectaries. If this relation, on the one hand, between the viscid matter requiring some little time to set hard, and the nectar being so lodged that moths are delayed in getting it; and, on the other hand, between the viscid matter being at first as viscid as ever it will become, and the nectar lying all ready for rapid suction, be accidental, it is a fortunate accident for the plant. If not accidental, and I cannot believe it to be accidental, what a singular case of adaptation!
CHAPTER II.
Ophreæ continued—Fly and Spider Ophrys—Bee Ophrys, apparently adapted for perpetual self-fertilisation, but with paradoxical contrivances for intercrossing—The Frog Orchis; fertilisation effected by nectar secreted from two parts of the labellum—Gymnadenia conopsea—Greater and Lesser Butterfly Orchis; their differences and means of fertilisation —Summary on the powers of movement in the pollinia.
WE now come to those genera of Ophreæ which chiefly differ from Orchis in having two separate pouch-formed rostellums,* instead of the two being confluent, as in Orchis. First for the genus Ophrys.
* It is not correct to speak of two rostellums, but the inaccuraty may be forgiven from its convenience. The rostellum strictly is a single organ, formed from the modification of the dorsal stigma and pistil; so that in Ophrys the two pouches and the intermediate space together form the true rostellum. Again, in Orchis I have spoken of the pouch-formed organ as the rostellum, but strictly the rostellum includes the little crest or fold of membrane projecting between the bases of the anther-cells. This folded crest (sometimes converted into a solid ridge) corresponds with the smooth ridge lying between the two pouches in Ophrys, and owes its protuberant and folded condition in Orchis to the two pouches having been brought together and become confluent. This modification will be more fully explained in the seventh chapter.
Fig. V.
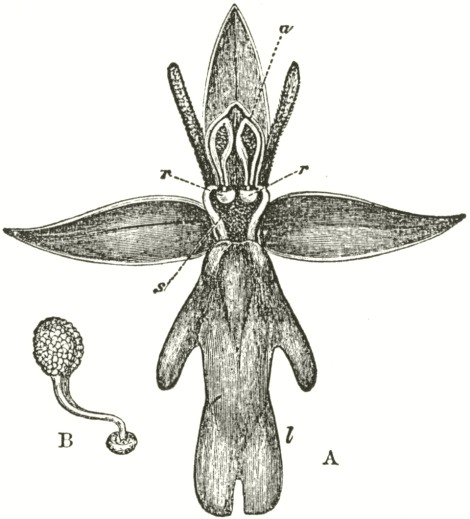
OPHRYS MUSCIFERA, OR FLY OPHRYS.
a. anther.
r r. rostellum.
s. stigma.
l. labellum.
A. Flower viewed in front: the two upper petals are almost cylindrical and hairy: the two rostellums stand a little in advance of the bases of the anther-cells; but this is not shown from the foreshortening of the drawing.
B. One of the two pollinia removed from its anther-cell, and viewed laterally.
In Ophrys muscifera, or the Fly Ophrys, the chief peculiarity is that the caudicle of the pollinium (B) is doubly and almost rectangularly bent. The nearly circular piece of membrane, to the under side of which the ball of viscid
matter is attached, is of considerable size, and plainly forms the summit of the rostellum, instead of forming, as in Orchis, the posterior and upper surface; consequently the attached end of the caudicle, after the flower has expanded, is exposed to the air. As might have been expected from this circumstance, the caudicle is not capable of that movement of depression, characteristic of all the species of Orchis; for this movement is always excited when the upper membrane of the disc is first exposed to the air. The ball of viscid matter is bathed with fluid within the pouch formed by the lower half of the rostellum, and this is necessary, as the viscid matter rapidly sets hard in the air. The pouch is not elastic, and does not spring up when the pollinium is removed. Such elasticity would have been of no use, as there is here a separate pouch for each viscid disc; whereas in Orchis, after one pollinium has been removed, the other has to be kept covered up and ready for action. Hence it would appear as if nature were so economical as to save even superfluous elasticity.
The pollinia cannot, as I have often tried,
be jarred out of the anther-cells by violence. That insects of some kind visit these flowers, though not frequently, and remove the pollinia, is certain, as we shall immediately see. Twice I have found abundant pollen on the stigmas of flowers, in which both their own pollinia were still in their cells; and, no doubt, had I oftener looked, I should have oftener observed this fact. The elongated labellum affords a good standing-place for insects: at its base, just beneath the stigma, there is a rather deep depression, representing the nectary in Orchis; but I could never see a trace of nectar; nor have I observed any insects, often as I have watched these inconspicuous and scentless flowers, even approach them. On each side of the base of the labellum there is a shining knob, with an almost metallic lustre, appearing like two drops of fluid; and if I could in any case believe in Sprengel's sham-nectaries, I should believe it in this instance. What induces insects to visit these flowers I can at present only conjecture. The two pointed pouches, covering the viscid discs, stand not far apart, and project over the stigma: any object gently pushed right against
one of them (in Orchis the push should be directed rather downwards) depresses the pouch, touches and adheres to the viscid ball, and the pollinium is easily removed.
The structure of the flower leads me to believe that small insects (as we shall see in the case of Listera) crawl along the labellum to its base, and that in bending their heads downwards or upwards they strike against one of the pouches; they then fly to another flower with a pollinium attached to their heads, and there bending down to the base of the labellum, the pollinium, owing to its doubly bent caudicle, strikes the sticky stigmatic surface, and leaves pollen on it. Under the next species we shall see good reason to believe that the doubly bent caudicle of the Fly Ophrys serves instead of the usual movement of depression.
That insects do visit the flowers of the Fly Ophrys and remove the pollinia, though not effectually or sufficiently, the following cases show. During several years before 1858 I occasionally examined some flowers, and found that out of 102 only thirteen had one or both pollinia removed. Although at the time
I recorded in my notes that most of the flowers were partly withered, I now think that I must have included a good many young flowers, which might perhaps have been subsequently visited; so I prefer trusting to the following observations.
Number of Flowers. |
||
Both Pollinia or one removed by Insects. |
Both Pollinia in their Cells. |
|
In 1858, 17 plants, bearing 57 flowers, growing near each other |
30 |
27 |
In 1858, 25 plants growing in another spot, and bearing 65 flowers |
15 |
50 |
In 1860, 17 plants, bearing 61 flowers |
28 |
33 |
In 1861, 4 plants bearing 24 flowers from S. Kent, all the previous plants having grown in N. Kent |
15 |
9 |
Total |
88 |
119 |
We here see that, out of 207 flowers examined, not half had been visited by insects. Of the eighty-eight flowers visited, thirty-one had only one pollinium removed. As the visits of insects are indispensable to the fertilisation of this Orchid, it is remarkable (as in the case of Orchis fusca) that this Ophrys has not been rendered more attractive to insects. The num-
ber of seed-capsules produced is proportionably even less than the number of flowers visited by insects. The year 1861 was extraordinarily favourable to this species in this part of Kent, and I never saw such numbers in flower; accordingly, I marked eleven plants, which bore forty-nine flowers, but these produced only seven capsules. Two of the plants each bore two capsules, and three other plants each bore one, so that no less than six plants did not produce a single capsule! What are we to conclude from these facts? Are the conditions of life unfavourable to this species, though it was so numerous in some places this year as to deserve being called quite common? Could the plant nourish more seed; and would it be of any advantage to it to produce more seed? Why does it produce so many flowers, if a larger number of seed would not be advantageous to it? Something seems to be out of joint in the machinery of its life. We shall presently see what a remarkable contrast another species of this same genus, the Ophrys apifera or Bee Ophrys, presents in producing seed.
Ophrys aranifera, or the Spider Ophrys.—I
am indebted to Mr. Oxenden for a few spikes of this rare species. The caudicle (Fig. A) rises straight up from the viscid disc, and is then curved or bent forwards in the same manner, but not in so great a degree, as in the last species.
Fig. VI.
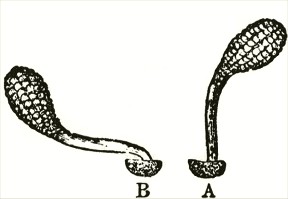
Ophrys aranifera.
A. Pollinium before the act of depression.
B. Pollinium after the act of depression.
The point of attachment of the caudicle to the membrane of the disc is hidden within the bases of the anther-cells, and is thus kept damp; consequently, as soon as the pollinia are exposed to the air, the usual movement of depression takes place, and the pollinia sweep through an angle of about ninety degrees. By this movement the pollinia (supposing them to have become attached to an insect's head) assume a position exactly adapted to strike the stigmatic surface, which is situated, relatively to the pouch-formed rostellums, rather lower down in the flower than in the Fly Ophrys. If we compare the woodcut of the pollinium of the present species, or Spider Ophrys, after the movement, with that of the Fly Ophrys, which is incapable of
movement, it is impossible to doubt that the permanent rectangular bend near the disc of the latter serves the same end as the movement of depression.
I examined fourteen flowers of the Spider Ophrys, several of which were partly withered; both pollinia were removed in none, and in three alone one pollinium had been removed. Hence this species, like the Fly Ophrys, apparently is not much visited by insects.
The anther-cells are remarkably open, so that, in travelling in a box, two pair of pollinia had fallen out, and were sticking by their viscid discs to the flower. Here we have, as throughout nature, evidence of gradation; for though the wide opening of the anther-cells is of no use to this species, it is of the highest importance, as we shall immediately see, to the following species, namely, the Bee Ophrys. So, again, the flexure of the upper end of the caudicle of the pollinium towards the labellum, though of service to the Spider and Fly Ophrys, in order that the pollinium, when removed by insects and carried to another flower, should strike the stigma, is exaggerated in the following species, and serves
for the very different purpose of self-fertilisation.
Ophrys apifera.—In the Bee Ophrys we meet with widely different means of fertilisation as compared with the other species of the genus, and, indeed, as far as I know, with all other Orchids. The two pouch-formed rostellums, the viscid discs, and the position of the
Fig. VII.
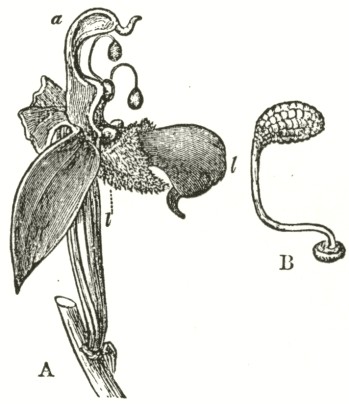
OPHRYS APIFERA, OR BEE OPHRYS.
a. anther.
l l. labellum.
A. Side view of flower, with the upper sepal and the two upper petals removed. One pollinium, with its disc still in its pouch, is represented as just falling out of the anther-cell; and the other has fallen almost to its full extent, opposite to the hidden stigmatic surface.
B. Pollinium in the position in which it lies embedded.
stigma, are nearly the same as in other species of Ophrys; but to my surprise, I have observed that the distance of the two pouches from each other, and the shape of the mass of pollen-grains, are variable. The caudicles of the pollinia are remarkably long, thin, and flexible, instead of being, as in all the other Ophreæ, rigid enough to stand upright. They are necessarily curved forward at their upper ends, owing to the shape of the anther-cells; and the pear-shaped masses of pollen lie embedded high above and directly over the stigma. The anther-cells naturally open soon after the flower is fully expanded, and the thick ends of the pollinia fall out, the viscid discs still remaining in their pouches. Slight as is the weight of the pollen, yet the caudicle is so thin, and soon becomes so flexible, that, in the course of a few hours, they sink down, until they hang freely in the air (see lower pollen-mass in Fig. A) exactly opposite to and in front of the stigmatic surface. When in this position a breath of air, acting on the expanded petals, sets the flexible and elastic caudicles vibrating, and they almost immediately strike the viscid stigma, and, being there secured,
impregnation is effected. To make sure that no other aid was requisite, though the experiment was superfluous, I covered up a plant under a net, so that some wind, but no insects, could pass in, and in a few days the pollinia had become attached to the stigmas; but the pollinia of a spike kept in water in a still room, remained free, suspended in front of the stigma.
Robert Brown* first observed that the structure of the Bee Ophrys is adapted for self-fertilisation. When we consider the unusual and perfectly-adapted length, as well as the remarkable thinness, of the caudicles of the pollinia; when we see that the anther-cells naturally open, and that the masses of pollen, from their weight, slowly fall down to the exact level of the stigmatic surface, and are there made to vibrate to and fro by the slightest breath of wind till the stigma is struck; it is impossible to doubt that these points of structure and function, which occur in no other British Orchid, are specially adapted for self-fertilisation.
* 'Transact. Linn. Soc.' vol. xvi. p. 740. Brown erroneously believed that this peculiarity was common to the genus. As far as the British species are concerned, it applies to this one alone of the four species.
The result is what might have been anticipated. I have often noticed that the spikes of the Bee Ophrys apparently produced as many seed-capsules as flowers; and near Torquay I carefully examined many dozen plants, some time after the flowering season; and on all I found from one to four, and occasionally five, fine capsules, that is, as many capsules as there had been flowers; in extremely few cases (excepting a few deformed flowers, generally on the summit of the spike) could a flower be found which had not produced a capsule. Let it be observed what a contrast this case presents with that of the Fly Ophrys, which requires insect agency, and which from forty-nine flowers produced only seven capsules!
From what I had seen of other British Orchids, I was so much surprised at the self-fertilisation of this species, that during many years I have looked at the state of the pollen-masses in hundreds of flowers, and I have never seen, in a single instance, reason to believe that pollen had been brought from one flower to another. Excepting in a few monstrous flowers, I have never seen an instance of the pollinia failing to reach their own stig-
mas. In a very few cases I have found one pollinium removed, but in some of these cases the marks of slime led me to suppose that slugs had devoured them. For instance, in 1860, I examined in North Kent twelve spikes bearing thirty-nine flowers, and three of these had one pollinium removed, all the other pollinia being glued to their own stigmas. In another lot, from another locality, however, I found the unparalleled case of two flowers with both pollinia removed, and two others with one removed. I have examined some flowers from South Kent with the same result. Near Torquay I examined twelve spikes bearing thirty-eight flowers, and in these one single pollinium alone had been removed. We must not forget that blows from animals or storms of wind might occasionally cause the loss of a pollinium.
In the Isle of Wight Mr. A. G. More was so kind as to examine carefully a large number of flowers. He observed that in plants growing singly both pollinia were invariably present. But on taking home several plants, from a large number growing in two places, and selecting plants which seemed to have had
some pollinia removed, he examined 136 flowers: of these ten had lost both pollinia, and fourteen had lost one; so here there seems at first evidence of the pollinia having been removed by their adhesion to insects; but then Mr. More found no less than eleven pollinia (not included in the above cases of removal) with their caudicles cut or gnawed through, but with their viscid discs still in their pouches, and this proves that some other animals, not insects, probably slugs, had been at work. Three of the flowers were much gnawed. Two pollinia, which had apparently been thrown out by strong wind, were sticking to the sepals, and three pollinia were found loose in his collecting box, so that it is very doubtful whether many, if indeed any, of the pollinia had been removed by adhesion to insects. I will only add that I have never seen any insect visit these flowers.* Robert Brown imagined that the flowers resembled bees in order to deter insects from visiting
* Mr. Gerard E. Smith, in his 'Catalogue of Plants of S. Kent, ' 1829, p. 25, says: "Mr. Price has frequently witnessed attacks made upon the Bee Orchis by a bee, similar to those of the troublesome Apis muscorum." What this sentence means I cannot conjecture.
them; I cannot think this probable. The equal or greater resemblance of the Fly Ophrys to an insect does not deter the visits of some unknown insect, which, in that species, are indispensable for the act of fertilisation.
Whether we look to the structure of the several parts of the flower as far as hitherto described, or to the actual state of the pollinia in numerous plants taken during different seasons from different localities, or to the number of seed-capsules produced, the evidence seems conclusive that we here have a plant which is self-fertilised for perpetuity. But now let us look to the other side of the case. When an object is pushed (as in the case of the Fly Ophrys) right against one of the pouches of the rostellum, the lip is depressed, and the large extremely viscid disc adheres firmly to the object, and the pollinium is removed. Even after the pollinia have naturally fallen out of their cells and are glued to the stigma, their removal can sometimes be thus effected. As soon as the disc is drawn out of the pouch the movement of depression commences by which the pollinium would be brought in front of an insect's head ready to strike the stigma. When
a pollen-mass is placed on the stigma and then withdrawn, the elastic threads by which the packets are tied together, break, and leave several packets on the viscid surface. In all other Orchids the meaning of these several contrivances—namely, the downward movement of the lip of the rostellum when gently pushed—the viscidity of the disc—the act of depression of the caudicle after the disc has been removed—the rupturing of the elastic threads by the viscidity of the stigma, so that the pollen may be left on several stigmas—is unmistakably clear. Are we to believe that these contrivances in the Bee Ophrys are absolutely purposeless, as would certainly be the case if this species is perpetually self-fertilised? If the discs had been small or only viscid in a slight degree, if the other related contrivances had been imperfect in any degree, we might have concluded that they had begun to abort; that Nature, if I may use the expression, seeing that the Fly and Spider Ophrys were imperfectly fertilised and produced few seed-capsules, had changed her plan and effected complete and perpetual self-fertilisation, in order that more seed might be produced.
The case is perplexing in an unparalleled degree, for in the same flower we apparently have elaborate contrivances for directly opposed objects.
We have already seen many curious structures and movements, as in Orchis pyramidalis, which evidently lead to the fertilisation of one flower by the pollen of another flower, and we shall meet with numerous other and very different contrivances for the same object throughout the whole great Orchidean Family. Hence it is impossible to doubt that some great good is derived from the union of two distinct flowers, often borne on distinct plants; but the good in the case of the Fly and Spider Ophrys is gained at the expense of much lessened fertility. In the Bee Ophrys great fertility is gained at the expense if apparently perpetual self-fertilisation; but the contrivances are still present which are assuredly adapted to give an occasional cross with another individual; and the safest conclusion, as it seems to me, is, that under certain unknown circumstances, and perhaps at very long intervals of time, one individual of the Bee Ophrys is crossed by another. Thus the generative
functions of this plant would be brought into harmony with those of other Orchidaceæ, and, indeed, with those of all other plants, as far as I have been able to make out their structure.
Ophrys arachnites.—This form is considered by some high botanical authorities as a mere variety of the varying Bee Ophrys. Mr. Oxenden sent me two spikes bearing seven flowers. The anther-cells do not stand so high above the stigma, and do not overhang it so much, as in the Bee Ophrys. The mass of pollen-grains is generally more elongated. The upper part of the caudicle is curved forward, and the lower part undergoes the movement of depression, as in the Spider and Bee Ophrys. The caudicle in length compared with that of the Bee Ophrys is only as two to three, or even as two to four; though thus relatively shorter, it is as thick and broad as that of the Bee Ophrys: it is much more rigid, so that, when the upper end of the pollinium is forced out of its anther-cell, the sticky disc remaining in the pouch, it can only with difficulty be bent
Fig. VIII.
Pollinium of Ophrys arachnites.
down to the stigma. We here see no adaptation for self-fertilisation.
The seven flowers sent me had evidently long been fully expanded, and the spikes, having travelled by railway, must have been well shaken, yet in six of the flowers both pollinia remained in their anther-cells; in the seventh, both pollinia adhered to the stigma with their discs still in their pouches; but this flower was much withered, and might have been crushed. Of the six flowers, three were so old that the pollen was mouldy and the petals discoloured; yet, as just stated, the pollinia were still in their cells. After having examined so many hundred flowers of the Bee Ophrys, I have never seen such a case. Considering this important functional difference in O. apifera and arachnites, and the lesser differences in the structure of their pollinia, which are likewise of functional signification, and the slight differences in their flowers, it seems to me that, until these forms can be shown to be connected by intermediate varieties, we must rank O. arachnites as a good species, more closely allied to O. aranifera in its manner of fertilisation than to O. apifera.
Herminium monorchis.—The Musk Orchis is generally spoken of as having naked glands or discs, but this is not strictly correct. The disc is of unparalleled size, nearly equalling the mass of pollen-grains: it is subtriangular in shape, unsymmetrical, somewhat resembling a helmet, but with one side protuberant: it is formed of hard membrane; the hollow base alone is viscid, and this rests on and is covered by a narrow strip of membrane, which is easily pushed away, and answers to the pouch in Orchis. The whole upper part of the helmet answers to the minute oval bit of membrane to which the caudicle is attached in Orchis, and which is of larger size and convex in the the Fly Ophrys. When the lower part of the helmet is moved by any pointed object, the point so readily slips into the hollow base, and is there so firmly held by the viscid matter, that it appears adapted to stick to some prominent part of an insect's head. The caudicle is short and very elastic; it is attached not to the apex of the helmet, but to the hinder end; if it had been attached to the apex, the point of attachment would have been freely exposed to the air, and would not have
contracted and caused the movement of depression in the pollinia when removed from their anther-cells. This movement is well marked, and is necessary to bring the end of the pollen-mass into a proper position to strike the stigma. The two viscid discs stand wide apart. There are two transverse stigmatic surfaces, meeting by their points in the middle; but the broad part of each lies directly under the disc.
The labellum is upturned, making the flower almost tubular. As far as I could ascertain, an insect, in crawling out of, or into, the flower, would be apt to strike against the upper and extraordinarily protuberant ends of the helmet-like discs, and so displace the inferior viscid surfaces, which would adhere to its head or body. There is so deep a hollow at the base of the labellum as almost to deserve to be called a nectary; but I could not perceive any nectar. The flowers are very small and inconspicuous, but emit a strong musky smell, especially at night. They seem highly attractive to insects; in a spike with only seven flowers recently open, four had both pollinia, and one had a single pollinium removed.
Peristylus (or Habenaria) viridis.—The Frog Orchis has also been described as having its viscid discs naked, which is incorrect. The two small pouches stand wide apart. The ball of viscid matter is oval, and does not soon set hard; its surface is protected by a minute pouch. The upper membranous surface of the disc is large, and as in the Fly Ophrys (O. muscifera) the point of junction with the caudicle is freely exposed to the air, and does not cause the pollinium to undergo the often described movement of depression. But the caudicles are not doubly bent as in the Fly Ophrys. The stigmatic surface is small and medial; and though the anther-cases are somewhat inclined backwards and converge a little at their upper ends, thus affecting the position of the pollinia when attached to any object, yet it is at first difficult to understand how the
Fig. IX.
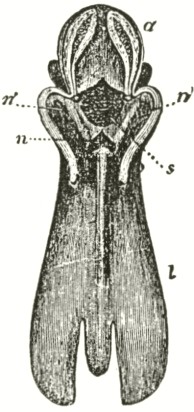
Peristylus viridis, or Frog Orchis.
Front view of flower.
a. anther.
s . stigma.
n. orifice of central nectary.
n'. lateral nectaries.
l. labellum.
pollinia, when removed by insects, can strike the stigma.
The explanation is rather curious. The elongated labellum forms a rather deep hollow in front of the stigma, and in this hollow, but some way in advance of the stigma, a minute slit-like orifice (n) leads into the short bilobed nectary. Hence an insect, in order to suck the nectar with which the nectary is filled, would have to bend down its head in front of the stigma. The labellum has a medial ridge, which would probably induce the insect to alight on either side; but, apparently to make sure of this, besides the true nectary, there are two spots (n') on each side at the base of the labellum, bordered by its prominent edges, directly under the two pouches, which secrete drops of nectar. Now let us suppose an insect to alight, probably on one side of the labellum, and first lick up the drop of nectar on either side; from the position of the pouches exactly over these drops, it would almost certainly get the pollinium of that side attached to its head; if it were then to go to the mouth of the true nectary, it would assuredly strike the pollinium against the stigma. So that we see this unique
case of nectar being secreted from the basal edges of the labellum as well as within the short medial nectary, replaces the power of movement in the pollinia, so general with other Orchids, and replaces the doubly bent caudicles of the Fly Ophrys.
As I have described the case, the flower would receive its own pollen; but if the insect first exhausted the richer source of nectar within the nectary, and afterwards licked up the lateral drops, it would not till then get the pollinia attached to its head, and, flying to another flower, a union would be effected between two distinct flowers or two distinct plants. If, indeed, the insect were to suck the lateral drops first, from what Sprengel has observed in the case of Listera,* the insect would perhaps be disturbed by the attachment of the pollinium, and would not go on sucking immediately, but would fly to another flower, and thus a union between distinct individuals would ensue.
I am endebted to the Rev. B. S. Malden of Canterbury for two spikes of the Frog Orchis. Several of the flowers had one pol-
* 'Das entdeckte Geheimniss der Natur,' s. 407.
linium removed, and one flower had both removed.
We now come to two genera, namely, Gymnadenia and Habenaria, including four British species, which really have uncovered viscid discs. The viscid matter, as before remarked, is of a somewhat different nature from that in Orchis, and does not rapidly set hard. Their nectaries are stored with nectar. With respect to the uncovered condition of the discs, the last species, or Peristylus viridis, is in an almost intermediate condition. The four following forms compose a much broken series. In Gymnadenia conopsea the viscid discs are narrow and much elongated, and lie close together; in G. albida they are less elongated, but still approximate; in Habenaria bifolia they are oval and far apart; and, lastly, in Hab. chlorantha they are circular and much farther apart.
Gymnadenia conopsea.—In general appearance this plant resembles pretty closely some species of true Orchis. The pollinia differ in having naked, narrow, strap-shaped discs, which are nearly as long as the caudicles
Fig. X.
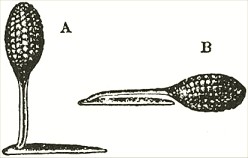
Gymnadenia conopsea.
A. Pollinium, before the act of depression.
B. Pollinium, after the act of depression, but before it has closely clasped the disc.
(Fig. X). When the pollinia are exposed to the air the caudicle is depressed in from 30 to 60 seconds; and as its anterior surface is slightly hollowed out, it closely clasps the upper membranous surface of the disc. The mechanism of this movement will be described in the last chapter. The elastic threads by which the packets of pollen are bound together are unusually weak, as is likewise the case with the two following species of Habenaria: this was well shown by the state of the specimens which had been kept in spirits of wine. This weakness apparently stands in relation to the viscid matter of the discs not setting hard and dry as in Orchis; so that a moth with a pollinium attached to its proboscis might be enabled to visit several flowers and not have the whole pollinium dragged off by the first stigma which was struck. The two strap-shaped discs lie close together, and form the roof of the mouth of the nectary. They are
not enclosed, as in Orchis, by a lower lip or pouch, so that the structure of the rostellum is simpler. When we come to treat of the homologies of the rostellum we shall see that this difference is due to a small change, namely, to the lower and exterior cells of the rostellum resolving themselves into viscid matter; whereas in Orchis the exterior surface retains its early cellular or membranous condition.
As the two viscid discs form the roof of the mouth of the nectary, and are thus brought down near the labellum, the two stigmas, instead of being confluent and standing beneath the rostellum, are necessarily lateral and separate. They form two protuberant, almost horn-shaped, processes on each side of the mouth of the nectary. That their surfaces are really stigmatic I ascertained by finding them deeply penetrated by a multitude of pollen-tubes. As in the case of O. pyramidalis, it is a pretty little experiment to push a fine bristle into the narrow mouth of the nectary, and to observe how certainly the narrow elongated viscid discs, forming the roof, stick to the bristle. When the bristle is withdrawn, the pollinia are withdrawn, adhering to its upper side, and
slightly divergent owing apparently to their original position in the anther-cells. They then quickly depress themselves till they lie in the same plane with the bristle; and if the bristle, held in the same relative position, be now inserted into the nectary of another flower, the two ends of the pollinia accurately strike the two stigmatic surfaces lying close on each side of the mouth of the nectary. I am, however, not quite sure that I understand the cause of the divergence of the pollinia, for I find that moths often remove one pollinium alone; and this fact leads me to suspect that they insert their probosces obliquely into the nectary.
The flowers smell sweet, and the abundant nectar always contained in their nectaries seems highly attractive to Lepidoptera, for the pollinia are soon and effectually removed. For instance, in a spike with forty-five open flowers, forty-one had their pollinia removed, or had pollen left on their stigmas: in another spike with fifty-four flowers, thirty-seven had both pollinia, and fifteen had one pollinium, removed; so that only two flowers in the whole spike had neither pollinium removed.
Gymnadenia albida.—The structure of this
flower resembles in most points that of the last species; but, owing to the upturning of the labellum, it is rendered almost tubular. The naked glands are minute, but elongated and approximate. The stigmatic surfaces are partly lateral and divergent. The nectary is short, and full of nectar. Minute as the flowers are, they seem highly attractive to insects: of the eighteen lower flowers in one spike, ten had both pollinia, and seven had one, removed; in some other and older spikes all the pollinia had been removed, except from two or three flowers at the summit.
Habenaria chlorantha.—The pollinia of the Large Butterfly Orchis differ considerably from those of the species hitherto mentioned. The two anther-cells are separated from each other by a wide space of connective membrane; the pollinia slope backwards (Fig. XI.), and the viscid discs are brought out in advance of the stigmatic surface, and front each other. In relation to this position of the discs, the caudicles and pollen-masses are much elongated. The viscid disc is circular, and, in the early bud, consists of a mass of cells, of which the exterior layers (answering to the lip or pouch
Fig. XI.
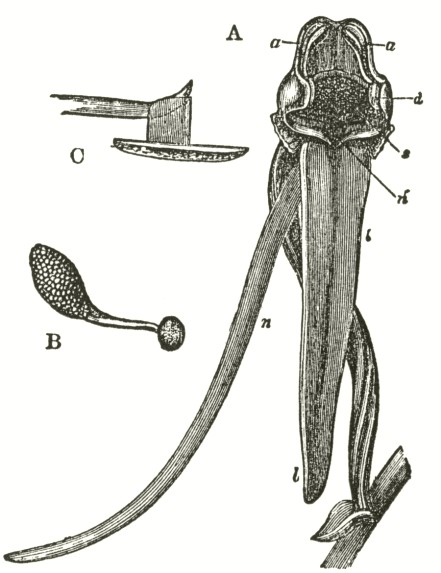
HABENARIA CHLORANTHA, OR LARGE BUTTERFLY ORCHIS.
a a. anther.
d. disc.
s. stigma.
n. nectary.
n'. orifice of nectary.
l. labellum.
A. Flower viewed in front, with all the sepals and petals removed except the labellum with its nectary.
B. A pollinium. (This has hardly a sufficiently elongated appearance.) The drum-like pedicel is hidden behind the disc.
C. Diagram, giving a section through the viscid disc, which is formed of an upper membrane with a layer of viscid matter beneath, and through the drum-like pedicel, and through the lower end of the caudicle.
in Orchis) resolve themselves into adhesive matter. This matter has the property of remaining adhesive for at least twenty-four hours after the pollinium has been removed from its cell. The disc, externally covered with a thick layer of adhesive matter (see Sect. C, which stands so that the layer of viscid matter is below), is produced on its opposite and embedded side into a short drum-like pedicel. This pedicel is continuous with the membranous portion of the disc, and is formed of the same tissue. At the embedded end of the pedicel, the caudicle of the pollinium is attached in a transverse direction, and its extremity is prolonged, as a bent rudimentary tail, just beyond the drum. The caudicle is thus united to the viscid disc in a very different manner, and in a plane at right angles, to what occurs in other British Orchids. In the short drum-like pedicel, we see a small development of the long pedicel of the rostellum, which in many exotic Vandeæ is so conspicuous, and which connects the viscid disc with the true caudicle of the pollinium.
The drum-like pedicel is of the highest importance, not only by rendering the viscid
disc more prominent and more likely to stick to the face of an insect whilst inserting its proboscis into the nectary beneath the stigma, but on account of its power of contraction. The pollinia lie inclined backwards in their cells (see Fig. A), above and some way on each side of the stigmatic surface; if attached in this position to the head of an insect, the insect might visit any number of flowers, and no pollen could be left on the stigma. But observe what takes place: in a few seconds after the inner end of the drum-like pedicel is removed from its embedded position and exposed to the air, one side of the drum contracts, and this contraction draws the thick end of the pollinium inwards, so that the caudicle and the viscid surface of the disc are no longer parallel, as they were at first, and as they are presented in the section C. At the same time the drum rotates through nearly a quarter of a circle, and this moves the caudicle downwards, like the hand of a clock, depressing the thick end of the pollinium or mass of pollen-grains. After this double movement, the right-hand disc, for instance, being supposed to be affixed to the right side of an insect's face,
when the insect, after a short interval of time, visits another flower, the pollen-bearing end of the pollinium will have moved downwards and inwards, and will now infallibly strike the viscid surface of the stigma, situated in the middle beneath and between the two anther-cells.
The little rudimentary tail of the caudicle projecting beyond the drum-like pedicel is an interesting point to those who believe in the modification of species; for it shows us that the disc has moved a little inwards, and that primordially the two discs stood even still further in advance of the stigma than they do at present. It shows us that the parent-form approached a little more closely in structure to that extraordinary Orchid the Bonatea speciosa of the Cape of Good Hope.
The remarkable length of the nectary, containing much nectar, the white colour of the conspicuous flower, and the strong sweet odour emitted at night, all show that this plant depends for it fertilisation on the larger nocturnal Lepidoptera. I have often found spikes with almost all the pollinia removed. From the lateral position and distance of the two viscid
discs from each other, the same moth would generally remove only one pollinium at a time; and in a spike which had not as yet been much visited, three flowers had both pollinia, and eigth flowers had only one pollinium, removed. From the position of the discs it might have been anticipated that they would adhere to the side of the head or face of moths; and Mr. F. Bond sent me a specimen of Hadena dentina with one eye covered and blinded by a disc, and a specimen of Plusia v. aureum with a disc attached to the edge of the eye. Although the discs are so extremely adhesive that almost all the pollinia of a bunch of flowers, when carried in the hand, will be removed by the shaken petals and sepals touching the discs, yet it is certain that moths, perhaps the smaller species, often visit these flowers without removing the pollinia; for on carefully examining the discs of a larger number of pollinia still in their cells I found minute Lepidopterous scales glued to them.
Habenaria bifolia, or Lesser Butterfly Orchis.—I am aware that this form and the last are considered by Mr. Bentham and some other botanists as mere varieties of each other; for
it is said that intermediate gradations occur in the position of the viscid discs. But we shall immediately see that the two forms differ in a great number of characters, not to mention the differences in general aspect and in the stations inhabited; with which we are not here concerned. Should these two forms be hereafter proved to graduate at the present day into each other, it would be a remarkable case of variation; and I, for one, should be as much pleased as surprised at the fact, for these two forms certainly differ from each other more than do most species of the genus Orchis.
The viscid discs of the Lesser Butterfly Orchis are oval; they face each other, and stand far closer together than in the last species; so much so, that in the bud, when their surfaces are cellular, they almost touch. They are not placed so low down relatively to the mouth of the nectary. The viscid matter is of a somewhat different chemical nature, as shown by its much greater viscidity, when moistened, after having been long dried, or after being kept in weak spirits. The drum-like pedicel can hardly be said to be present, and is represented by a longitudinal ridge,
Fig. XII.
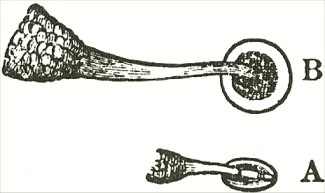
B. Disc and caudicle of H. chlorantha, seen from above, with the drum-like pedicel fore-shortened.
A. Disc and caudicle of Habenaria bifolia, seen from above.
truncated at the end where the caudicle is attached, and there is hardly a vestige of the rudimentary tail of the caudicle. In Fig. XII. the discs of both species, of the proper proportional sizes, are represented as seen from vertically above. The pollinia, after removal from their cells, undergo the same movements; but the inward movement seemed to be greater than in the last species, in conformity with the position of the stigma. In both forms the movement is well seen by removing, with a pair of pincers, a pollinium by the thick end, and holding it motionless under the microscope, when the plane of the viscid disc will be seen to move through an angle of at least 45°. The caudicles of the Lesser Butterfly Orchis are relatively very much shorter than in the other species; the little packets of pollen are shorter, whiter, and, in a mature flower, separate much more readily from each other. Lastly, the stigmatic surface is differently shaped, being more plainly tripartite,
with two lateral prominences, situated beneath the viscid discs. These prominences contract the mouth of the nectary, making it subquadrangular. Hence I cannot doubt that the Larger and Lesser Butterfly Orchids are distinct species, masked by close external similarity.
As soon as I had examined the Lesser Butterfly Orchis, I felt convinced, from the position of the viscid discs, that it would be fertilised in a different manner from the Larger Butterfly Orchis; and now, owing to the kindness of Mr. F. Bond, I have examined two moths, namely, Agrotis segetum and Anaitis plagiata, one with three pollinia, and the other with five pollinia, attached not to the side of the face as in the last species, but to the base of the proboscis. I may remark that the pollinia of these two species of Habenaria, when attached to moths, could be distinguished at a glance.
We have now finished with the Ophreæ; but before passing on to the next Division; I will recapitulate the chief facts on the movements of the pollinia, all due to the nicely
regulated contraction of that small portion of membrane (together with the pedicel in the case of Habenaria) lying between the layer or ball of adhesive matter and the extremity of the caudicle. In most of the species of Orchis the stigma lies directly beneath the anther-cells, and the pollinia simply move vertically downwards. In Orchis pyramidalis and in Gymnadenia there are two lateral and inferior stigmas, and the pollinia move downwards and outwards, diverging at the proper angle (by a different mechanism in the two cases), so as to strike the two lateral stigmas; in Habenaria the stigmatic surface lies beneath and between the two widely separated anther-cells, and the pollinia here again move downward, but, at the same time, converge. A poet might imagine, that whilst the pollinia are borne from flower to flower through the air, adhering to a moth's body, they voluntarily and eagerly place themselves, in each case, in that exact position in which alone they can hope to gain their wish and perpetuate their race.
CHAPTER III.
Epipactis palustris; curious shape of the labellum and its apparent importance in the fructification of the flower—Cephalanthera grandiflora; rostellum aborted; early penetration of the pollen-tubes; case of imperfect self-fertilisation; fertilisation aided by insects—Goodyera repens—Spiranthes autumnalis; perfect adaptation by which the pollen of a younger flower is carried to the stigma of an older flower on another plant.
WE now come to another great tribe of British Orchids, the Neotteæ, which have a free anther standing behind the stigma: their pollen-grains are tied together by fine elastic threads, which partially cohere and project at the upper end of the pollen-mass, and are attached (with some exceptions) to the back of the rostellum. Consequently the pollen-masses have no true and distinct caudicles. In one genus alone (Goodyera) the pollen-grains are collected into packets as in Orchis. Epipactis and Goodyera agree pretty closely in their manner of fertilisation with the Ophreæ, but are more simply organized: Spiranthes comes under the same category, but in some respects is differently modified. Cephalanthera seems to be a de-
Fig. XIII.
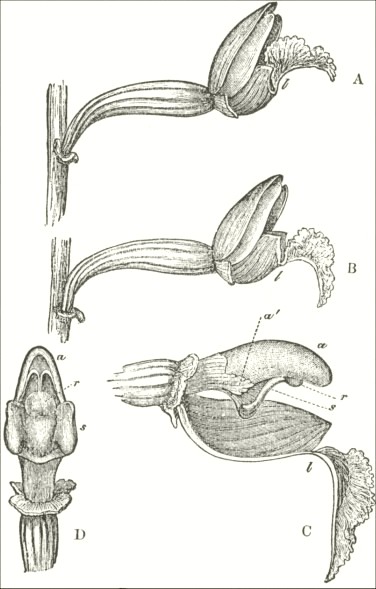
EPIPACTIS PALUSTRIS.
a. anther, with the two open cells seen in the front view D.
a'. rudimentary anther, or auricle, referred to in a future chapter.
r. rostellum.
s. stigma.
l. labellum.
A. Side view of flower (with the lower sepals alone removed) in its natural position.
B. Side
graded or simplified Epipactis; as it does not possess a rostellum—that eminently characteristic organ—and as its pollen-grains are single, it bears almost the same sort of relation to other Orchids, which a wingless bird does to other birds.
Epipactis palustris.*—The lower part of the stigma is bilobed and projects in front of the column (see s in the side and front views, C, D, Fig. XIII.). On its square summit a single, small, nearly globular rostellum is seated. The anterior face of the rostellum (r, C, D) projects a little beyond the surface of the upper part of the stigma, and this is of importance. In the early bud the rostellum consists of a friable mass of cells, with the exterior surface rough:
__________
B. Side view of flower, with the distal portion of the labellum depressed, as if by the weight of an insect.
C. Side view of flower, with all the sepals and petals removed, excepting the labellum, of which the near side has been cut away; the massive anther is seen to be of large size.
D. Front view of column, with all the sepals and petals removed: the rostellum has shrunk down a little in the specimen figured, and ought to have stood higher, so as to hide more of the anther-cells.
__________
* I am much indebted to Mr. A. G. More, of Bembridge in the Isle of Wight, for repeatedly sending me fresh specimens of this beautiful Orchis.
these superficial cells undergo a great change during development, and become converted into a soft, smooth, highly elastic membrane or tissue, so excessively tender that it can be penetrated by a human hair; when thus penetrated, or when slightly rubbed, the surface becomes milky and in some degree viscid, so that the pollen-grains adhere to it. In some cases, though I observed this more plainly in E. latifolia, the surface of the rostellum apparently becomes milky and viscid without having been touched. This exterior soft elastic membrane forms a cap to the rostellum, and is internally lined with a layer of much more adhesive matter; this matter, when exposed to the air, dries in from five to ten minutes. By a slight upward and backward push with any object, the whole cap, with its viscid lining, is removed with the greatest ease; and a minute square stump, the basis of the rostellum, alone is left on the summit of the stigma.
In the bud-state the anther stands quite free behind the rostellum and stigma; it opens longitudinally whilst the flower is still unexpanded, and exposes the two oval pollen-masses,
which are now loose in their cells. The pollen consists of spherical granules, cohering in fours, but not affecting each other's shapes; and these compound grains are tied together by fine elastic threads. These threads are collected into bundles, extending longitudinally along the middle of the front face of each pollinium, where it comes into contact with the back of the uppermost part of the rostellum. From the number of these threads this middle line looks brown, and each pollen-mass here shows a tendency to be divided longitudinally into two halves: in all these respects there is a close general resemblance to the pollinia of the Ophreæ.
The line where the parallel threads are the most numerous is the line of greatest strength; elsewhere the pollinia are extremely friable, so that the masses of pollen can be easily broken off. In the bud-state the rostellum is curved a little backwards, and is pressed against the recently-opened anther; and the above-mentioned slightly projecting bundles of threads become firmly attached to the posterior flap of the membranous cap of the rostellum. The point of attachment lies a little beneath
the summit of the pollen-masses; but the exact point is somewhat variable, for I have met with specimens in which the attachment was one-fifth of the length of the pollen-masses from their summits. This variability is so far interesting, as it is a step leading to the structure of the Ophreæ, in which the confluent threads, or caudicles, spring from the lower ends of the pollen-masses. After the pollinia are firmly attached by their threads to the back of the rostellum, the rostellum bends a little forwards, and this partly draws the pollinia out of their anther-cells. The upper end of the anther consists of a blunt, solid point, not including pollen; this blunt point projects slightly beyond the face of the rostellum, which circumstance, as we shall see, is important.
The flowers stand out (Fig. A) horizontally from the stem. The labellum is curiously shaped, as may be seen in the drawings: the distal half, which projects beyond the other petals and forms an excellent landing-place for insects, is joined to the basal half by a narrow hinge, and naturally (Fig. A) is turned a little upwards, so that its edges pass within the
edges of the basal portion. So flexible and elastic is the hinge that the weight of even a fly, as Mr. More informs me, depresses the distal portion; it is represented in Fig. B in this state; but when the weight is removed it instantly springs up to its former (Fig. A) and ordinary position, and with its curious medial ridges partly closes the entrance into the flower. The basal portion of the labellum forms a cup, which at the proper time is filled with nectar.
Now let us see how all the parts, which I have been obliged to describe in detail, act. When I first examined these flowers I was much perplexed: trying in the same way as I should have done with a true Orchis; I slightly pushed the protuberant rostellum downwards, and it was very easily ruptured; some of the viscid matter was withdrawn, but the pollinia remained in their cells. Reflecting on the structure of the flower, it occurred to me that an insect in entering to suck the nectar, from depressing the distal portion of the labellum, would not touch the rostellum; but that, when within the flower, from the springing up of the distal half of the labellum, it would be almost compelled to back out parallel to the
stigma by the higher part of the flower. I then brushed the rostellum lightly upwards and backwards with the end of a feather and other such objects; and it was pretty to see how easily the membranous cap of the rostellum came off, and how well, from its great elasticity, it fitted the object, whatever its shape might be, and how firmly it clung to it from the viscidity of its under surface. Together with the cap large masses of pollen, adhering by the threads, were necessarily withdrawn.
Nevertheless the pollen-masses were not nearly so cleanly removed as those which had been naturally removed by insects. I tried dozens of flowers, always with the same imperfect results. It then occurred to me that an insect in backing out of the flower would naturally push with some part of its body against the blunt and projecting upper end of the anther which overhangs the stigmatic surface. Accordingly I so held the brush that, whilst brushing upwards against the rostellum, I pushed against the blunt solid end of the anther (see Sect. C ); this at once eased the pollinia, and they were withdrawn in an entire state. At last I understood the mechanism of the flower.
The large anther stands nearly parallel and behind the stigma (Sect. C), so that the pollinia when withdrawn by an insect would naturally adhere to its body in a position fitted to strike, as soon as it visited another flower, the almost parallel stigmatic surface. Hence we have not here, or in any Neotteæ, that movement of depression so common with the pollinia of the Ophreæ. When an insect with the pollinia attached to its back or head enters another flower, the easy depression of the distal portion of the labellum probably plays an important part; for the pollen-masses are extremely friable, and, if struck against the tips of the petals in the act of entering, much of the pollen would be lost; but as it is, an open gangway is offered, and the viscid stigma, with its lower part protuberant, lying in front, is the first object against which the pollen-masses projecting forwards from the insect's back would naturally strike.* I did not count the flowers,
* As it is quite possible that I may have overrated the importance of the peculiar structure of the labellum, I asked Mr. A. G. More to remove the distal half of the labellum from some flowers before they had expanded, but I was too late in my application. He was able to try only two flowers, which were situated near the summit of the spike. These flowers
but in one lot of spikes sent me by Mr. More a large majority of the pollinia had been naturally and cleanly removed by some unknown insect.
Epipactis latifolia.—This species agrees with the last in all the foregoing specified details, excepting that the rostellum projects considerably further beyond the face of the stigma, and the blunt upper end of the anther projects less. The viscid matter lining the elastic cap of the rostellum takes a longer time in setting dry. The upper petals and sepals are more widely expanded than in E. palustris: the distal portion of the labellum is smaller, and is firmly united to the basal portion (Fig. XIV.); so that it is not flexible and elastic: it apparently serves only as a landing-place for insects. The fertilisation of this species depends simply on an insect striking in an upward and backward direction the highly-protuberant rostellum, which it would be apt to do in
formed seed-capsules, which were certainly small; but this may have been owing to their position. Unfortunately also these capsules shed most of their seed in being sent to me; so that I could not ascertain whether the seeds were well formed. Of the few seeds which did remain within these two capsules many were shrivelled and bad.
retreating after having sucked the copious nectar in the cup of the labellum. Apparently it is not at all necessary that the insect should push back the less protuberant blunt upper end of the anther; at least I found that the pollinia could be easily removed by simply dragging off the cap of the rostellum in an upward and backward direction.
Fig. XIV.
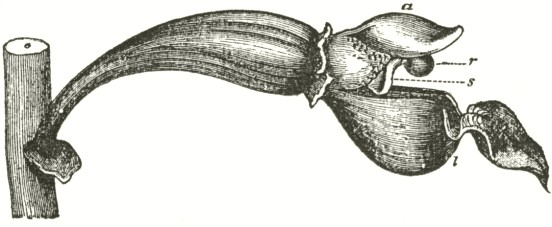
EPIPACTIS LATIFOLIA.
Flower viewed sideways, with all the sepals and petals removed, except the labellum.
a. anther.
r. rostellum.
s. stigma.
l. labellum.
In Germany C. K. Sprengel caught a fly with the pollinia of this species attached to its back. In England the flowers are much visited by insects: during the wet and cold season of 1860 a friend in Sussex examined five spikes bearing eighty-five expanded flowers: of these, fifty-three had the pollinia removed, and thirty-
two had them in place; but as many of the latter were immediately beneath the buds, ultimately a larger proportion would almost certainly have been renewed. In Devonshire I found a spike with nine open flowers, and the pollinia in all were removed with one exception, in which a fly, too small to remove the pollinia, had become glued to it and to the stigma, and had there miserably perished.
Cephalanthera grandiflora.—This Orchid appears to be closely allied to Epipactis, though it has been ranked by some authors in a widely different position. The stigma holds the same relative position to the anther as in Epipactis; but we have here the unique case (my remarks never apply to the very different group of the Cypripediæ) of there being no rostellum. The anther resembles that of Epipactis, but stands rather higher up relative to the stigma. The pollen is extremely friable, and readily adheres to any object rubbed against it; the spherical grains are separate, instead of being united by threes or fours, as in all other Orchids;* they are tied together by a few
* This separation of the grains was observed, and is represented, by Bauer in the plate published by Lindley in his magnificent 'Illustrations of Orchidaceous Plants.'
Fig. XV.
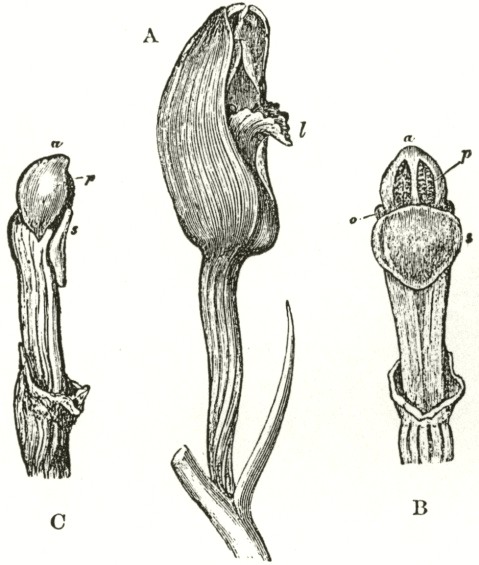
CEPHALANTHERA GRANDIFLORA.
a. anther; in the front view, B, the two cells with the pollen are seen.
o. rudimentary anther, or auricle.
p. masses of pollen.
s. stigma.
l. distal portion of the labellum.
A. Oblique view of perfect flower, when fully expanded.
B. Front view of column, with all the sepals and petals removed.
C. Side view of column, with all the sepals and petals removed; the narrow pillars of pollen between the anther and stigma can just be seen.
F3
weak elastic threads; so that in the state of the pollen, as well as in the abortion of the rostellum, we have degradation of structure. The anther opens whilst the flower is in bud and partly expels the pollen, which stands in two almost free upright pillars, each nearly divided longitudinally into two halves. These subdivided pillars rest in front against the upper square edge of the stigma, which rises to about one-third of their height (see front view B and side view C).
Whilst the flower is still in bud, or before it is as fully open as ever it becomes, the pollen-grains which rest against the upper sharp edge of the stigma (but not those in the upper or lower parts of the mass) emit a multitude of tubes, deeply penetrating the stigmatic tissue. After this period the stigma bends a little forward, and the result is that the two friable pillars of pollen almost stand free from their anther-cells, being tied to and supported in front by the penetration of their pollen-tubes into the edge of the stigma. Without this support the pillars would soon fall down.
Differently from Epipactis, the flower stands upright; the lower part of the labellum is
turned up parallel to the labellum (Fig. A), and the tips of the lateral petals never become quite separate;* so that the pillars of pollen are protected from the wind, and as the flower stands upright they do not tumble down from their weight. These are points of high importance to the plant, as the pollen would otherwise be blown or fall down and be wasted. The labellum is formed of two portions, as in Epipactis; and when the flower is mature, the small triangular distal portion turns down at right angles to the basal portion; thus forming a small landing-place in front of a triangular door, situated half-way up the almost tubular flower, by which insects can enter. I did not observe any nectar; but as the lower cup-formed portion of the labellum presents the same structure as in Epipactis, I presume it is secreted. After a short time, as soon as the flower is fully fertilised, the small distal portion of the labellum rises up, shuts the triangular door, and again perfectly encloses the organs of fructification.
* Bauer figures the flowers much more widely expanded: all that I can say is that I have not seen them in this condition.
In the early penetration of the stigma by a multitude of pollen-tubes, which I traced far down the stigmatic tissue, we apparently have another case, like that of the Bee Ophrys, of perpetual self-fertilisation. I was much surprised at this fact, and asked myself: Why does the distal portion of the labellum open for a short period? what is the use of the great mass of pollen above and below the layer of grains, the tubes of which alone penetrate the upper edge of the stigma? The stigma has a large flat viscid surface; and during several years I have almost invariably found masses of pollen adhering to its surface, and the friable pillars of pollen by some means broken down. It occurred to me, that although the flowers stand upright, and the pillars are well protected from the wind, yet that the pollen-masses might ultimately topple over from their own weight, and so fall on the stigma, thus completing the act of self-fertilisation. Accordingly, I covered up with a net a plant with four buds, and examined the flowers as soon as they had withered; the broad stigmas of three of these flowers were perfectly free from pollen, but
a little had fallen on one corner of the fourth. With the exception, also, of the summit of the pillar of pollen in this latter flower, all the other pillars still stood upright and unbroken. I looked at the flowers of some surrounding plants, and everywhere found, as I had so often done before, broken-down pillars and masses of pollen on the stigmas.
Hence we may safely infer that insects visit the flowers, disturb the pollen, and leave masses of it on the stigmas. We thus see that the reflexion of the distal portion of the labellum, by which a temporary landing-place and door are formed,—the upturned labellum, by which the flower is made tubular and insects are compelled to crawl close by the stigmatic surface,—the pollen readily cohering to any object, and standing in friable pillars protected from the wind,—and, lastly, the large masses of pollen standing above and below that layer of pollen-grains, of which alone the tubes penetrate the edge of the stigma,—are all co-ordinated structures, far from useless; and useless they would be if these flowers were capable of perfect self-fertilisation.
To ascertain how far the early and invariable penetration of the upper edge of the stigma by the pollen-grains, which rest on it, is effectual for fertilisation, I covered up a plant, just before the flowers opened, and removed the thin net as soon as they had begun to wither. From long experience I am sure that this temporary covering could not have injured their fertility. The four covered flowers produced as fine seed-capsules as any on the surrounding plants. When ripe, I gathered them, and likewise capsules from several surrounding plants, growing under similar conditions, and weighed the seed in a chemical balance. The seed from the four capsules from the uncovered plants weighed 1.5 grain, from the covered plant the seed of an equal number of capsules weighed under one grain; but this does not give a fair idea of the relative difference of fertility, for I observed that a great number of the seeds from the covered plant were mere minute and shrivelled husks. Accordingly I mixed the seeds well together, and took four little lots from one heap and four little lots from the other heap, and, having soaked them in
water, compared them under the compound microscope: out of forty seeds from the uncovered plants there were only four bad, whereas of forty seeds from the covered-up plants there were at least twenty-seven bad; so that there were nearly seven times as many bad seeds in the covered-up plants as in those left to the free accsess of insects.
Hence we have the following curious and complex case. Perpetual self-fertilisation, but in an extremely imperfect degree, by the early penetration of the pollen-tubes; this would be useful to the plants, if insects failed to visit the flowers. The penetration of the pollen-tubes, however, is apparently more important in retaining the pillars of pollen in their proper places, so that the insects, in crawling into the flowers, might get dusted with pollen. This imperfect self-fertilisation is habitually and largely aided by insects, which may carry a flower's own pollen on to its stigma; but an insect thus smeared with pollen could not fail likewise to cross distinct individuals. From the relative position of the parts, it seems indeed probable (but I omitted to prove this by the early
removal of the anthers, so as to observe whether pollen was brought to the stigma from other flowers) that an insect would more frequently get dusted by crawling out of a flower than by crawling into one; and this would of course facilitate a union between two distinct individuals. Hence Cephalanthera offers only a partial exception to the almost universal rule that the flowers of Orchids are fertilised by the pollen of other flowers.
Goodyera repens.*—This genus, in most of its characters with which we are concerned, is rather closely related to Epipactis. The shield-like rostellum is almost square, and projects beyond the stigma; it is supported on each side by sloping sides rising from the upper edge of the stigma, in nearly the same manner as we shall presently see in Spiranthes. The surface of the protuberant part of the rostellum is rough, and when dry can be seen to be formed of cells; it is delicate, and, when slightly pricked, exudes a little milky viscid fluid; it is lined by a layer of very adhesive matter, which
* Specimens of this rare Highland Orchid were most kindly sent me by the Rev. G. Gordon of Elgin.
quickly sets hard when exposed to the air. The protuberant surface of the rostellum, when gently rubbed upwards, is easily removed, and carries with it a strip of membrane, to the hind part of which the pollinia are attached. The sloping sides which support the rostellum are not removed at the same time, but remain projecting up like a fork and soon wither. The anther is borne on an elongated broad filament; a membrane on both sides unites this filament to the edges of the stigma, forming an imperfect cup or clinandrum. The anther-cells open in the bud, and the pollinia become attached by their anterior faces, just beneath their summits, to the back of the rostellum; ultimately the anther opens widely, leaving the pollinia almost naked, but partially protected within the membranous cup or clinandrum. Each pollinium is partially divided length-ways; the pollen-grains cohere in subtriangular packets, including a multitude of grains, each grain consisting of four granules; and these packets are tied together by strong elastic threads, which at their upper ends run together and form a single flattened brown elastic ribbon, of which the truncated extremity adheres to the back of the rostellum.
The surface of the orbicular stigma is remarkably viscid, which is necessary in order that the unusually strong threads connecting the packets of pollen should be ruptured. The labellum is partially divided into two portions; the terminal portion is reflexed, and the basal portion is cup-formed and filled with nectar. The passage into the flower between the rostellum and labellum is contracted. Since my examination of Spiranthes, immediately to be described, I have suspected that the labellum moves further from the column in mature flowers, in order to allow insects, with the pollinia adhering to their heads or probosces, to enter the flowers more freely. In many of the specimens received, the pollinia had been removed by insects, and the fork-shaped supporting sides of the rostellum were partially withered.
Goodyera is an interesting connecting link between several very distinct forms. In no other member of the Neotteæ have I seen so near an approach to the formation of a true caudicle,* like that in the Ophreæ; and it
* In a foreign species, the Goodyera discolor, sent me by Mr. Bateman, the pollinia approach in structure still closer to those of the Ophreæ; for the pollinia thin out into long caudicles, closely resembling in form those of an Orchis. The caudicle is
is curious that in this genus alone (as far as I have seen) the pollen-grains cohere in large packets, as in the Ophreæ. If the nascent caudicles had been attached to the lower ends of the pollinia, and they are attached a little beneath the summit, the pollinia would have been almost identical with those of a true Orchis. In the rostellum being supported by sloping sides, which wither when the viscid disc is removed,—in the membranous cup or clinandrum between the stigma and anther,—and in some other respects, we have a clear affinity to Spiranthes. In the anther having a broad filament we see a relation to Cephalanthera. In the structure of the rostellum, with the exception of the sloping sides, and in the shape of the labellum, we see the affinity of Goodyera to Epipactis. Goodyera probably shows us the state of the organs of fructifica-
formed of a bundle of elastic threads, with very small and thin packets of pollen-grains attached to them, and arranged like tiles one over the other. The two caudicles are united together near their bases, where they are attached to a disc of membrane lined with viscid matter. From the small size and extreme thinness of the basal packets of pollen, and from the strength of their attachment to the threads, I believe that they are in a functionless condition; if so, these prolongations of the pollinia are true caudicles.
tion in a large group of Orchids, now mostly extinct, but the parents of many living descendants.
Spiranthes autumnalis.—This Orchid, with its pretty name of Ladies'-tresses, presents some interesting peculiarities.* The rostellum is a long, thin, flat, projection, joined by sloping shoulders to the summit of the stigma. In the middle of the rostellum a narrow vertical brown object (Fig. XVI., C) may be seen, bordered on each side and covered by transparent membrane. This brown object I will call "the boat-formed disc." It forms the middle portion of the posterior surface of the rostellum, and consists of a narrow strip of exterior membrane in a modified condition. Its summit (Fig. E) is pointed, its lower end rounded, and it is slightly bowed, so as altogether to resemble a boat or canoe. It is rather more than 4/100 of an inch in length, and less than 1/100 in breadth. It is nearly rigid, and appears fibrous, but is really formed of elongated and thickened cells, partially confluent.
* I am much indebted to Dr. Battersby of Torquay, and to Mr. A. G. More of Bembridge, for sending me specimens; but I have subsequently examined many growing plants.
Fig. XVI.
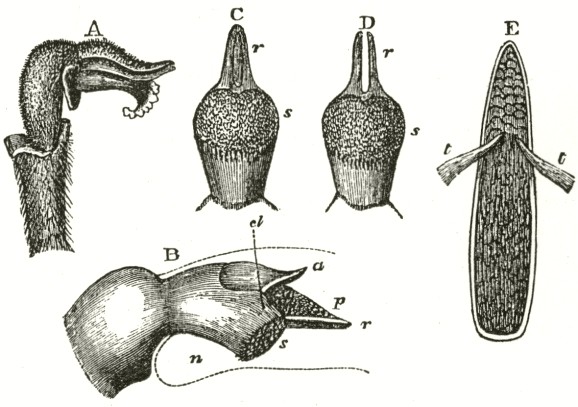
SPIRANTHES AUTUMNALIS, OR LADIES' TRESSES.
a. anther.
p. pollen-grains.
t. threads of the pollen-masses.
cl. margin of clinandrum.
r. rostellum.
s. stigma.
n. nectar-receptacle.
A. Side view of flower in its natural position, with the two lower sepals alone removed. The labellum can be recognised by its fringed and reflexed lip.
B. Side view of a mature flower, with all the sepals and petals removed. The position of the labellum (which has moved from the rostellum) and the upper sepal is shown by the dotted lines.
C. Front view of the stigma, and of the rostellum with its embedded central disc.
D. Front view of the stigma and of the rostellum after the viscid disc has been removed.
E. Viscid disc, removed from the rostellum, greatly magnified, viewed posteriorly, and with the attached elastic threads of the pollen-masses; the pollen-grains have been removed from the threads.
This boat, standing vertically up on its stern, is filled with thick, milky, extremely adhesive fluid, which, when exposed to the air, rapidly turns brown, and in about one minute sets quite hard. An object is well glued to the boat in four or five seconds, and when the cement is dry the attachment is wonderfully strong. The transparent sides of the rostellum, on each side of the disc, consist of membrane attached behind to the edges of the boat, and folded over in front, so as to form the anterior face of the rostellum. This folded membrane, therefore, covers, almost like a deck, the cargo of viscid matter within the boat.
The anterior face of the rostellum is slightly furrowed in a longitudinal line over the middle of the boat, and is endowed with a remarkable vital property; for, if the furrow be touched very gently by a needle, or if a bristle be laid along the furrow, it instantly splits along its whole length, and a little milky adhesive fluid exudes. This action is not mechanical, or due to simple violence. The fissure runs up the whole length of the rostellum, from the stigma beneath to the summit: at the summit the fissure bifurcates,
and runs down the back of the rostellum on each side and round the stern of the boat-formed disc. Hence after this splitting action the boat-formed disc lies quite free, but embedded in a fork in the rostellum. The act of splitting apparently never takes place spontaneously. I covered up under a net a plant with unexpanded flowers, and five of these flowers remained fully expanded for exactly a week under the net: I then examined their rostella, and not one had split; whereas almost every flower on the surrounded and uncovered spikes, which had been visited and touched by insects, after remaining expanded for only twenty-four hours, had their rostella fissured. Exposure for two minutes to the vapour of a little chloroform causes the rostellum to split; and this we shall hereafter see is likewise the case with some other Orchids.
When a bristle is laid for two or three seconds in the furrow of the rostellum, and the membrane has consequently become fissured, the viscid matter within the boat-formed disc lies so close to the surface, and, indeed, slightly exudes, that the disc is almost sure to be glued longitudinally to the bristle and
to be withdrawn with it. When the disc is withdrawn the two sides of the rostellum (Fig. D), which have been described by some botanists as two distinct foliaceous projections, are left sticking up like a fork. This is the common condition of the flowers after they have been open for two or three days, and have been visited by insects. The fork soon withers.
When the flower is in bud the back of the boat-formed disc is covered with a layer of large rounded cells, so that the disc does not strictly form the exterior surface of the back of the rostellum. These cells contain slightly viscid matter: they remain unaltered (as may be seen at Fig. E) towards the apex of the disc, but at the point where the pollinia are attached they disappear. Hence I concluded that the viscid matter contained in these cells, when they burst, served to fasten the threads of the pollinia to the disc; but, as in several large exotic Orchids I could see no trace of such cells, I presume that this view is erroneous.
The stigma lies beneath the rostellum, and projects with a sloping surface (see side-view,
B); its lower margin is rounded and fringed with hairs. On each side a membrane (cl, B) extends from the edges of the stigma to the filament of the anther, thus forming a membranous cup or clinandrum, in which the lower ends of the pollen-masses lie protected.
Each pollinium consists of two leaves of pollen, quite disconnected at their lower ends, and with their summits distinct, but they are united by elastic threads for about half of their length: a very slight modification would convert the two pollinia into four leaves of pollen, as occurs in the genus Malaxis and in many foreign Orchids. Each of the four leaves, moreover, consists of a double layer of pollen-grains, united only along their edges. The pollen-grains (each consisting of four granules) are united by elastic threads, which are more numerous along the edges of the leaves, and converge at the summit of each pollinium. The leaves of pollen are very brittle, and, when placed on the viscid stigma of a flower, large sheets are easily broken off.
Long before the flower expands, the anther-cells, which are pressed against the back of the rostellum, open in their upper part, so that
the included pollinia come exactly into contact with the back of the boat-formed disc; the projecting threads then become firmly attached to rather above the middle part of the back of the disc. The anther-cells afterwards open lower down, and their membranous walls contract and become brown; so that by the time the flower has fully expanded the upper parts of the pollinia lie quite naked, their bases rest in little cups formed by the withered anther-cells, and they are laterally protected by the clinandrum. As the pollinia thus lie loose, they are easily removed.
The tubular flowers are elegantly arranged in a spire round the spike, extending horizontally (Fig. A) from it. The labellum is channelled down the middle, and is furnished with a reflexed and fringed lip, on which bees alight; its basal internal angles are produced into two globular processes, which secrete an abundance of nectar. The nectar is collected ( n, Fig. B) in a small receptacle beneath. Owing to the protuberance of the lower margin of the stigma, and of the two lateral inflexed nectaries, the orifice into the nectar-receptacle is much contracted, and is central.
When the flower first opens the receptacle contains nectar, and at this period the front of the rostellum, which is slightly furrowed, lies close to the channelled labellum; consequently a passage is left, but so narrow that only a fine bristle can be passed down it.* In a day or two the labellum moves a little farther from the rostellum, and a wider passage is left to the stigmatic surface. On this slight movement of the labellum the fertilisation of the flower absolutely depends.
With most Orchids the flowers remain open for some time before they are visited by insects; but with Spiranthes I have generally found the boat-formed disc removed very soon after the expansion of the flower. For example, in the two last spikes which I happened to examine there were in one numerous buds on the summit, with the seven lowest flowers alone expanded, of which six had their discs and pollinia removed; the other spike had eigth expanded flowers, with the pollinia of all removed. We have seen
* Professor Asa Gray was so kind as to examine Spiranthes gracilis and cernua in the United States. He found the same general structure as in our S. autumnalis, and he was struck with the narrowness of the passage into the flower.
G2
that when the flower first opens it would be attractive to insects, for the receptacle already contains nectar; and the rostellum at this period lies so close to the channelled labellum that a bee or moth could not pass down its proboscis without touching the medial furrow of the rostellum. This I know to be the case by repeated trials with a bristle.
Let it be observed how beautifully everything is contrived that the pollinia should be withdrawn by an insect visiting the flower. The pollinia are already attached to the disc by their threads, and, from the early withering of the anther-cells, they hang loosely suspended but protected within the clinandrum. The touch of the proboscis causes the rostellum to split in front and behind, and frees the long, narrow, boat-formed disc, which is loaded with extremely viscid matter, sure to adhere longitudinally to the proboscis. When the bee flies away, so surely will it carry away the pollinia. As the pollinia are attached parallel to the disc, they will become attached parallel to the proboscis. Here, however, appears a difficulty; when the flower first opens and is best adapted for the removal of
the pollinia, the labellum lies so close to the rostellum, that the pollinia, when attached to the proboscis of an insect, could not possibly be forced into the flower so as to reach the stigma; they would be either upturned or broken off; but we have seen that after two or three days the labellum becomes more reflexed and moves from the rostellum,—a wider passage being thus left. When in this condition, I have tried with the pollinia attached to a fine bristle; and by inserting the bristle into the nectar-receptacle (n, Fig. B), it is pretty to see how sheets of pollen are left adhering to the viscid stigma. It may be observed in the diagram, B, that the orifice into the nectar-receptacle lies, owing to the projection of the stigma, close to the lower side of the flower; hence insects would insert their probosces on this side, and an open space is thus afforded for the attached pollinia to be carried down to the stigma, without being brushed off. The stigma evidently projects so that the ends of the pollinia may strike against it.
Hence, in Spiranthes, not only the pollen must be carried from one flower to another, as
in most Orchids, but a lately expanded flower, which has its pollinia in the best state for removal, cannot then be fertilised. Generally old flowers will be fertilised by the pollen of younger flowers, borne, as we shall see, on a separate plant. In conformity with this I observed that the stigmatic surfaces of the older flowers were far more viscid than those of the younger flowers. Nevertheless, a flower which in its early state had not been visited by insects would not necessarily, in its later and more expanded condition, have its pollen wasted; for insects, in inserting and withdrawing their probosces, bow them forwards, and would thus often strike the furrow of the rostellum. I imitated this action with a bristle, and often succeeded in withdrawing the pollinia: I was led to try this from at first choosing older flowers for examination; and passing a bristle, or fine culm of grass, straight down into the nectary, the pollinia were never withdrawn; but when I bowed it forward, I succeeded. Those flowers which have not their own pollinia removed can of course be fertilised, and I have seen not a few cases of flowers with their pollinia still
in place, with sheets of pollen on their stigmas.
At Torquay I watched a number of these flowers growing together for about half an hour, and saw three humble-bees of two kinds visit them. I caught one and examined its proboscis: on the upper lamina, some little way from the tip, two perfect pollinia were attached, and three other boat-formed discs without pollen; so that this bee had removed the pollinia from five flowers, and had probably left the pollen of three of them on the stigmas of other flowers. The next day I watched the same flowers for a quarter of an hour, and caught another humble-bee at work; one perfect pollinium and four boat-formed discs adhered to its proboscis, one on the top of the other, showing how exactly the same part had each time touched the rostellum.
The bees always alighted at the bottom of the spike, and, crawling spirally up it, sucked one flower after the other. I believe humble-bees generally act thus when visiting a dense spike of flowers, as it is most convenient for them; in the same manner as a woodpecker
always climbs up a tree in search of insects. This seems a most insignificant observation; but see the result. In the early morning, when the bee starts on her rounds, let us suppose that she alighted on the summit of the spike; she would surely extract the pollinia from the uppermost and last opened flowers; but when visiting the next succeeding flower, of which the labellum in all probability would not as yet have moved from the column (for this is slowly and very gradually effected), the pollen-masses would often be brushed off her proboscis and be wasted, But nature suffers no such waste. The bee goes first to the lowest flower,
and, crawling spirally up the spike, effects nothing on the first spike which she visits till she reaches the upper flowers, then she withdraws the pollinia: she soon flies to another plant, and, alighting on the lowest and oldest flower, into which there will be a wide passage from the greater reflexion of the labellum, the pollinia will strike the protuberant stigma: if the stigma of the lowest flower has already been fully fertilised, little or no pollen will be left on its dried surface; but on the next succeeding flower, of which the stigma is viscid, large sheets of pollen will be left. Then as soon as the bee arrives near the summit of the spike she will again withdraw fresh pollinia, will fly to the lower flowers on another plant, and fertilise them; and thus, as she goes her rounds and adds to her store of honey, she will continually fertilise fresh flowers and perpetuate the race of our autumnal Spiranthes, which will yield honey to future generations of bees.
G3
CHAPTER IV.
Malaxis paludosa; simple means of fertilisation—Listera ovata; sensitiveness of the rostellum; explosion of viscid matter; action of insects; perfect adaptation of the several oegans—Listera cordata—Neottia nidus-avis; its fertilisation effected in the same manner as in Listera.
WE now come to the last of the British Orchids, those in which no portion of the exterior membranous surface of the rostellum is permanently attached to the pollinia. This subdivision includes only three genera known to me, namely, Malaxis, Listera and Neottia, which are here grouped together merely for convenience. Malaxis in its fertilisation is an uninteresting form; Listera and Neottia, on the other hand, are amongst the most remarkable of all Orchids from the manner in which their pollinia are removed by insects, through the sudden explosion of viscid matter contained within their rostellums.
Malaxis paludosa.—This rare Orchid,* the least of the British species, differs from the
* I am greatly indebted to Mr. Wallis, of Hartfield, in Sussex, for numerous living specimens of this Orchid.
others in the position of its flowers. In Malaxis the labellum is turned upwards,* instead of downwards, so as to afford, as in other Orchids, a landing-place for insects; its lower margin clasps the column, making the entrance into the flower tubular. From its position it partially protects the organs of fructification. [Fig. XVII.] In most Orchids the upper sepal and the two upper petals afford protection; but here they are reflexed in a singular manner (as may be seen in the drawing, Fig. A), apparently to allow insects freely to visit the flower. This position of the flower is the more remarkable, because it has been purposely acquired, as shown by the ovarium being spirally twisted. In all Orchids the labellum is properly directed upwards, but assumes its usual position as the lower lip by the twisting of the ovarium; but in Malaxis the twisting has been carried to that degree that the flower occupies the position which it should have held if the ovarium
* Sir James Smith, I believe, first noticed this fact in the 'English Flora,' vol. iv. p. 47, 1828. Towards the summit of the spike the upper sepal does not depend, as represented in the woodcut (Fig. A), but sticks out nearly at right angles; nor is the flower always so completely twisted around.
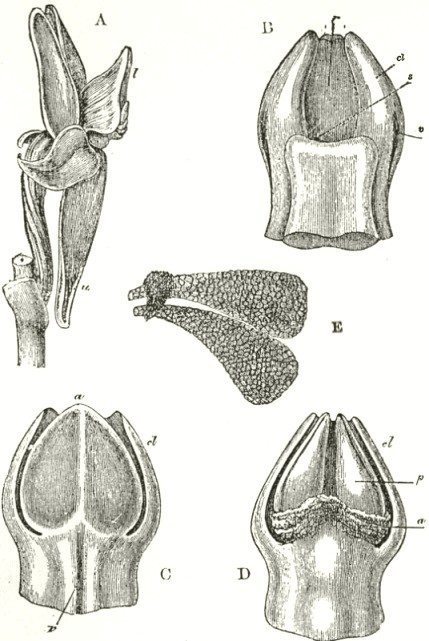
MALAXIS PALUDOSA.
(Partly copied from Bauer, but modified from living specimens.)
a. anther.
p. pollen.
cl. clinandrum.
v. spiral vessels.
r. rostellum.
s. stigma.
l. labellum.
u. upper sepal.
A. Perfect
had not been at all twisted, and which the ovarium ultimately assumes when ripe, by a process of gradual untwisting.
When the minute flower is dissected, the column is seen to be longitudinally tripartite; the middle portion of the upper half (see Fig. B) is the rostellum. The upper edge of the lower part of the column projects where united to the base of the rostellum, and forms a rather deep fold. This fold is the stigmatic cavity, and may be compared to a waistcoat-pocket. I found pollen-masses which had their broad ends pushed by insects into this pocket; and a bundle of pollen-tubes had here penetrated the stigmatic tissue.
The rostellum, or middle portion, is a tall membranous projection of a whitish colour,
_______
A. Perfect flower viewed laterally, with the labellum in its natural position, upwards.
B. Column viewed in front, showing the rostellum, the pocket-like stigma, and the anterior lateral portions of the clinandrum.
C. Back view of the column in a flower-bud, showing the anther with the pear-shaped included pollinia dimly seen, and the posterior edges of the clinandrum. D. Back view of an expanded flower, with the anther now contracted and shrivelled, exposing the pollinia.
E. The two pollinia attached to a little transverse mass of viscid matter, hardened by spirits of wine.
formed of square cells, and is covered with a thin layer of viscid matter: it is slightly concave posteriorly, and its crest is surmounted by y minute tongue-shaped mass of viscid matter. The column, with its narrow pocket-like stigma and with the rostellum above, is united on each side to a green membranous expansion, convex exteriorly and concave interiorly, of which the summits on each side are pointed, and stand a little above the crest of the rostellum. These two membranes sweep round (see back views, Figs. C and D), and are united to the filament or base of the anther; thus forming a deep cup or clinandrum behind the rostellum. The use of this cup is to afford protection, as we shall immediately see, to the pollen-masses. When I have to treat of the homologies of the different parts, it will be shown by the course of the spiral vessels that these two membranes, forming the clinandrum, consist of the two upper anthers of the inner whorl, in a rudimentary condition, but utilised for this special purpose.
In a flower before it expands, a little mass or drop of viscid fluid may be seen on the crest of the rostellum, rather overhanging its
front surface. After the flower has remained open for a little time, this drop shrinks and becomes more viscid. Its chemical nature is different from that of the viscid matter in most Orchids, for it remains fluid for many days, though fully exposed to the air. From these facts I concluded that the viscid fluid exuded from the crest of the rostellum; but fortunately I examined a closely-allied Indian form, namely, the Microstylis Rhedii (sent me from Kew by Dr. Hooker), and in this, before the flower opened, there was a similar drop of viscid matter; but on opening a still younger bud, I found a minute, regular, tongue-shaped projection on the crest of the rostellum, formed of cells, which when slightly disturbed resolved themselves into a drop of viscid matter. At this age, also, the front surface of the whole rostellum, between its crest and the pocket-like stigma, was coated with cells filled with similar brown viscid matter; so that there can be no doubt, had I examined a young enough bud of Malaxis, I should have found a similar minute tongue-shaped cellular projection on the crest of the rostellum.
The anther opens widely whilst the flower is in bud, and then shrivels and contracts downwards, so that, when the flower is fully expanded, the pollinia are quite naked, with the exception of their broad lower ends, which rest in two little cups formed by the shrivelled anther-cells. This contraction of the anther is represented in Fig. D in comparison with Fig. C , which shows the state of the anther in a bud. The upper and much pointed ends of the pollinia rest on, but project beyond, the crest of the rostellum; in the bud they are unattached, but by the time the flower opens they are always caught by the posterior surface of the drop of viscid matter, of which the anterior surface projects slightly beyond the face of the rostellum; that they are caught without any mechanical aid I ascertained by allowing some buds to open in my room. In Fig. E the pollinia are shown exactly as they appeared, but not quite in their natural position, when removed by a needle from a specimen kept in spirit of wine, in which the irregular little mass of viscid matter had become hardened and adhered firmly to their tips.
The pollinia consist of two pair of very
thin leaves of waxy pollen; these four leaves are formed of angular grains (each apparently subdivided into four granules), which never separate. As the pollinia are almost loose, being retained merely by their tips adhering to the viscid fluid, and by their bases resting in the shrivelled anther-cells, and as the petals and sepals are so much reflexed, the pollinia would be exposed in a remarkable degree when the flower is fully expanded, and would be liable to be blown out of their proper position, had it not been for the membranous expansions on each side of the column forming the clinandrum, within which they safely lie.
When an insect inserts its proboscis, or head, into the narrow space between the upright labellum and the rostellum, it will infallibly touch the little projecting viscid mass, and when it flies away it will withdraw the pollinia, already attached to the viscid matter, but otherwise loose. I easily imitated this action by inserting any small object into the tubular flower between the labellum and rostellum. When the insect visits another flower, the very thin pollen-leaves attached parallel to the proboscis, or head, will be
forced in, and their broad ends will enter the pocket-like stigma. I found pollinia in this position glued to the upper membranous expansion of the rostellum, and with a large number of pollen-tubes penetrating the stigmatic tissue. The use of the thin layer of viscid matter, which coats the surface of the rostellum in this genus and in Microstylis, and which is of no use in the transportal of the pollen from flower to flower, seems to be to keep the leaves of pollen, when brought by insects, in a proper position for entering and remaining in the narrow stigmatic cavity. This fact is rather interesting under a homological point of view, for, as we shall hereafter see, the primordial nature and purpose of the viscid matter of the rostellum is that common to the viscid matter on the stigmas of most flowers, namely, the retention of the pollen, when brought by any means to its surface.
The flowers of the Malaxis, though so small and inconspicuous, are highly attractive to insects; for the pollinia had been removed from all the flowers on each spike, excepting from one or two close under the buds. In
some old flower-spikes every single pollinium had been removed. Insects sometimes remove only one of the two pairs. I noticed one flower with all its four pollinia in place, with a single pollen-leaf within the stigmatic cavity; and this must clearly have been brought by some insect. I found pollen-leaves on the stigmas of many other flowers. The plant produces plenty of seed: on one spike, thirteen of the twenty-one lower flowers had formed large capsules.
Listera ovata, or Tway-blade.—The structure and action of the rostellum of this Orchid has been the subject of a highly remarkable paper in the Philosophical Transactions, by Dr. Hooker,* who has described minutely, and of course correctly, its curious structure; he did not, however, attend to the part which insects play in the fertilisation of this flower. C. K. Sprengel well knew the importance of insect-agency, but he misunderstood both the structure and the action of the rostellum.
The rostellum is of large size, thin, or foliaceous; convex in front and concave be-
* 'Philosophial Transactions,' 1854, p. 259.
hind, with its sharp summit slightly hollowed out on each side; it arches over the stigmatic surface (Fig. XVIII. r. s. A). Internally, according to Dr. Hooker, it is divided by longitudinal septa into a series of loculi, which contain and subsequently expel with violence viscid matter. These loculi show traces of their original cellular structure. I have met with this structure of the rostellum in no other genus excepting the closely allied Neottia. The anther, situated behind the rostellum, and protected by a broad expansion of the top of the column, opens in the bud. The pollinia, when the flower is fully expanded, are left quite free, supported behind by the anther-cells, and lying in front against the concave back of the rostellum, with their upper and pointed ends resting on its crest. Each pollinium is almost divided into two masses. The pollen-grains are attached together in the usual manner by a few elastic threads; but the threads are weak, and large masses of pollen can be easily broken off. After the flower has long remained open, the pollen becomes more friable. The labellum is much elongated, contracted at the
Fig. XVIII.
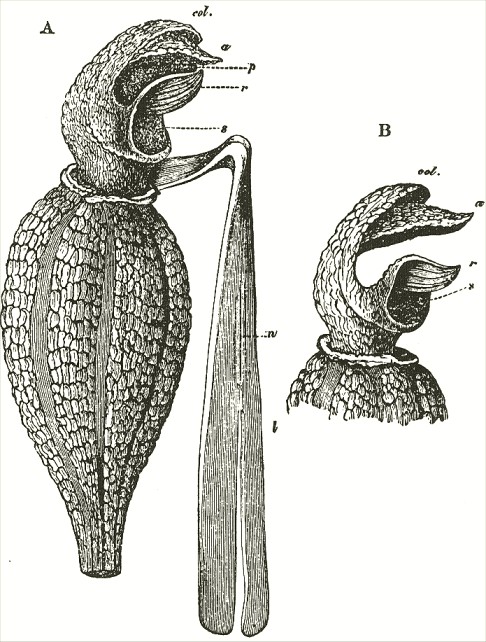
LISTERA OVATA, or Tway-blade (partly copied from Hooker).
col. summit of column.
a. anther.
p. pollen.
r. rostellum.
s. stigma.
l. labellum.
n. nectar-secreting furrow.
A. Flower viewed laterally, with all the sepals and petals, except the labellum, removed.
B. Ditto, with the pollinia removed, and with the rostellum more reflexed after its explosion.
base, and bent downwards, as represented in the drawing; it is furrowed along the middle, from its bifurcation close up to the base of the stigma; and the borders of the furrow are glandular and secrete much nectar.
As soon as the flower opens, if the crest of the rostellum be touched ever so lightly, a large drop of viscid fluid is instantaneously expelled; this, as Dr. Hooker has shown, is formed by the coalescence of two drops proceeding from the two depressed spaces on each side of the crest. A good proof of this fact was afforded by some specimens kept in weak spirits of wine, which had apparently expelled the viscid matter slowly, and in which two separate little spherical balls of viscid matter had hardened and had become attached to the two pollinia. The fluid is at first slightly opaque and milky; but as soon as exposed to the air, in less time than a second, a film is formed over it, and in two or three seconds the whole drop sets hard, and soon assumes a purplish-brown tint. So exquisitely sensitive is the rostellum, that a touch from the thinnest human hair suffices to cause the explosion. It will take place
under water. Exposure to the vapour of chloroform for about one minute also causes the explosion. The viscid fluid when pressed between two plates of glass before it has set hard is seen to be structureless; but it assumes a reticulated appearance, perhaps caused by the presence and union of globules of a denser fluid immersed in a thinner fluid. As the pointed tips of the loose pollinia lie on the crest of the rostellum, they are always caught by the exploded drop: I have never once seen this fail. So rapid is the explosion and so viscid the fluid, that it is difficult to touch the rostellum with a needle quickly enough not to catch the pollinia already attached to the partially-hardened drop. Hence, if a bunch of flowers be carried home in the hand, some of the sepals and petals, being shaken, will be almost sure to touch the rostellum and withdraw the pollinia, which will cause the false appearance of their having been violently ejected to a distance.
After the anther-cells have opened and have left the naked pollinia resting on the concave back of the rostellum, the rostellum curves a little forwards, and perhaps the
anther also moves a little backwards. This movement is of much importance, as the tips of the anther would otherwise be caught by the exploded viscid matter, and the pollinia would for ever be locked up and rendered useless. I once found an injured flower which had been pressed and had exploded before fully expanding, and the anther, with the enclosed pollen-masses, was permanently glued down to the crest of the rostellum. At the moment of the explosion the rostellum, which already stands arched over the stigma, quickly bends forwards and downwards as as to stand (Fig. B) at right angles to its surface. The pollinia, if not removed by the touching object that causes the explosion, become fixed to the rostellum, and by its movement are drawn a little forward. If their lower ends are now freed by a needle from the anther-cells, they spring up; but they are not by this movement placed on the stigma. In the course of some hours, or of a day, the rostellum not only slowly recovers its original slightly-arched position, but becomes quite straight and parallel to the stigmatic surface. This backward movement of the rostellum is
of importance; for if after the explosion it permanently remained projecting at right angles close over the stigma, pollen could with difficultly have been left on its viscid surface. When the rostellum is touched so quickly that the pollinia are not removed, they are, as I have said, drawn at the moment of explosion a little forward by the movement of th rostellum; but by its subsequent backward movement the pollinia are pushed back into their original position.
From the account here given we may safely infer how the fertilisation of this Orchid is effected. Small insects alight on the broad lower end of the labellum for the sake of the nectar copiously secreted by it; as they lick up the nectar they slowly crawl up its narrowed surface until their heads stand directly under the over-arched crest of the rostellum; as they raise their heads they touch the crest, which explodes, and the pollinia become firmly cemented to them. The insect in flying away withdraws the pollinia, carries them to another flower, and leaves masses of the friable pollen on its viscid stigma.
In order to witness what I felt sure would take place, I watched a group of plants on two or three occasions for an hour; each day I saw numerous specimens of two small Hymenopterous insects, namely, a Hæmiteles and a Cryptus, flying about the plants and licking up the nectar; most of the flowers, which were visited over and over again, had already had their pollinia removed, but at last I saw both these insect-species crawl into younger flowers, and suddenly retreat with a pair of bright yellow pollinia sticking to their foreheads; I caught them, and found the point of attachment was to the inner edge of the eye; on the other eye of one specimen there was a ball of the hardened viscid matter, showing that it had previously removed another pair of pollinia, and had subsequently in all probability left them on the stigma of one of the flowers. As I caught these insects, I did not witness the act of fertilisation; but C. K. Sprengel actually saw a Hymenopterous insect leave its pollen-mass on the stigma. My son watched another bed of this Orchid at some miles' distance, and brought me home the same Hymenopterous insects with attached
pollinia, and he saw Diptera also visiting the flowers. He was struck with the number of spider-webs spread over these plants, as if the spiders were aware how attractive the Listera was to insects, and how necessary they were to its fertilisation.
To show how delicate a touch suffices to cause the rostellum to explode, I may mention that I found an extremely minute Hymenopterous insect vainly struggling with its whole head burried in the hardened viscid matter, thus cemented to the crest of the rostellum and to the tips of the pollinia; the insect was not so large as one of the pollinia, and after causing the explosion it had not force to remove them, and was thus punished for attempting a work beyond its strength, and perished miserably.
In Spiranthes the young flowers, which have their pollinia in the best state for removal, cannot possibly be fertilised; they must remain in a virgin condition until they are a little older and the labellum has moved from the column. Here the same thing apparently occurs, for the stigmas of the older flowers were more viscid than those of
the younger flowers. These latter have their pollinia quite ready for removal; but immediately after the explosion the rostellum, as we have seen, curls forwards and downwards, thus protecting the stigma for a time, until the rostellum slowly becomes quite straight, when the stigma in its more mature condition is left freely exposed, and is ready for fertilisation.
I was curious to ascertain whether the rostellum would ultimately explode, if never touched; but I have found it difficult to ascertain this fact, as the flowers are so attractive to insects, and the touch of such minute insects suffices to cause the explosion, that it was scarcely possible to exclude them.
I have covered up plants several times, and left them till long after all uncovered plants had set their pods; and without going into unnecessary details, I may positively state that the rostella in several flowers had not exploded, though the stigma was withered, the pollen quite mouldy and incapable of removal. Some few of the very old flowers, however, when roughly touched, were still capable of feeble explosion. Other flowers
under the nets had exploded, and the tips of their pollinia were fixed to the crest of the rostellum; but whether these had been touched by some excessively minute insect, or had exploded spontaneously, it is impossible to say. In the case of these latter flowers, it is of some importance to note that not a grain of pollen had got on their stigmas (and I looked carefully), and their ovaria had not swollen. These several facts clearly show that the removal of the pollinia by insect-agency is absolutely necessary to the fertilisation of this species.
That insects do their work effectually, the following cases show:—A rather young spike, with many buds in the upper part, had the pollinia unremoved from the seven upper flowers, but they had been completely removed from the ten lower flowers: there was pollen on the stigmas of six of these ten lower flowers. In two spikes taken together, the twenty-seven lower flowers had their pollinia removed, with pollen on the stigmas of all; these were
succeeded by five open flowers with pollinia not removed, and with no pollen on their stigmas; and these were succeeded by eighteen buds. Lastly, in an older spike with forty-four flowers, all fully expanded, the pollinia had been removed from every one; and on all the stigmas that I examined there was pollen, generally in large quantity.
It will perhaps be worth while to recapitulate the several special adaptations for the fertilisation of this Orchid. The anther-cells open early, leaving the pollen-masses quite loose, with their tips resting on the concave crest of the rostellum. The rostellum then slowly curves over the stigmatic surface, so that its explosive crest stands at a little distance from the anther; and this is very necessary, otherwise the anther would be caught by the viscid matter, and the pollen for ever locked up. This curvature of the rostellum over the stigma and base of the labellum is excellently well adapted to favour an insect striking the crest when it raises its head, after having crawled up the labellum, and licked up the last drop of nectar at its base. The labellum, as C. K. Sprengel has remarked, becomes narrower where it joins the column beneath the rostellum, so
that there is no risk of the insect going too much to either side. The crest of the rostellum is so exquisitely sensitive, that a touch from a minute insect causes it to rupture at two points, and instantaneously two drops of viscid fluid are expelled, which coalesce. This viscid fluid sets hard in so wonderfully rapid a manner that it rarely fails to cement the tips of the pollinia, nicely laid on the crest of the rostellum, to the insect's forehead. As soon as the rostellum has exploded it suddenly curves downward till it projects at right angles over the stigma, protecting it in its early state from impregnation, in the same manner as the stigma of Spiranthes is protected by the labellum clasping the column. But as in Spiranthes the labellum after a time moves from the column, leaving a free passage for the introduction of the pollinia, so here the rostellum moves back, and not only recovers its former arched position, but stands upright, leaving the stigmatic surface, now become more viscid, perfectly free for pollen to be left on it. The pollen-masses, when once cemented to an insect's forehead, will generally remain firmly attached to it
until the viscid stigma of a mature flower remove these encumbrances from the insect, by rupturing the weak elastic threads by which the grains are tied together—receiving at the same time the benefit of fertilisation.
Listera cordata.—Professor Dickie of Aberdeen was so kind as to send me two sets of specimens, but I applied rather too late in the season. The structure is essentially the same as in the last species, and the loculi of the rostellum were very plain. In the middle, on the crest of the rostellum, two or three little hairy points project; whether these have any functional importance I know not. The labellum has two basal lobes (of which vestiges may be seen in L. ovata) which curve up on each side, and would compel an insect to approach the rostellum straight in front. Two flowers had been touched either during the journey, or too quickly by some insect, and had exploded; and their pollinia in consequence were firmly cemented to the crest of the rostellum; but in most of the spikes all the pollen-masses had been renoved by insects.
Neottia nidus-avis.—I made numerous ob-
servations on this, the Bird's-nest Orchis,* but they are not worth giving, as the action and structure of every part is almost identically the same as in Listera ovata. The labellum secretes plenty of nectar, which I mention merely as a caution, because during one cold and wet season I looked several times and could not see a drop, and was perplexed at the apparent absence of any attraction for insects; nevertheless, had I looked more perseveringly, I should probably have found nectar.
Whether the rostellum ultimately explodes, if not touched, I could not ascertain; that it long remained unexploded, though ready to act, is certain; but I found in 1860 so many flowers exploded, with a bead of the purplish hardened cement attached to the crest of the rostellum and to the unremoved pollinia, that I suspect that it does explode, after a time, spontaneously, without the excitement of a touch. In one large spike, every flower had
* This unnatural sickly-looking plant has generally been supposed to be parasitic on the roots of the trees under the shade of which it lives; but, according to Irmisch ('Beiträge zur Biologie und Morphologie der Orchideen,' 1853, s. 25), this certainly is not the case.
been visited by insects, and all the pollinia had been removed. Another unusually fine spike from South Kent, sent me by Mr. Oxenden, had borne forty-one flowers, and produced twenty-seven large seed-capsules, besides some smaller ones.
The pollen resembles that of Listera, in consisting of grains (formed of four granules) tied together by a few weak threads; it differs in being much more incoherent, so that after a few days it swells and overhangs the sides and summit of the rostellum: hence, if the rostellum of a rather old flower be touched, and an explosion be caused, the pollinia are not so neatly caught by their tips as in Listera; consequently a good deal of the friable pollen is often left behind in the anther-cells and is apparently wasted. Portions of this fall on the corolla; and as the pollen in this state readily adheres to any object, it is not improbable that insects crawling about would thus get dusted, and leave some on the viscid stigma, without having touched the rostellum and caused it to explode. Certainly if the labellum were more upturned, so that insects were forced to
brush against the anther and column, they would get smeared with the pollen as soon as it had become friable, and might thus effectually fertilise the flower.
This observation interested me, because, when previously examining Cephalanthera, with its aborted rostellum, its upturned labellum, and its friable pollen, I had speculated how a transition, with each gradation useful to the plant, could possibly have been effected from the state of the pollen and flower in the allied Epipactis, with its pollinia attached to a well-developed rostellum, to the present condition of Cephalanthera. Neottia nidus-avis shows to a certain extend how such a transition might have been effected. This Orchid is at present mainly fertilised by means of the explosive rostellum, which acts effectually only as long as the pollen remains in mass; but unless we suppose that the circumstance of the pollen soon becoming friable is a mere injury to the plant, we may believe that the pollen in this condition is sometimes transported to the stigma by its adhesion to the hairy bodies of insects. If this be so, we can see that a slight change in the form of the
flower, and by the pollen becoming friable at a still earlier age, this means of fertilisation might be rendered more and more effectual, and the explosive rostellum less and less useful. Ultimately the rostellum would become a superfluity; and then, on the great principle of the economy of organisation, rendered so necessary by the struggle for live, by which every part of every being tends to be saved, the rostellum would be absorbed or aborted. In this case we should have the production of a new Orchid in the condition of Cephalanthera, as far as its means of fertilisation are concerned, but in general structure still closely allied to Neottia and Listera.
CHAPTER V.
Cattleya, simple manner of fertilisation—Masdevallia, curious closed flower—Dendrobium, contrivance for self-fertilisation—Vandeæ, diversified structure of the pollinia; importance of the elasticity of the pedicel; its power of movement—Elasticity and strength of the caudicle—Calanthe with lateral stigmas, manner of fertilisation—Angræcum sesquipedale, wonderful length of nectary—Acropera, perplexing case, a male Orchid.
HAVING examined the means of fertilisation in so many British Orchids, belonging to fourteen genera, I was anxious to ascertain whether the exotic forms, belonging to quite distinct Tribes, equally required insect-agency. I especially wished to ascertain whether rule holds that each flower is necessarily fertilised by pollen brought from a distinct flower; and in a secondary degree I was curious to know whether the pollinia underwent those curious movements of depression by which they are placed, after transportal by insects, in the proper position to strike the stigmatic surface.
By the kindness of many friends and strangers I have been enabled to examine fresh flowers of several species, belonging
to forty-three exotic genera, well dispersed through the sub-families of the vast Orchidean series.* It is not my intention to describe the means of fertilisation in all these genera, but merely to pick out a few curious cases, and other cases which illustrate the foregoing descriptions. The diversity of the contrivances, almost all adapted to favour the intercrossing of distinct flowers, seems to be exhaustless.
Mr. James Veitch, jun., has generously given me many beautiful Orchids, some of which were of especial service. Mr. R. Parker also sent me an extremely valuable series of forms. Lady Dorothy Nevill most kindly placed her magnificent collection of Orchids at my disposal. Mr. Rucker of West Hill, Wandsworth, sent me repeatedly large spikes of Catasetum, a Mormodes of extreme value to me, and some Dendrobiums. Mr. Rodgers of Sevenoaks has given me interesting information. Mr. Bateman, so well known for his magnificent work on Orchids, sent me a number of interesting forms, including the wonderful Angræcum sesquipedale.
I am greatly indebted to Mr. Turnbull of Down for allowing me the free use of his hot-houses, and for giving me some interesting Orchids; and to his gardener, Mr. Horwood, for his aid in some of my observations.
Professor Oliver has kindly aided me with his large stores of knowledge, and has called my attention to several papers. Lastly, Dr. Lindley has sent me fresh and dried specimens, and has in the kindest manner helped me in various ways.
To all these gentlemen I can only express my cordial thanks for their unwearied and generous kindness.
* I am particularly indebted to Dr. Hooker, who on every occasion has given me his invaluable advice, and has never become weary of sending me specimens from the Royal Gardens at Kew.
EPIDENDREÆ.
In the arrangement given by Lindley in that invaluable work the Vegetable Kingdom, we first meet with the two great tribes of the Malaxeæ and Epidendreæ. They are characterised by the pollen-grains cohering into large waxy masses, not congenitally attached to the rostellum. In the Malaxeæ (of which one British form has been described) the pollinia properly have no caudicle; whereas, in the Epidendreæ (of which tribe no British representative exists), they have an unattached caudicle.
For my purpose these two tribed might have been run together; and as the pollinia of some of the Malaxeæ have a minute but perfectly efficient caudicle, the division between them, in their leading character, blends away. But this is a misfortune which every naturalist encounters in attempting to clarify a largely-developed or so-called natural group, in which, relatively to other groups, there has been little extinction. In order that the naturalist may be enabled to give precise and clear definitions of his
divisions, whole ranks of intermediate or gradational forms must have been utterly swept away: if here and there a member of the intermediate ranks has escaped annihilation, it puts an effectual bar to any absolute distinct definition.
I will begin with the genus Cattleya, of which I have seen several species, and which I fertilised in a very simple manner, different from that in any British Orchid. The rostellum (r, Fig. A, B) is a broad, tongue-shaped projection, which arches slightly over the stigma: the upper surface is formed of smooth membrane; the lower surface and the central portions (originally a mass of cells) consist of a very thick layer of viscid matter. This viscid mass is hardly separated from the viscid matter thickly coating the stigmatic surface which lies close beneath the rostellum. The generally projecting upper lip of the anther rests on, and opens close over, the base of the upper membranous surface of the tongue-shaped rostellum. The anther is kept closed by a sort of spring at the back, at its point of attachment to the top of the column. The pollinia consist of
Fig. XIX.
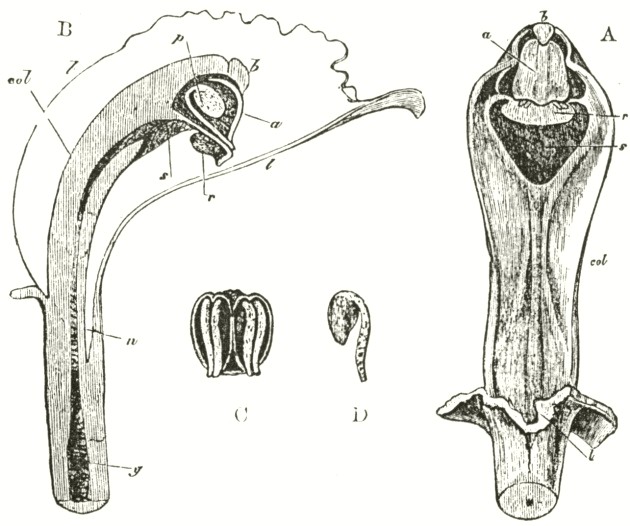
CATTLEYA.
a. anther.
b. spring at the top of the column.
p. pollen-masses.
r. rostellum.
s. stigma.
col. column.
l. labellum.
n. nectary.
g. ovarium, or germen.
A. Front view of column, with all the sepals and petals removed.
B. Section and lateral view of the flower, with all the sepals and petals removed, except the bisected labellum shown only in outline.
C. Anther viewed on the under side, showing the four caudicles with the four pollen-masses beneath.
D. A single pollinium, viewed laterally, showing the pollen-mass and caudicle.
four (or eight in Cattleya crispa) waxy masses, each furnished (see Fig. C and D) with a ribbon-like tail, formed of a bundle of highly elastic threads, with numerous pollen-grains adhering to them. Hence the pollen is of two kinds, waxy masses and separate grains (each, as usual, consisting of four granules) united by elastic threads: this latter kind of pollen is identical with that of Epipactis and other Neotteæ.* These tails, though consisting of good pollen, act also as caudicles, and are thus designated, for they serve as the means for removal of the large waxy masses from the anther-cells. The tips of the caudicles are generally reflexed, and in the mature flower protrude a little way out of the anther-case (see Fig. A), and lie on the base of the upper membranous lip of the rostellum. The labellum enfolds the column, making the flower tubular, and its lower part is produced into a nectary, which penetrates the ovarium.
* The pollen-masses of Bletia are admirably represented on a gigantic scale in Bauer's drawings, published by Lindley in his 'Illustrations.'
Now for the action of these parts. If any body of size proportional to that of the tubular flower be forced into it—a dead humble-bee acted best—the tongue-shaped rostellum is depressed, and the object often gets slightly smeared with viscid matter; but in withdrawing the object, the tongue-formed rostellum is upturned, and a surprising quantity of viscid matter is forced over its edges and sides, and at the same time into the lip of the anther, which is slightly raised by the upturning of the rostellum. Thus the protruding tips of the caudicles are instantly glued to the retreating body, and the pollinia are withdrawn. This hardly ever failed to occur in my repeated trials. A living bee or other large insect alighting on the fringed edge of the labellum, and scrambling into the flower, would depress the labellum and would be less likely to disturb the rostellum, until it had sucked the nectar and began to retreat. When a dead bee, with the four waxy balls of pollen dangling by their caudicles from its back, is forced into another flower, some or all of these balls are surely caught by the broad,
shallow, and very viscid stigmatic surface, which likewise tears off the grains of pollen from the threads of the caudicles.
That living humble-bees can thus remove the pollinia is certain. Sir W. C. Trevelyan sent to Mr. Smith of the British Museum (who forwarded to me) a Bombus hortorum— caught in his hot-house, where a Cattleya was in flower—with its whole back, between the wings, smeared with dried viscid matter, and with the four pollinia attached to it by their caudicles, and ready to be caught by the stigma of another flower if the bee had entered one.
The caudicles of the pollinia being free, and the viscid matter from the rostellum not coming into contact with them without mechanical aid, as well as the general manner of fertilisation, are the same in those species which I have seen in Lælia, Leptotes, Sophronitis, Barkeria, Phaius, Evelyna, Bletia and Coelogyne. In Coelogyne cristata the upper lip of the rostellum is much elongated. In Evelyna caravata eight balls of waxy pollen are all united to one caudicle. In the Barkeria the labellum,
instead of enfolding the column, is pressed against it, and this would even more effectually compel insects to brush against the rostellum. In Epidendrum we have a slight difference; for the upper surface of the rostellum, instead of permanently remaining membranous, as in the above-named genera, is so tender that by a touch it breaks up, together with the whole lower surface, into a mass of viscid matter. In this case the whole of the rostellum, with the adherent pollinia, is removed by inects when they retreat from the flower. I observed in E. glaucum that viscid matter exuded from the upper surface of the rostellum when touched, as I have seen in Epipactis; in fact it is difficult to say, in these cases, whether the upper surface of the rostellum should be called membrane or viscid matter.
In Epidendrum floribundum there is a rather greater difference: the anterior horns of the clinandrum (i.e. the cup at the summit of the column in which the pollinia lie) approach each other so closely as to adhere to the two sides of the rostellum, which consequently lies in a nick, with the pollinia over
it: and as, in this species, the upper surface of the rostellum resolves itself into viscid matter, the pollinia become glued to it without any mechanical aid. The pollinia, though thus attached, cannot, of course, be removed out of their anther-cells without the aid of insects. In this species it seems possible (though, from the position of parts, not probable) than an insect might drag the pollinia on to its own stigma. In all the other species of Epidendrum which I have examined, and in all the above-named genera, as the pollinia lie unattached above the rostellum, it is evident that the viscid matter has to be forced upwards into the lip of the anther by a retreating insect, which would thus necessarily carry the pollinia from one flower to the stigma of another flower.
MALAXEÆ
Turning now to the Malaxeæ: in Pleurothallis prolifera and ligulata (?) the pollinia have a minute caudicle, and mechanical aid is requisite to force the viscid matter from the under side of the rostellum into the anther, thus to catch the caudicles and
remove the pollinia. On the other hand, in our British Malaxis and in the Microstylis Rhedii from India, the upper surface of the minute tongue-shaped rostellum becomes viscid and adheres to the pollinia without mechanical aid. In these two genera we have the curious case of the lower and flat surface of the rostellum being coated with a thin layer of viscid matter, apparently for the purpose of securing the pollinia in a proper position when brought by insects, so as to enter or remain in the slit-like stigma. In Stelis racemiflora the pollinia had also apparently (for the flowers were not in a good condition) become spontaneously attached to the rostellum; and I mention this latter flower chiefly because some insect in the hot-house at Kew had removed most of the pollinia, and had left some of them adhering to the lateral stigmas. These curious little flowers are widely expanded and much exposed; but afer a time the three petals close with perfect exactness and shut up the flower, so that it is scarcely possible to distinguish an old flower from a bud; yet, to my surprice, I found that the closed flowers opened under water.
The allied Masdevallia fenestrata is an extraordinary flower; for the three sepals, instead of closing, as in Stelis after the flower has remained for a time expanded, always cohere together and never open. Two minute, lateral, oval windows (hence the name fenestrata), seated high up the flower and opposite each other, afford the only entrance into the flower; but the presence of these two minute windows (Fig. XX.) shows how necessary it is that insects should have access in this case as with all other Orchids. How insects perform the act of fertilisation I have failed to understand. At the bottom of the roomy and dark chamber formed by the closed sepals, the minute column is placed, in front of which the furrowed labellum stands, with a highly flexible hinge, and on each side the two upper petals; a little tube being thus formed. Hence, when a minute insect enters, or a larger insect inserts its proboscis through either window, it has by
Fig. XX.
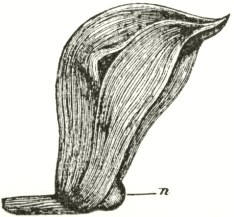
Masdevallia fenestrata.
The window on the near side is shown darkly shaded.
n. nectary.
touch to find the inner tube in order to reach the curious nectary at its base. Within this little tube, formed by the column, labellum and lateral petals, a very broad and hinged rostellum projects at right angles, the under surface of which is viscid; the minute caudicles of the pollinia, projecting out of the anther-case, rest on the base of the upper membranous surface of the rostellum. The stigmatic cavity is deep. After cutting away the sepals I vainly endeavoured, by pushing a bristle into the tubular flower, to remove the pollinia. The whole structure of the flower seemed carefully intended to prevent the withdrawal of the pollinia, as well as their subsequent insertion into the stigmatic chamber! Some new and curious contrivance has here to be made out.
Of Bolbophyllum I examined the curious little flowers of four species, which I will not attempt fully to describe. In B. cupreum and cocoinum, the upper and lower surfaces of the rostellum resolve themselves into viscid matter, which has to be forced upwards by the insects into the anther, so as to secure the pollinia. I effected this easily by passing a needle
down and withdrawing it from the flower, which is rendered tubular by the position of the labellum. In B. rhizophoræ the anther-case moves back, when the flower is mature, and leaves the two pollen-masses fully exposed and adhering spontaneously to the upper surface of the rostellum. The two pollen-masses adhere together by viscid matter, and, judging from the action of the bristle, are always removed together. The stigmatic chamber is very deep, and its orifice is oval, and is exactly fitted by one of the two pollen-masses. After the flower has remained open for some time, the sides of the oval orifice of the stigmatic chamber close in and shut it completely,—a fact which I have observed in no other Orchid, and which, I presume, is here related to the much exposed condition of the whole flower. When the two pollinia were attached to a needle or bristle, or were forced against the stigmatic chamber, one of the two masses glided into the small orifice more readily than could have been anticipated. Nevertheless it is evident that insects must place themselves on successive visits in precisely the same position, so as first to remove the two pollinia,
and then force one of them into the stigmatic orifice. The two upper filiform petals would serve as guides to the insect; but the labellum, instead of making the flower tubular, hangs just like a tongue out of a widely open mouth.
The labellum in all the species which I have seen, more especially in B. rhizophoræ, is remarkable by being joined to the base of the column by a very narrow, thin, white strap, which is highly elastic and flexible; it is even highly elastic when stretched, like an india-rubber band. When the flowers of this latter species were blown by a breath of wind the tongue-like labellums all waggled about in a very odd manner. In some species not seen by me, as in B. barbigerum, the labellum is furnished with a beard of fine hairs, and these cause the labellum to be in almost constant motion from every breath of air. What the use can be of this extreme flexibility and liability to movement in the labellum, I cannot conjecture, unless it be to attract the notice of insects to their dull-coloured, small, and inconspicuous flowers, in the same manner as the bright colours and strong odours of many other Orchids apparently serve to attract insects.
Of the many singular properties of Orchids, the irritability of the labellum in several distantly-allied forms is highly remarkable. When touched, it is described as quickly moving. This is the case with some of the species of Bolbophyllum, but I could not detect any irritability in the species which I examined; nor have I, much to my regret, seen any Orchid with an irritable labellum. The Australian genus Calæna is endowed with this property in a most remarkable degree; for when an insects alights on its labellum it suddenly shuts up against the column, and encloses its prey as it were in a box. Dr. Hooker* believes that this action aids in some manner in the fertilisation of the plant.
The last genus of the Malaxeæ which I will mention is Dendrobium, of which one at least of the species, namely D. chrysanthum, is interesting, from being apparently contrived to affects its own fertilisation, if an insect, when visiting the flower, should accidentally fail to remove the pollen-masses. The rostellum has an upper and a small lower surface composed of membrane; and between these a thick
* 'Flora of Tasmania,'vol. ii. P. 17, under Calæna.
Fig. XXI.
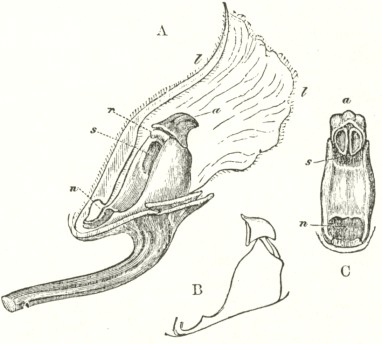
DENDROBIUM CHRYSANTHUM.
a. anther.
r. rostellum.
s. stigma.
l. labellum.
n. nectary.
A. Lateral view of flower, with the anther in its proper position, before the ejection of the pollinia. All the sepals and petals are removed except the labellum, which is longitudinally bisected.
B. Outline of column, viewed laterally, after the anther has ejected the pollinia.
C. Front view of column, showing the empty cells of the anther, after it has ejected its pollinia. The anther is represented hanging too low, and covering more of the stigma than it really does.
mass of milky-white matter is included, which can be very easily forced out. This white matter is less viscid than usual; when exposed to the air a film forms over it in less than half a minute, and it soon sets into a waxy or cheesy substance. Beneath the rostellum the large concave but shallow viscid stigmatic surface is seated. The produced anterior lip of the anther (see A) almost entirely covers the upper surface of the rostellum. The filament of the anther is of considerable length, but is hidden in the sideview, A, behind the middle of the anther; in the section, B, it is seen, after it has sprung forward: it is elastic, and presses the anther firmly down on the inclined surface of the clinandrum (see section B) which lies behind the rostellum. When the flower is expanded the two pollinia, united into a single mass, lie quite loose on the clinandrum and under the anther-case. The labellum embraces the column, leaving a tubular passage in its front; the middle portion (as may be seen in its section, in Fig. A) is thickened; the thickened portion extends up as far as the top of the stigma. The lowest part of the labellum is
developed into a saucer-like nectary, which secretes honey.
If an insect forced its way into one of these flowers, the labellum, which is elastic, would yield, and the projecting lip of the anther would protect the rostellum from being disturbed; but when the insect retreats, the lip of the anther will be lifted up, and the viscid matter from the rostellum will be forced into the anther, gluing the pollen-mass to the insect, which will thus transport it to another flower. I easily imitated this action; but as the pollen-masses have no caudicle, and lie rather far back within the clinandrum under the anther, and as the matter from the rostellum is not highly viscid, they were sometimes left behind and not caught.
Owing to the inclination of the base of the clinandrum, and owing to the length and elasticity of the filament, when the anther was lifted up it was always shot over the rostellum, and remained hanging there, with its lower empty surface (Fig. C) suspended over the summit of the stigma. The filament now stretches across the space (see Fig. B) which was originally covered by the anther. Several
times, having cut off all the petals and labellum, and laid the flower under the microscope, I raised with a needle the lip of the anther, without disturbing the rostellum, and saw the anther assume, with a spring, the position represented sideways in Fig. B, and frontways in Fig. C. By this springing action the anther scoops the pollen-mass out of the concave clinandrum, and pitches it up in the air, with exactly the right force so as to fall down on the middle of the viscid stigma, where it sticks.
Under nature, however, the action cannot be as thus described; for the labellum hangs downwards; and to understand what follows, the drawing should be placed nearly upside down, in an almost reversed position. If an insect failed to remove the pollinium by means of the viscid matter from the rostellum, the pollinium would first be jerked downwards on the protuberant surface of the labellum, placed immediately beneath the stigma. But it must be remembered that the labellum is elastic, and that at the same instant that the insect, in the act of leaving the flower, lifted up the lid of the anther, and so caused the
pollen-mass to be shot out, the labellum would rebound back, and striking the pollen-mass would pitch it upwards, so as to hit the sticky stigma. Twice I succeeded in effecting this, with the flower held in its natural position, by imitating the retreat of an insect; and on opening the flower I found the pollen-mass glued to the stigma.
This view of the use of the elastic filament, seeing how complicated the action must be, may appear fanciful; but we have seen so many and such curious adaptations, that I cannot believe the strong elasticity of the filament and the thickening of the middle of the labellum to be useless points of structure. If the action be as I have described, it might be an advantage to the plant that its single large pollen-mass should not be wasted, if it failed to adhere to an insect by means of the viscid matter from the rostellum. This contrivance is not common to all the species of the genus; for in neither D. bigibbum nor D. formosum was the filament of the anther elastic, nor was the middle line of the labellum thickened. In D. tortile the filament was elastic; but as I saw only one flower,
before I had made out the structure of D. chrysanthum, I cannot say how it acts.
VANDEÆ
We now come to Lindley's immense Tribe of the Vandeæ, which includes many of the most extraordinary productions of our hot-houses, but has no British representative. I have examined twenty-four genera. The pollen consists of waxy masses, as in the two last Tribes, and each ball of pollen is furnished with a caudicle, which becomes, at an early period of growth, united to the rostellum. The caudicle is seldom attached directly to the viscid disc, as in the Ophreæ, but to the upper and posterior surface of the rostellum; and this part, together with the disc, is removed by insects. The imaginary diagram (Fig. XXII), with the parts separated, will best explain the type-structure of the Vandeæ. The middle organ (2) is the dorsal or posterior pistil of the three pistils always present in Orchids; its upper part is modified into and forms the rostellum, and is curved over the stigma. The stigma consists of two stigmas belonging to
Fig. XXII.
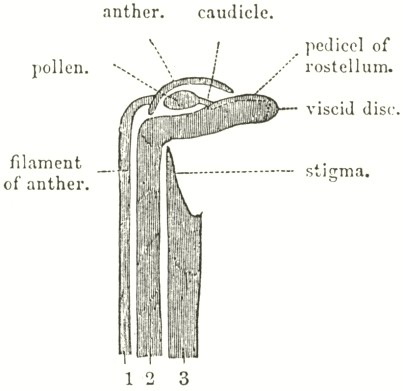
DIAGRAM, Illustrative of the structure of the column in the
VANDEÆ.
(1.) The filament, bearing the anther with its pollen-masses: the anther is represented after it has opened along its whole under surface, which consequently does not appear in the section here given.
(2.) The upper pistil modified into the rostellum.
(3.) The two lower confluent pistils, bearing the two confluent stigmas.
the two other confluent (3) pistils. On the left hand we have the filament (1) bearing the anther. The anther opens at an early period, and the tips of the two caudicles protrude through a small slit (only one caudicle and one pollen-mass is represented in the diagram) in a not fully-hardened condition, and adhere to the back of the
rostellum. The surface of the rostellum is generally hollowed out for the reception of the pollen-masses; it is represented as smooth in the diagram, but is really often furnished with crests or knobs for the attachment of the two caudicles. The anther afterwards opens more widely on its under surface, and leaves the two pollen-masses unattached, excepting by their caudicles to the rostellum.
During this early period of growth, a remarkable change has been going on in the rostellum: either its extremity or its lower surface becomes extremely viscid, and a line of separation, at first appearing as a mere hyaline zone of tissue, gradually is formed, which separates the viscid extremity or disc, as well as the whole upper surface of the rostellum, as far back as the point of attachment of the caudicles. If any object now touches the viscid disc, it, the whole back of the rostellum, the caudicles and pollen-masses, can all be readily removed together. In botanical works the whole structure between the disc (generally called the gland) and the waxy balls of pollen is designated as the caudicle; but as these parts play an
essential part in the fertilisation of the flower, and as they are fundamentally different in their origin and in their minute structure, I shall call the two elastic ropes, which are developed strictly within the anther-cells, the caudicles; and the portion of the rostellum to which the caudicles are attached (see diagram), and which is not viscid, the pedicel. The viscid portion of the rostellum I shall call, as heretofore, the viscid disc. The whole may be conveniently spoken of as the pollinium.
In the Ophreæ we always have (except in O. pyramidalis) two separate viscid discs. In the Vandeæ, with the exception of Angræcum, we have only one disc. The disc is naked, or is not enclosed in a pouch. In Habenaria the discs, as we have seen, are separated from the two caudicles by short drum-like pedicels, answering to the single and generally much more largely developed pedicel in the Vandeæ. In Ophreæ the caudicles of the pollinia, though elastic, are rigid, and serve to place the packets of pollen at the right distance from the insect's head or proboscis, so as to reach the stigma. In the Vandeæ this end is gained by the
pedicel of the rostellum. The two caudicles in the Vandeæ are attached and embedded within a deep cleft in the pollen-masses, and until stretched are rarely visible, for the pollen-masses lie close to the pedicel of the rostellum. These caudicles answer both in position and function to the elastic threads, by which the packets of pollen are tied together in the Ophreæ, at the point where they become confluent and where they form the upper part of the caudicle; for the function of the true caudicle in the Vandeæ is to break when the masses of pollen, transported by insects, adhere to the stigmatic surface.
In many Vandeæ the caudicles are easily ruptured, and the fertilisation of the flower, as far as this point is concerned, is a simple affair; but in other cases the strength of the caudicles and the length to which they can be stretched before they break is surprising. I was at first perplexed to understand what good purpose the great strength of the caudicles and their capacity of extension could serve. It is obvious that, when projecting far out from an insect' head, whilst flying about (and the insect, in the
case of the larger Orchids, must be of considerable size), the strength of the caudicles would protect the pollen-masses from being brushed off and lost. So again, when an insect transporting a pollinium visits a flower either too young, with its stigma not yet sufficiently viscid, or one already impregnated, with its stigma beginning to dry, the strength of the caudicle would prevent the pollen-masses from being uselessly removed. It should be remembered that the pollen-masses are precious objects, for each flower, in most of the genera, produces only two; and in many cases, judging from the size of stigma, both pollen-masses would be left on one stigma, though in other cases the size of the orifice of the stigma allows the introduction of one pollen-mass alone; so that, in the latter case, the pollen from one flower probably suffices to fertilise two flowers.
Although at the proper period the stigmatic surface is astonishingly viscid in many cases, as in Phalænopsis and Saccolabium, yet when, having removed the pollinia adhering to a rough scalpel by their viscid discs, I inserted the balls of pollen into
the stigmatic chamber, they did not adhere to the surface with sufficient force to prevent their withdrawal. I even left them for some little time in contact with the viscid surface, as an insect would do whilst feeding; but when I pulled the pollinia straight out of the stigmatic chamber, the caudicles, though they were stretched to a great length, did not rupture, nor did the viscid disc separate from the scalpel; consequently the balls of pollen were not left on the stigma. It then occurred to me that an insect in flying away would not pull the pollinia straight out of the chamber, but would pull at nearly right angles to its orifice. When I thus acted, the stretched caudicles were necessarily dragged over the margin of the chamber, and the friction thus caused, together with the vicidity of the stigmatic surface, generally ruptured them and left the pollen-masses on the stigma. Thus, it seemed that the great strength and extensibility of the caudicles, which, until stretched, lie embedded within the pollen-masses, serve to protect the pollen-masses from being wasted, and yet, by friction being brought into play, allow
them, at the proper time and by means of insects, to be left adhering to the stigmatic surface, and the fertilisation of the flower to be safely effected.
The discs and the pedicel of the rostellum in the Vandeæ present great diversities in shape, and an apparently exhaustless number of adaptations. Even in species of the same genus, as in Oncidium, these parts differ greatly. I have here given a few figures (Fig. XXIII.), taken almost at hazard. The pedicel generally consists (as far as I have seen) of a piece of thin ribbon-
Fig. XXIII.
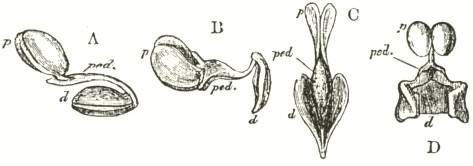
POLLINIA OF THE VANDEÆ.
d. viscid disc.
ped. pedicel.
p. pollen-masses.
The caudicles, being embedded within the pollen-masses, are not shown.
A. Pollinium of Oncidium grande after partial depression.
B. Pollinium of Brassia maculata (copied from Bauer).
C. Pollinium of Stanhopea saccata after depression.
D. Pollinium of Sarcanthus teretifolius after depression.
shaped membrane (Fig. A), long or short; but sometimes it is almost (Fig. C) cylindrical and often of all sorts of shapes. The pedicel is generally nearly straight, but in Miltonia Clowesii it is naturally curved; and in some other cases, as we shall immediately see, it assumes, after removal, various shapes. The extensible and elastic caudicles, by which the pollen-masses are attached to the pedicel, are here not visible, being embedded in a cleft or hollow within each pollen-mass. The disc, which is viscid on the under side, consists of a piece of thin or thick membrane of the most diversified shapes. In Acropera it is like a pointed cap; in some cases it is tongue-shaped, or heart-shaped (Fig. C), or saddle-shaped, as in some Maxillarias, or like a thick cushion (Fig. A), as in many species of Oncidium, with the pedicel attached at one end, instead of, as is more usual, nearly to its centre. In Angræcum distichum and sesquipedale the rostellum is notched, and two separate, thin, membranous discs can be removed, each carrying by a short pedicel its pollen-mass. In Sarcanthus teretifolius the disc (Fig. D) is very oddly shaped; and as
the stigmatic chamber is very deep, and likewise curiously shaped, one is tempted to believe that the disc has to be fastened with great precision on to the square projecting head of some insect.
In most cases there is a plain relation between the length of the pedicel and the depth of the stigmatic chamber, into which the pollen-masses have to be inserted; in some few cases, however, in which there is a long pedicel and a shallow stigma, we shall meet with curious compensating actions. After the disc and pedicel have been removed, the shape of the rostellum is altered, being generally only slightly shortened and made thinner: sometimes it becomes notched: in Stanhopea, the entire circumference of the extremity of the rostellum is removed, and a thin, pointed, needle-like process alone is left, which originally ran up its centre.
If we turn to the former imaginary diagram (Fig. XXII., ), and suppose the rectangularly bent rostellum to be thinner and the stigma to lie closer under it, we shall see that, if an insect with the pollinium attached to its head were to fly to another flower and
occupy almost exactly the same position which it held when the attachment of the disc was effected, the pollen-masses would strike the stigma, especially if, from their weight, they had become in the least degree depressed. This is all that takes place in Lycaste Skinnerii, Cymbidium giganteum, Zygopetalum Mackai, Angræcum eburneum, Miltonia Clowesii, in a Warrea, and, I believe in Galeandra Funkii. But if in our diagram we suppose the stigma to be seated lower down at the bottom of a deep cavity, or suppose the anther to be seated higher up so that the pedicel of the rostellum sloped upwards, and under other contingencies not worth detailing, all of which occur,—in such cases, an insect with the pollinium attached to its head, on flying to another flower would not strike the stigma with the pollen-masses unless some great change had intervened in their position after attachment.
This change is effected in many Vandeæ in the same manner as is so general with the Ophreæ, namely, by a movement of depression in the pollinium in the course of about half a minute after its removal from
the rostellum. I have seen this movement conspicuously displayed, generally causing the pollinium after detachment to rotate through about a quarter of a circle, or 90°, in several species of Oncidium, Odontoglossum, Vanda, Aerides, Sarcanthus, Saccolabium, Acropera, and Maxillaria. In Rodriguezia suaveolens the movement of depression is remarkable from its extreme slowness; in Eulophia viridis from its small extent. In some, indeed, of the cases specified of the pollinia undergoing no movement, I am not sure that there is not a very slight depression. In the Ophreæ the anther-cells are sometimes seated exteriorly and sometimes interiorly with respect to the stigma; and we accordingly have outward and inward movements in the pollinia; but in the Vandeæ the anther-cells lie directly over the stigma, and the movement of the pollinium is always directly downwards. In Calanthe, however, the two stigmas are placed exteriorly to the anther-cells, but the pollinia, as we shall see, are made to strike them by a mechanical arrangement of the parts.
In the Ophreæ the seat of contraction,
which causes the act of depression, is in the upper surface of the viscid disc, near the point of attachment of the caudicle: in the Vandeæ the seat is likewise in the upper surface of the viscid disc, but at the point where the pedicel is united to it, and therefore at a considerable distance from the point of attachment of the true caudicles. The contraction and consequent movement is hygrometric in its nature (but the subject, as we shall see in the seventh chapter, is rather obscure), and consequently does not take place until the pollinium is removed from the rostellum, and the point of union of the disc and pedicel has been exposed for a few seconds to the air. If, after the contraction and consequent movement of the pedicel, the whole body be placed into water, the pedicel slowly moves back and resumes the same position with respect to the viscid disc which is held when forming part of the rostellum. When taken out of the water, it again undegoes the movement of depression. It is of importance to notice these facts, as we thus get a test by which this movement can be distinguished from certain other movements.
In one species of Maxillaria, viz. in the M. ornithorhyncha, we have a unique case. The pedicel of the rostellum is much elongated, and is entirely covered by the produced front lip of the anther, and is thus kept damp. When removed there is no movement at the junction of the disc and the pedicel; but the pedicel itself, at a point rather more than half-way up, quickly bends backwards on itself, in a reversed direction compared with all other cases. When the pedicel is placed in water it resumes its original straight form. If we suppose the long upright neck of a bird to represent the pedicel, the head representing the balls of pollen, then in all ordinary cases the movement is that of a bird picking up food from the ground, but bending only the lower vertebræ close to its body; whereas, in this Maxillaria, the movement is as if the bird threw its head backwards so as nearly to touch its own back, the middle vertebræ of the neck being alone bent. I stated above that, when the pedicel is long and the stigmatic cavity shallow, as in this Maxillaria, we have a compensating action; and here we have one instance. The labellum
has a square projection in front of the stigma, the passage into the flower being thus contracted; and if the pedicel of the rostellum did not somehow become shortened, the flower could hardly be fertilised. After the reversed movement just described, and the consequent shortening of the pedicel, the pollinium, when attached to any small object, can be inserted into the flower, and the balls of pollen are so placed as readily to adhere to the stigmatic surface.
In some cases, besides these hygrometric movements, elasticity comes into play. In Aerides odorata and virens, and in Oncidium (roseum?), the pedicel of the rostellum is fastened down in a straight line, by the disc at one extremity and by the anther at the other; it has, however, a strong natural elastic tendency to spring up at right angles to the disc. Consequently, when the pollinium is removed by its viscid disc sticking to any object, the pedicel instantly springs up and stands at nearly right angles to its former position, with the pollen-masses carried aloft. This has been noticed by other observers; and I agree with then that the object gained
is to free the pollen-masses from their anther-cells. After this upward elastic spring, the downward hygrometric movement immediately commences, which, oddly enough, carries the pedicel back again into almost the same position, relative to the disc, which it held whilst adhering to the rostellum. The end of the pedicel, however, which in Aerides carries the pollen-mass by short dangling caudicles, remains curved a little upwards; and this seems well adapted to drop the pollen-masses into the deep stigmatic cavity over a ledge in front. The difference between the first elastic and the second or reversed hygrometric movement, was well seen (I tried it only in the Oncidium) by placing the pollinium, after both movements had taken place, into water, when the pedicel moved into the position which it has at first acquired by elasticity, which latter position was not in any way affected by the water. When taken out of the water the hygrometric movement of depression soon recommenced.
In Rodriguezia secunda there was no slow movement of depression in the pedicel as in
the before-mentioned R. suaveolens, but there was a rapid downward movement, due, apparently, in this one case, to elasticity; for when the pedicel was put into water it showed no tendency to recover its original position, as occurs with all the many other Orchids which I have examined.
In Phalænopsis grandiflora and amabilis the stigma is shallow and the pedicel of the rostellum long. Consequently a compensating action is requisite, which, differently from the case of the Maxillaria, is effected by elasticity. There is no movement of depression; but, when the pollinium is removed, the straight pedicel suddenly curls up, thus (.—^—). The full-stop on the left hand may represent the balls of pollen, but the disc on the right hand must be imagined to be a triangular piece of membrane. The pedicel does not straighten itself in water. The end carrying the balls of pollen after the contraction is a little raised up, and the pedicel, with one end raised, and with the middle part upwardly bowed, seems well adapted to drop the pollen-masses, over a ledge in front, into the deep stigmatic cavity.
In Calanthe Masuca and the hybrid C. Dominii the structure is very different. We here have two oval, pit-like stigmas standing quite laterally on each side of the rostellum (Fig. XXIV.). The viscid disc is oval (Fig. B) and has no pedicel, but eight masses
Fig. XXIV.
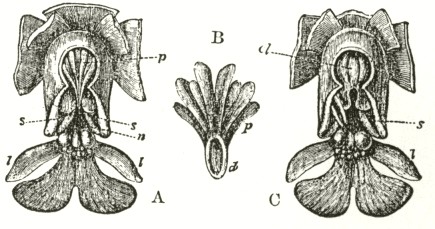
CALANTHE MASUCA.
p. pollen-masses.
s s. the two stigmas.
n. mouth of nectary.
l. labellum.
d. viscid disc.
cl. clinandrum with pollen-masses removed.
A. Flower viewed from above, with the anther-case removed, showing the eigth pollen-masses in their proper position within the clinandrum. All the sepals and petals have been cut away except the labellum.
B. Pollen masses attached to the viscid disc, seen from the under side.
C. Flower in the same position as in A, but with the disc and pollen-masses removed, showing the now divided rostellum and the empty clinandrum in which the pollen-masses lay. Within the left-hand stigma two pollen-masses may be seen adhering to its viscid surface.
of pollen are attached to it by very short and easily ruptured caudicles. These pollen-masses radiate from the disc like the spokes of a fan. The rostellum is broad and its sides slope on each side towards the lateral pit-like stigmas. When the disc is removed the rostellum is seen (Fig. C) to be deeply divided in the middle. The labellum is united to the column almost up to its summit, leaving a passage (n, A) to the long nectary close beneath the rostellum. The labellum is studded with singular, wart-like, globular excrescences.
If a thick needle be inserted into the mouth of the nectary (Fig. A), and then withdrawn, the viscid disc will be withdrawn, bearing with it the elegant fan of radiating pollen-masses. These undergo no change in position. But if the needle be now inserted into the nectary of another flower the ends of the pollen-masses necessarily hit the upper and laterally sloping sides of the rostellum, and, glancing off both ways, they strike down into the two lateral pit-like stigmas. The thin caudicles being easily ruptured, the pollen-masses are left adhering like little darts (see
left-hand stigma in Fig. C) to the viscid surface of both stigmas, and the fertilisation of the flower is completed in a simple manner pleasing to behold.
I should have stated that a narrow transverse rim of stigmatic tissue, running beneath the rostellum, connects the two lateral stigmas; and it is probable that some of the middle pollen-masses may be inserted through the notch in the rostellum or beneath its surface. I am more inclined to this opinion from having found in the elegant Calanthe vestita that the rostellum extends so widely over the two lateral stigmas, that apparently all the pollen-masses would have been inserted beneath its surface.
I fear that the reader will be wearied, but I must say a few words on the Angræcum sesquipedale, of which the large six-rayed flowers, like stars formed of snow-white wax, have excited the admiration of travellers in Madagascar. A whip-like green nectary of astonishing length hangs down beneath the labellum. In several flowers sent me by Mr. Bateman I found the nectaries eleven and a half inches long, with only the lower
inch and a half filled with very sweet nectar. What can be the use, it may be asked, of a nectary of such disproportional length? We shall, I think, see that the fertilisation of the plant depends on this length and on nectar being contained only within the lower and attenuated extremity. It is, however, surprising that any insect should be able to reach the nectar: our English sphinxes have probosces as long as their bodies: but in Madagascar there must be moths with probosces capable of extension to a length of between ten and eleven inches!
The rostellum is broad and foliaceous, and arches rectangularly over the stigma and over the orifice of the nectary: it is deeply cleft, with the cleft enlarged or widened at the end. Hence the rostellum pretty closely resembles (see Fig. XXIV., C) that of Calanthe after the disc has been removed. The under surfaces of both margins of the cleft near its end are bordered by narrow strips of viscid membrane, easily removed; so that there are two distinct viscid discs. To the middle of each disc a short membranous pedicel is attached; and each pedicel carries
at its other end a pollen-mass. Beneath the rostellum a narrow, ledge-like, viscid stigma is seated.
I could not for some time understand how the pollinia of this Orchid were removed, or how it could be fertilised. I passed bristles and needles down the open entrance into the nectary and through the cleft in the rostellum with no result. It then occurred to me that, from the length of the nectary, the flower must be visited by large moths, with a proboscis thick at the base; and that to drain the last drop of nectar even the largest moth would have to force its proboscis as far down as possible. To effect this, whether or not the moth first inserted its proboscis by the open entrance into the nectary (as is most probable, from the shape of the flower, &c.) or through the cleft in the rostellum, it would ultimately force its proboscis into this cleft, for this is the straightest course, and by slight pressure the whole foliaceous rostellum can be depressed: the distance from the outside of the flower to the extremity of the nectary can be thus shortened by about a quarter of an inch. Hence I took a cylinder, one-tenth of an inch
in diameter, and pushed it down through the cleft in the rostellum: the margins readily separated, and were pushed downwards together with the whole rostellum. When I slowly withdrew the cylinder the rostellum rose from its elasticity, and the margins of the cleft upturned and clasped the cylinder. Thus the viscid strips of membrane on the under sides of the cleft rostellum came into contact with the cylinder, and firmly adhered to it; and the pollen-masses were withdrawn. By this means alone I succeeded in each case in withdrawing the pollinia; and it cannot, I think, be doubted that a large moth must thus act; namely, by driving its proboscis up to the very base, through the cleft of the rostellum, so as to reach the extremity of the nectary; and then withdrawing its proboscis with the pollinia attached to it.
I did not succeed in imitating the fertilisation of the flower so well as I did in withdrawing the pollinia, but I effected it twice. As the margins of the cleft rostellum must be upturned before the discs adhere to the cylinder, they become, during its with-
drawal, affixed some little way from its actual base. The two discs did not always adhere at exactly corresponding points. Now, when a moth inserts its proboscis, with the pollinia affixed to it near the base, into the mouth of the nectary, the pollen-masses will probably be first inserted beneath the rostellum; and during the final exertion, when the moth pushes its proboscis through the cleft of the rostellum, the pollen-masses will almost necessarily be placed on the narrow, ledge-like stigma projecting beneath the rostellum. By acting thus with the pollinia attached to the cylinder the pollen-masses were twice torn off and left glued to the stigmatic surface.
If the Angræcum in its native forests secretes more nectar than did the vigorous plants sent me by Mr. Bateman, so that the nectary becomes filled, small moths might obtain their share, but they would not benefit the plant. The pollinia would not be withdrawn until some huge moth, with a wonderfully long proboscis, tried to drain the last drop. If such great moths were to become extinct in Madagascar, assuredly the Angræcum would become extinct. On the other
hand, as the nectar, at least in the lower part of the nectary, is stored safe from depredation by other insects, the extinction of the Angræcum would probably be a serious loss to these moths. We can thus partially understand how the astonishing length of the nectary may have been acquired by successive modifications. As certain moths of Madagascar became larger through natural selection in relation to their general conditions of life, either in the larval or mature state, or as the proboscis alone was lengthened to obtain honey from the Angræcum and other deep tubular flowers, those individual plants of the Angræcum which had the longest nectaries (and the nectary varies much in length in some Orchids), and which, consequently, compelled the moths to insert their probosces up to the very base, would be fertilised. These plants would yield most seed, and the seedlings would generally inherit longer nectaries; and so it would be in successive generations of the plant and moth. Thus it would appear that there has been a race in gaining length between the nectary of the Angræcum and the proboscis
of certain moths; but the Angræcum has triumphed, for it flourishes and abounds in the forests of Madagascar, and still troubles each moth to insert its proboscis as far as possible in order to drain the last drop of nectar.
Finally, another genus, Acropera, must be noticed from an independent reason. Although Dr. Hooker sent me, over and over again, fresh flowers of two species (A. luteola* and Loddigesii), this genus for a long time remained the opprobrium of my work. All the parts seemed determinately contrived that the plant should never be fertilised. I believe I have at last partly solved the mystery; yet the use of some important parts remains quite unintelligible. But I do not suppose that I completely understand the contrivances in any one Orchid; for I find that the more I study our most commonest British species, continually new and admirable adaptations become apparent.
* Dr. Lindley informs me that he knows of no species so named; nor is the origin of the name known at Kew. This species or variety differs in little or nothing from A. Loddigesii, excepting in its uniform yellow colour.
The rostellum of Acropera is thin and elongated, projecting at right angles to the column (see diagram, Fig. XXII.); the pedicel of the pollinium is of course equally long and very thin; the disc is extraordinarily small, and forms a little cap, viscid within, fitting the extremity of the rostellum. After repeated trials I find that the disc does not adhere to any object until it is drawn quite off the tip of the rostellum; and this can only be well effected by the whole rostellum being pushed upwards so as to slide over and against the touching object: when the small disc is thus removed it adheres well to the object. The upper sepal forms a hood enclosing and protecting the column. The labellum is an extraordinary organ, baffling description: it is articulated to the base of the column by a thin strap, so elastic and flexible that a breath of wind sets it vibrating. It hangs downwards; and this seems to be of importance, for the plant is pendulous, and to place the labellum in this position the footstalk (ovarium) of each flower is curved into a semicircle. The two upper petals serve as lateral guides leading
into the hood-like upper sepal, But how all these parts concur in leading an insect to push some part of its body into the hood-like upper sepal and then to raise the rostellum, thus brushing off the little sticky disc, I do not in the least understand.
The pollinium, when adhering by its disc to any object, undergoes the common movement of depression; and this seems superfluous, for the stigmatic cavity lies (see diagram, Fig. XXII.) high up at the base of the rectangularly projecting rostellum. But this is a comparatively trifling difficulty; the real difficulty lies in the orifice of the stigmatic chamber being so narrow that the pollen-masses can hardly be forced in. I repeatedly tried, and succeeded only three or four times. Even after allowing the pollen-masses to dry for an hour, and thus to shrink a little, I rarely succeeded in forcing them in. I examined young flowers and almost withered flowers, for I imagined that the mouth of the chamber at different periods of growth might become larger or smaller, for we have seen that the mouth actually closes up in one species of Bolbophyllum; but the difficulty of insertion
always remained the same. Now when we observe, that the viscid disc is extraordinarily small, and consequently its power of attachment not so firm as with Orchids having a large disc, and that the pedicel is long and thin, it would seem almost indispensable that the stigmatic chamber should be unusually large for the easy insertion of the pollinium. Far from this being the case, it is, as just stated, so much contracted that rarely by any force could even one pollen-mass be forced in. Moreover the stigmatic surface, as Dr. Hooker also observed, is singularly little viscid!
I had given up the whole case as inexplicable, when it occurred to me that, although no instance of the separation of the two sexes was known in Orchids, yet that Acropera might be a male plant. I first examined the utriculi of the stigmatic surface from specimens which had been kept in spirits of wine, and I found them empty like little glass cases, but generally with a faint areola or nucleus visible.* Now I have looked at the
* See R. Brown in 'Linn. Transact.,' vol. xvi. p. 710, on the nucleus of the stigmatic utriculi; and Bauer's beautiful drawing in Lindley's great work.
utriculi of very many Orchids, and have hitherto found no exception to the rule that spirits of wine causes a considerable quantity of yellowish-brown matter to coagulate within them. I have taken fresh Orchids and placed them in spirits, and in twenty-four hours have found the contents coagulated and the nuclei much darkened. More numerous observations would be requisite for much stress to be laid on this fact, but I must at present infer that the utriculi of Acropera are in a different condition from those of all other Orchids. The state of the ovarium offers better evidence.
When a thin transverse slice of the ovarium is taken and examined under a quite weak power, small projections are seen on the three proper ovule-bearing cords or segments, which at first seem like true ovules. But when these are more closely examined, they are seen to consist of sub-branched, quite thin and transparent fringes of membrane, which in some specimens exhibited cellular structure far more plainly than in others. If these fringes are placentæ, they are more largely developed than in other Orchids; if they are ovules (or rather the testæ of ovules) in an
atrophied condition, as I believe to be the case, they are more firmly fixed to the placentæ than in other cases; they do not exhibit the proper opening at their free ends, and no nucleus is visible; nor were any of them inverted. I examined six ovaria of young and old flowers of the Acropera, some fresh and some which had been kept in spirits of wine, and all the ovule-bearing cords were nearly in the same condition. I examined, for comparison, the ovaria of Orchids belonging to nearly all the main Tribes, of young and old (but not fertilised) flowers, some of which were fresh, and some kept in spirits, and invariably the ovules presented a widely different appearance.
From these several facts—namely, the narrowness of the mouth of the stigmatic chamber, into which the pollen-masses can hardly be forced, whereas the length and thinness of the pedicel of the rostellum, the smallness of the viscid disc, and the movement of depression, all indicate the necessity of a large stigmatic cavity seated low down—the slight viscidity of the stigmatic surface—the empty condition of the stigmatic utriculi—and espe-
cially the condition of the ovule-bearing cords—lead me to infer that the plant at Kew, from which the many flowers of the Acropera luteola were at different times gathered, is a male plant. From having examined many Orchids grown in hot-houses,* I have no reason to believe that cultivation affects the female organs in the manner described. It is scarcely possible to believe that cultivation could contract the solid edges of the stigmatic chamber. Therefore I see no reason to doubt my conclusion on the male sex of this plant.
What the female of hermaphrodite form of the Acropera luteola may prove to be—whether resembling in most respects the male, or whether it be at present named and masked as some distinct genus—it is impossible to say. In Acropera Loddigesii, which closely resembles, in all respects except in colour, A. luteola, I found the same almost insuperable difficulty in inserting the pollen-
* In a spike of the Brazilian Goodyera discolor, sent me by Mr. Bateman, which had all its flowers monstrous and distorted, with the stigmas imperfect, I found the ovules with their nuclei projecting far out of the testæ (exactly as figured by Brongniart in Epipactis in 'Annales des Sciences,' tom. 24, 1831, pl. 9), and apparently well developed.
masses into the stigmatic cavity, but I did not at that time suspect the masculine nature of the genus, and did not examine the ovarium.
I have now described, perhaps in too much detail, a few of the many contrivances by which the Vandeæ are fertilised. The relative position of the parts—friction, viscidity, elastic and hygrometric movements, all nicely related to each other—come into play. But all these appliances are subordinate to the action of insects. Without their aid, not a plant in this tribe, in the twenty-four genera examined, would set a seed. It is also evident that, in a vast majority of cases, insects would withdraw the pollinia only when retreating from a flower, and, carrying them away, would thus effect a union between two distinct flowers. This fact is conclusively shown in all those many cases in which the pollinia undergo a change of position, after removal from the rostellum, in order to stand in a proper direction to strike the stigma; for this could only be effected after the insect had left one flower, which would serve as the male, and before it visited a second flower, which would serve as the female.
CHAPTER VI.
Catasetidæ, the most remarkable of all Orchids—The mechanism by which the pollinia of Catasetum are ejected to a distance, and are transported by insects—Sensitiveness of the horns of the rostellum—Extraordinary difference in the male, female, and hermaphrodite forms of Catasetum tridentatum—Mormodes ignea, curious structure of flower; ejection of its pollinia—Cypripedium, importance of the slipper-like form of the labellum—Secretion of nectar—Advantage derived from insects being delayed in sucking the nectar—Singular excrescences on the labellum, apparently attractive to insects.
I have reserved for separate description one sub-family of the Vandeæ, namely the Catasetidæ, which may, I think, be considered as the most remarkable of all Orchids.
I will begin with the most complex genus, Catasetum. A brief inspection of the flower shows that here, as with other Orchids, some mechanical aid is requisite to remove the pollen-masses from their receptacles, and to carry them to the stigmatic surface. We shall, moreover, presently see that the three following species of Catasetum are male plants; hence it is certain that their pollen-masses must be transported to female plants,
in order that seed may be produced. The pollinium is furnished with a viscid disc, in this genus of huge size; but the disc, instead of being placed, as in other Orchids, in a position likely to touch and adhere to an insect visiting the flower, is turned inwards and lies close to the upper and back surface of a chamber, which must be called the stigmatic chamber, though functionless as a stigma. There is nothing in this chamber to attract insects; and even if they did enter it, it is hardly possible that the disc should adhere to them, for its viscid surface lies in contact with the roof of the chamber.
How then does nature act? She has endowed these plants with, what must be called for want of a better term, sensitiveness, and with the remarkable power of forcibly ejecting their pollinia to a distance. Hence, when certain definite points of the flower are touched by an insect, the pollinia are shot out like an arrow which is not barbed, but has a blunt and excessively adhesive point. The insect, disturbed by so sharp a blow, or after having eaten its fill, flies sooner or later to a female plant, and, whilst standing in the
same position as it did when struck, the pollen-bearing end of the arrow is inserted into the stigmatic cavity, and a mass of pollen is left on its viscid surface. Thus, and thus alone, at least three species of the genus Catasetum are fertilised.
In many Orchids, as in Listera, Spiranthes, Orchis, we have seen that the surface of the rostellum is so far sensitive, that, when touched or when exposed to the vapour of chloroform, it ruptures in certain defined lines. So it is in the tribe of the Catasetidæ, but with this remarkable difference, that in Catasetum the rostellum is prolonged into two curved tapering horns, or, as I shall call them, antennæ, which stand over the labellum where insects alight, and the excitement of a touch is conveyed along these antennæ to the membrane which has to be ruptured; and when this is effected, the disc of the pollinium is suddenly set free. We have also seen that in several Vandeæ the pedicels of the pollinia are fastened down flat, but are elastic and tend to spring up, so that, as soon as they are freed, they suddenly curl upwards, apparently for the purpose of detaching the pollen-masses
from their anther-cells. In the genus Catasetum, on the other hand, the pedicels are fastened down in a curved position; and when freed by the rupture of the attached edges of the disc, they straighten themselves with such force, that not only do they drag the balls of pollen and anther-cells from their places of attachment, but the whole pollinium is jerked forward, over and beyond the tips of the so-called antennæ, to the distance of two or three feet. Thus, as throughout nature, pre-existing structures and capacities are utilised for new purposes.
Catasetum saccatum.*—I will now enter on details. The general appearance of this species is represented in the following woodcut, Fig. XXV.; B, being a side view of the whole flower, but with all the petals and sepals excepting the labellum cut off, and A, being a front view of the column. The upper sepal and two upper petals surround and protect the column; the two lower sepals project out
* I am much indebted to Mr. James Veitch of Chelsea for the first specimen which I saw of this Orchid; subsequently Mr. S. Rucker, so well known for his magnificent collection of Orchids, generously sent me two fine spikes, and has aided me in the kindest manner with other specimens.
at right angles. The flower stands more or less inclined to either side, but with the labellum downwards. The dull coppery and orange-spotted tints,—the yawning chasm in the great fringed labellum,—the one antenna stuck out with the other hanging down—give to these flowers a strange, lurid, and reptilian appearance.
In front, in the middle of the column (Fig. A), the deep stigmatic chamber (s) may be seen; this is shown in the section (Fig. XXVI. C), in which all the parts are a little separated from each other, in order that the mechanism may be made intelligible. In the middle of the roof of the stigmatic chamber, far back (d, in A), the upturned anterior end of the viscid disc of the pollinium may be discerned. The disc is continuous on each side with a little fringe of membrane, which joins the bases of the two antennæ. Over the disc the protuberant heart-shaped rostellum projects, and this is closely covered by a thin membrane. This membrane is the pedicel of the pollinium, with its lower end attached (ped in sect. C, and in A) to the superior surface of the viscid disc, and with
Fig. XXV.
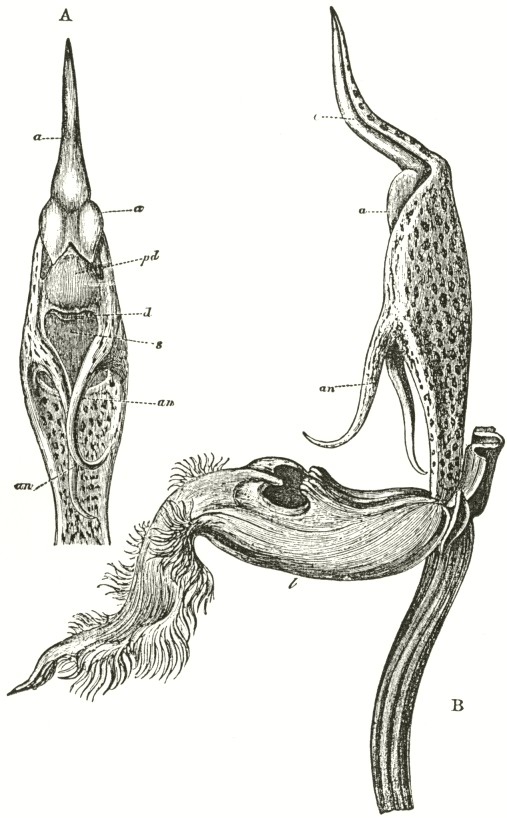
CATASETUM SACCATUM.
Fig. XXVI.
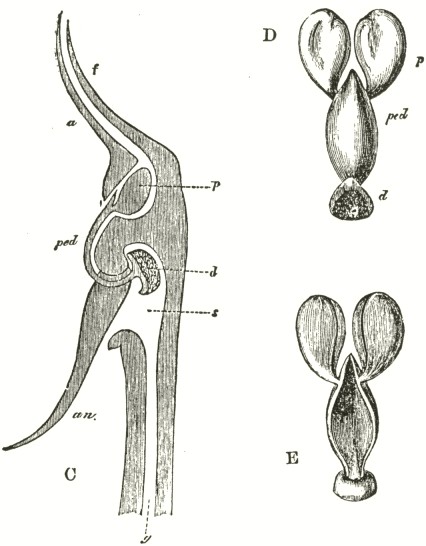
CATASETUM SACCATUM.
a. anther.
an. antennæ of the rostellum.
d . disc of pollinium.
f. filament of anther.
g. germen or ovarium.
l. labellum.
p. pollen-masses.
pd or ped. pedicel of pollinium.
s. stigmatic chamber.
A. Front view of column.
B. Side view of flower, with all the sepals and petals removed except the labellum.
C. Section through the column, with all the parts a little separated.
D. Pollinium, upper surface.
E. Pollinium, lower surface, which lies in contact with the rostellum.
its upper end running under the anther-cells (a), and there united to the two pollen-masses. In its natural position the pedicel lies much bowed round the protuberant rostellum; when freed it forcibly straightens itself, and at the same time its lateral edges curl inwards. In the bud, at an early period of growth, the membranous pedicel forms part of the rostellum, but subsequently becomes separated from it by the solution of a layer of cells.
The pollinium when let free, and after it has straightened itself, is represented at D, Fig. XXVI., and its under surface, which lies in contact with the rostellum, is shown at E, with the lateral edges of the pedicel now much curled inwards. In this latter view, the clefts in the under sides of the two pollen-masses are shown. Within the cleft, at its base, a layer of strong extensible tissue is attached, forming the caudicle, by which the pollen-masses are united to the pedicel. The lower end of the pedicel is joined to the disc by a flexible hinge, which occurs in no other genus, so that the pedicel can play backwards and forwards as far as the upturned end
(Fig. D) of the disc permits. The disc is large and thick; it consists of a strong upper membrane, to which the pedicel is united, with an inferior cushion of great thickness, of pulpy, flocculent, and viscid matter. The posterior margin (or lower margin in Fig. D) is much the most viscid portion, and this necessarily first strikes any object when the pollinium is ejected. The viscid matter soon sets hard. The whole surface of the disc is kept damp before ejection, by resting against the roof of the stigmatic chamber; in the section (Fig. C) it is represented, like the other parts, a little separated from the roof.
The connective membrane of the anther (a in all the figures) is produced into a spike, which adheres loosely to the pointed end of the column, this pointed end (f, Fig. C) homologically is the filament of the anther. The anther is thus shaped apparently to give leverage, so that it may be easily torn off by a pull at its lower end, when the pollinium is let free and is jerked out by the elasticity of the pedicel.
The labellum stands at right angles to the
column, or hangs a little downwards; its lateral and basal lobes are turned under the middle portion, so that an insect can stand only in front of the column. In the middle of the labellum there is a deep cavity, bordered by crests: this cavity does not secrete nectar, but its walls are thick and fleshy, and have a slightly sweet nutritious taste. I believe, as we shall hereafter see, that insects visit the flowers to gnaw these fleshy walls and crests. The extremity of the left-hand antenna stands immediately over the cavity, and would almost certainly be touched by an insect visiting this part of the labellum for any purpose.
The antennæ are the most singular organs of the flower, and occur in no other genus. They form rigid, curved horns, tapering to a point. They consist of a narrow ribbon of membrane, with the edges curled inwards so as to touch, but the edges are not united; hence each horn is tubular, like an adder's fang, with a slit down one side. They are composed of numerous, much elongated, generally hexagonal cells, pointed at both ends; and these cells (like those in most of the other tissues of the flower) have nuclei
with nucleoli. The antennæ are prolongations of the sides of the anterior face of the rostellum. As the viscid disc is continuous with the little fringe of membrane on each side, and as this fringe is continuous with the bases of the antennæ, these latter organs are put in direct connection with the disc. The pedicel of the pollinium passes between the bases of the two antennæ. The antennæ are not free for their whole length; but their exterior edges, for a considerable space, are firmly united to and blended with the margins of the stigmatic chamber.
In all the flowers which I have examined, gathered from three plants, both antennæ occupied the same position; but though otherwise alike, they do not stand symmetrically. The extreme part of the left-hand antenna bends upwards (see Fig. B, in which the position is shown plainer than in A), and at the same time a little inwards, so that its tip is medial and guards the entrance into the pit of the labellum. The right-hand antenna hangs downwards, with its tip turned a little outwards; owing to this position, the crease or furrow formed by its inflexed edges is
externally visible in this antenna; whilst in the other it is hidden along the under side. As we shall immediately see, the depending right-hand antenna is almost paralysed, and is apparently functionless.
Now for the action of the parts. When the left-hand antenna of this species (or either antenna of the two following species) is touched, the edges of the upper membrane of the disc, which are continuously united to the surrounding surface, instantaneously rupture, and the disc is set free. The highly elastic pedicel then instantly flirts the heavy disc out of the stigmatic chamber with such force, that the whole pollinium is ejected, bringing away with it the two balls of pollen, and tearing the loosely attached spike-like anther from the top of the column. The pollinium is always ejected with its viscid disc foremost. I imitated this action with a minute strip of whalebone, slightly weighted at one end to represent the disc; and by bending it round a cylindrical object, gently holding at the same time the upper end under the smooth head of a pin, to represent the retarding action of the anther, I then let the lower end suddenly free, and
the whalebone was pitched forward, like the pollinium of the Catasetum, with the weighted end foremost.
That the disc is first jerked out, I ascertained by pressing with a scalpel on the middle of the pedicel and by then touching the antenna; instantaneously out came the disc, but, owing to the pressure on the pedicel, the pollinium was not ejected. Besides the spring from the straightening of the pedicel, elasticity in a transverse direction comes into play: if a quill be split lengthways, and the half be forced longitudinally on a too thick pencil, immediately the pressure is removed the quill jumps off; and an analogous action takes place with the pedicel of the pollinium, owing to the sudden inward curling of its edges. These combined forces suffice to eject the pollinium with considerable force to the distance of two or three feet. Several persons have told me that, when touching the flowers of this genus in their hot-houses, the pollinia have struck their faces. I touched the antennæ of C. callosum whilst holding the flower at about a yard's distance from the window, and the pollinium hit the pane of
glass, and adhered to the smooth vertical surface by its adhesive disc.
The following observations on the nature of the excitement which causes the disc to separate from the surrounding parts, include some made on the two succeeding species. Several flowers were sent me by post and railroad, and must have been much jarred, but had not exploded. I let two flowers fall from a height of two or three inches on the table, but the pollinia were not ejected. I cut off the ovarium close under the flower, and the sepals, and in some cases the thick labellum, with a crash by a pair of scissors; but this violence produced no effect. Nor did deep pricks in various parts of the column even within the stigmatic chamber. A blow, sufficiently hard to knock off the anther suddenly, caused the ejection of the pollinium, as occurred to me once by accident. Twice I pressed rather hard on the pedicel, and consequently on the underlying rostellum, without any effect. Whilst pressing on the pedicel, I gently removed the anther, and then the pollen-bearing end of the pollinium sprang up from its elasticity, and this movement caused
the disc to separate. M. Ménière,* however, has stated that the anther-case sometimes detaches itself, or can be gently detached, without the disc separating, and that then the pedicel swings downwards in front of the stigmatic chamber.
After trials made on fifteen flowers of three species, I find that no moderate degree of violence on any part of the flower, excepting the antennæ, produces any effect. But when the right-hand antenna of C. saccatum, or either antenna of the two following species, is touched, the pollinium is instantly ejected. The extreme tip and the whole length of the antennæ are sensitive. In one specimen of C. tridentatum a touch from a bristle sufficed; in five specimens of C. saccatum a gentle touch from a fine needle was necessary; but in four other specimens a slight blow was requisite. In C. tridentatum a stream of air and of cold water from a small pipe did not suffice; nor in any case did a touch from a human hair; so that the antennæ are less sensitive than the rostellum of Listera. Such extreme sensitiveness would indeed have been useless to the
* 'Bull. de la Soc. Bot. de France,' tom. I. 1854, p. 367.
plant, for we have reason to believe that the flowers are visited by bulky insects.
That the disc does not separate by the simple mechanical movement of the antennæ is almost certain; for the antennæ firmly adhere for a considerable space to the sides of the stigmatic chamber, and are thus immoveably fixed near their bases. The flowers in some cases, when they first arrived, were not sensitive, but after the spikes had stood for a day or two in water they became sensitive. Whether this was owing to fuller maturity or to the absorption of water, I do not know. Two flowers of C. callosum, which were completely torpid, were immersed in tepid water for an hour; and then the antennæ became highly sensitive; this indicates that the cellular tissue of the antennæ must be turgid in order to receive and convey the effects of a touch; and it would lead to the suspicion that a vibration is conveyed along them; if this be so, the vibration must be of some special nature, for the ordinary jars of manifold greater force do not cause the act of rupture. Two flowers placed in hot water, but not so hot as to scald my fingers, spontaneously ejected their pollinia. The loss
of a plant, on which I intended to try other experiments, prevented my ascertaining whether drops of vapour of acrid fluids would act. From these latter facts it may be doubted whether it can be a vibration from the gentle touch of a needle which is conveyed along the antennæ. In C. tridentatum I found that the antennæ were one inch and one-tenth of an inch in length, and a gentle touch from a bristle on the extreme tip was conveyed, as far as I could perceive, instantaneously throughout this length. I measured the length of several cells in the tissue composing the antennæ, and on a rough average it appeared that the stimulus must travel through no less than from seventy to eighty closed cells.
We may, at least, safely conclude that the antennæ, which are characteristic of the genus Catasetum, are specially adapted to receive and convey the effects of a touch to the disc of the pollinium; causing the membrane to rupture, and the whole pollinium to be ejected by its elasticity. If we required further proof, nature has afforded it in the case of the so-called genus Monachanthus, which, as we shall see, is the female plant of Catasetum tridentatum, and
has no pollinia to eject, and here the antennæ are entirely absent.
I have stated that in C. saccatum the right-hand antenna invariably hangs down, with the tip turned slightly outwards, and that it is almost paralysed, I ground my belief on five trials, in which I violently hit, bent, and pricked this antenna, and produced no effect; but immediately afterwards touching the left-hand antenna with much less force, the pollinium was shot forth. In a sixt case a forcible blow on the right-hand antenna did cause the act of ejection, so that it is not completely paralysed. As this antenna does not guard the labellum, which seems in all Orchids to be the part attractive to insects, its sensitiveness would be useless.
From the large size of the flower, more especially of the viscid disc, and from its wonderful power of adhesion, we may safely infer that the flowers are visited by large insects. The viscid matter sticks so firmly when it has set hard, and the pedicel is so strong (though very thin and only one-twentieth of an inch in breadth at the hinge), that to my surprise it supported for a few
seconds a weight of 1262 grains, that is nearly three ounces; and it supported for a considerable time a slightly less weight. When the pollinium is shot forth, the large spike-like anther is generally carried with it. When the disc strikes a flat surface like a table, the momentum from the weight of the anther often carries the pollen-bearing end beyond the disc, and the pollinium is thus affixed in a wrong direction, supposing it to have been attached to an insect's body, for the fertilisation of another flower. The flight is also often rather crooked.* But it must not
* Mr. Baillon ('Bull. de la Soc. Bot. de France,' tom. I. 1854, p. 285) states that Catasetum luridum ejects its pollinia always in a straight line, and in such a direction that it sticks fast to the bottom of the concavity of the labellum: when in this position he imagines that it fertilises the flower in a manner not clearly explained. In a subsequent paper in the same work (p. 367) M. Ménière justly disputes Mr. Baillon's conclusion. He remarks that the anther-case is easily detached, and sometimes naturally detaches itself; in this case the pollinia swing downward by the elasticity of the pedicel, and the viscid disc still remains attached to the roof of the stigmatic chamber. Mr. Ménière then hints that, by the subsequent and progressive retraction of the pedicel, the pollen-masses might be carried into the stigmatic chamber. This is not possible in the three species which I have examined, and would be useless. But M. Ménière himself then goes on to show how important insects are to the fertilisation of Orchids; and apparently he infers that their agency comes into play with Catasetum, and that this plant does not fertilise itself. Both M. Baillon and M. Ménière correctly describe the curved position in which the elastic pedicel lies before it is set free. Neither of these botanists seems to be aware that the species of Catasetum (at least the three which I have examined) are exclusively male plants.
be forgotten that under nature the ejection is caused by the antennæ being touched by a large insect standing on the labellum, which will thus have its head and thorax placed near the anther. A rounded object thus held is always accurately struck in the middle, and, when removed with the pollinium adhering to it, the weight of the anther depresses the hinge of the pollinium; and in this position the anther readily drops off, leaving the balls of pollen free and in a proper position for the act of fertilisation. The utility of so forcible an ejection may be to drive the soft and viscid cushion of the disc against the hairy thorax of a large hymenopterous insect, or the sculptured thorax of a flower-feeding beetle. When attached, assuredly no force which the insect could exert would remove the disc and the pedicel; but the caudicles are ruptured without much difficulty, and thus the balls of pollen would be left on the viscid stigmatic surface of a female flower.
Catasetum callosum.—This species* is smaller than the last, but resembles it in most respects. The edge of the labellum is covered with papillæ; the pit in the middle is small, and behind it there is an elongated anvil-like projection,—facts which I mention from the relation of this form to Myanthus barbatus, presently to be described. The yellow-coloured pedicel is much bowed, and is joined by a hinge to the extremely viscid disc. When either antenna is touched, the pollinia are ejected with much force. The two antennæ stand symmetrically on each side of the anvil-like projection, with their tips lying within the pit of the labellum. The walls of this pit have a pleasant nutritious taste. The antennæ are remarkable, from their whole surface being roughened with papillæ. The plant is a male.
Catasetum tridentatum.—The general appearance of this species, which is very different from that of the two former species, is represented by Fig. XXVII., with a sepal on each side cut off.
* A fine spike of flowers of this species was most kindly sent me by Mr. Rucker, and was named for me by Dr. Lindley.
Fig. XXVII.
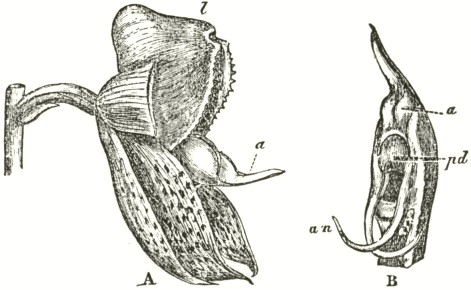
CATASETUM TRIDENTATUM.
a. anther.
pd. pedicel of pollinium.
an. antennæ.
l. labellum.
A. Side view of flower in its natural position with the (properly) lower sepals cut off.
B. Front view of column, placed upright.
The flower stands with the labellum uppermost, that is in a reversed position compared with most Orchids. The labellum forms a helmet or bucket, with the distal portion represented by three minute points. It is clear from this position that the labellum cannot hold nectar; but its walls are thick, and have, as in the other species, a pleasant nutritious taste. The stigmatic chamber, though functionless as a stigma, is of large
size. The summit of the column, and the spike-like anther, are not so much elongated as in C. saccatum. In other respects there is no important difference. The antennæ are of greater length; their tips for about one-twentieth of their length are roughened by cells produced into papillæ.
The pedicel of the pollinium is articulated as before by a hinge to the disc; the anterior end of the disc is upturned, so that, when attached to an insect's head, the pedicel cannot move backwards, only forwards,—a movement which apparently comes into play in the fertilisation of the female plant. The disc is, as in the other species, of large size, and its posterior end, which during ejection first strikes any object, is much more viscid than the rest of the surface. This latter surface is drenched with a milky fluid, which rapidly turns brown when exposed to the air, and sets into a cheesy consistence. The upper surface of the disc consists of strong membrane formed of polygonal cells, each containing one or several balls of brown translucent matter. This membrane rests on, and adheres to, a thick cushion, formed
of rounded balls of brown matter (which lower down in the cushion become extremely irregular in shape) separated from each other and embedded in transparent, structureless, highly elastic matter. This cushion towards the posterior end of the disc passes into the extremely viscid matter. This latter matter when consolidated is brown, translucent, and homogenous. Altogether the disc presents a much more complex structure than in the other Vandeæ.
I need not further describe this species, excepting the position of the antennæ. They occupied exactly the same position in the six flowers examined. They do not stand symmetrically. They are both sensitive,—whether in an equal degree I will not say. They both lie curled within the bucket-like labellum; the left-hand one stands higher up, with its inwardly bowed extremity in the middle; the right-hand antenna lies lower down and crosses the whole base of the labellum, with its tip just projecting beyond the left margin of the base of the column. From the position of the petals and sepals, an insect visiting the flower would almost certainly alight on the
crest of the labellum; but it could hardly gnaw any part of the great cavity of the labellum without touching one of the two antennæ, for the left-hand one guards the upper part, and the right-hand one the lower part: and when these are touched the pollinium will infallibly be ejected and strike the head or thorax of the insect.
The position of the antennæ in this Catasetum may be compared with that of a man with his left hand raised and bent so that his hand stands in front of his chest, and with his right arm crossed lower down so that the fingers project just beyond his left side. In Catasetum callosum both arms are held lower down, and are extended symmetrically. In C. saccatum the left arm is bowed and held in front, as in the C. tridentatum, but rather lower down; whilst the right arm hangs down almost paralysed, with the hand turned a little outwards. In every case notice will be given in an admirably manner, when an insect visits the labellum, and the time has at last arrived for the ejection of the pollinium and for its transportal to the female plant.
Catasetum tridentatum is interesting under another point of view. Botanists were astonished when Sir Robert Schomburgk* stated that he had seen three forms, believed to constitute three distinct genera, namely, Catasetum tridentatum, Monachanthus viridis, and Myanthus barbatus, all growing on the same plant. Lindley remarked† that "such cases shake to the foundation all our ideas of the stability of genera and species." Sir. R. Schomburgk affirms that he has seen hundreds of plants of C. tridentatum in Essequibo without ever finding one specimen with seeds, ‡ but that he was surprised at the gigantic seed-
* 'Transactions of the Linnæan Soc.' Vol. xvii. P. 522. Another account by Dr. Lindley appeared in the 'Botanical Register,' fol. 1951, of a distinct species of Myanthus and Monachanthus appearing in the same scape: he alludes also to other cases. Some of the flowers were in an intermediate condition, which is not surprising, seeing that in diœcious plants we sometimes have a partial resumption of the characters of both sexes. Mr. Rodgers of Riverhill informs me that he imported from Demerara a Myanthus, but that when it flowered a second time it was metamorphosed into a Catasetum. Dr. Carpenter ('Comparative Physiology,' 4th edit. P. 633) alludes to an analogous case which occurred at Bristol.
† The 'Vegetable Kingdom, 1853, p. 178.
‡ Brongniart states ('Bull. de la Soc. Bot. de France, ' tom. ii. 1855, p.20) that Mr. Neumann, a skilful fertiliser of Orchids, could never succeed in fertilising Catasetum.
vessels of the Monachanthus; and he correctly remarks that "here we have traces of sexual difference in Orchideous flowers."
From what I had myself previously observed, I was led to examine carefully the female organs of C. tridentatum, callosum, and saccatum. In no case was the stigmatic surface viscid, as it is in all other Orchids (except was we shall see in Cypripedium), and as is indispensable for the securing the pollen-masses by the rupture of the caudicles; I carefully looked to this point both in young and old flowers of C. tridentatum. When the surface of the stigmatic chamber and of the stigmatic canal of the above-named three species is scraped off, after having been kept in spirits, it is found to be composed of utriculi, with nuclei of the proper shape, but not nearly so numerous as with ordinary Orchids. The utriculi cohere more together and are more transparent; I examined for comparison the utriculi of many kinds of Orchids which had been kept in spirits, and in all found them much less transparent. In C. tridentatum, the ovarium is shorter, much less deeply furrowed, narrower at the base, and
internally more solid than in the Monachanthus. Again, in all three species of Catasetum the ovule-bearing cords are short; and the ovules present a considerably different appearance, in being thinner, more transparent; and less pulpy than in the numerous other Orchids examined for comparison. They were, however, in not so completely and atrophied condition as in Acropera. Although they correspond so closely in general appearance and position with true ovules, perhaps I have no strict right so to designate them, as I was unable in any case to make out the opening of the testa and the included nucleus; nor were the ovules ever inverted.
From these several facts, namely,—the shortness, smoothness, and narrowness of the ovarium, the shortness of the ovule-bearing cords, the state of the ovules themselves, the stigmatic surface not being viscid, the empty condition of the utriculi,—and from Sir R. Schomburgk never having seen C. tridentatum producing seed in its native home, we may confidently look at this species, as well as the other two species of Catasetum, as male plants.
Fig. XXVIII.
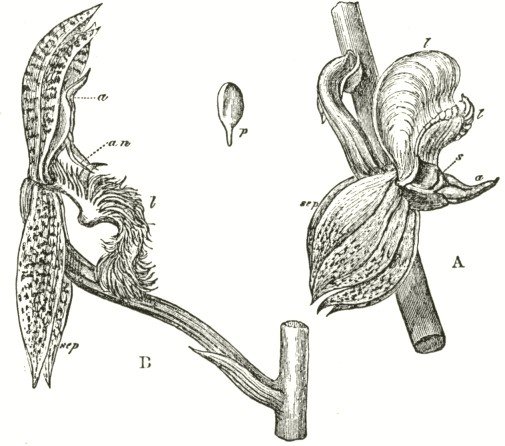
MYANTHUS BARBATUS. MONACHANTHUS VIRIDIS.
a. anther.
an. antennæ.
l. labellum.
p. pollen-mass, rudimentary.
s. stigmatic cleft. sep. two lower sepals.
A. Side view of Monachanthus viridis in its natural position. (The shading in both drawings has been added from M. Reiss' drawing in the 'Linnæan Transactions.')
B. Side view of Myanthus barbatus in its natural position.
With respect to Monachanthus viridis and Myanthus barbatus, the President and officers of the Linnæan Society have kindly permitted me to examine the spike bearing these two flowers, preserved in spirits, and sent home
by Sir R. Schomburgk. They are here represented (Fig. XXVIII.). The flower of the Monachanthus, like that of the Catasetum, grows lower side uppermost. The labellum is not nearly so deep, especially on the sides, and its edge is crenated. The other petals and sepals are all reflexed, and are not so much spotted as in Catasetum. The bract at the base of the ovarium is much larger. The whole column, especially the filament and the spike-like anther, are much shorter; and the rostellum is much less protuberant. The antennæ are entirely absent, and the pollen-masses are rudimentary. These are interesting facts, from corroborating the view taken of the function of the antennæ; for as there are no proper pollinia to eject, there could be no use in an organ to convey the stimulus from a touch to the rostellum. I could find no trace of a viscid disc or pedicel; if they exist, they must be extremely rudimentary, for there is hardly any space for the embedment of the disc.
Instead of a large stigmatic chamber, there is a narrow transverse cleft close beneath the small anther. I was able to insert one of the
pollen-masses of the male Catasetum into this cleft, which from having been kept in spirits was lined with coagulated beads of viscid matter, and with utriculi. The utriculi, differently from those in Catasetum, were charged (after having been kept in spirits) with brown matter. The ovarium is longer, thicker near the base, and more plainly furrowed than in Catasetum; the ovule-bearing cords are also much longer, and the ovules more opaque and pulpy, as in all common Orchids. I believe that I saw the opening at the partially inverted end of the testa, with a large nucleus projecting; but as the specimens had been kept many years in spirits and were somewhat altered, I dare not speak positively. From these facts alone it is almost certain that Monachanthus is a female plant; and Sir. R. Schomburgk saw it seeding abundantly. Altogether this flower differs in a most remarkable manner from that of the male Catasetum tridentatum, and it is no wonder that they were formerly ranked as distinct genera.
The pollen-masses offer so curious and good an illustration of a structure in a rudimentary condition that they are worth description;
but first I must briefly describe the perfect pollen-masses of the male Catasetum. These may be seen at D and E, Fig. XXVI. , attached to the pedicel: they consist of a large sheet of cemented or waxy pollen-grains, folded over so as to form a sack, with an open slit (E) along the lower surface, into this slit cellular tissue enters whilst the pollen is in the course of development. Within the lower and produced end of each pollen-mass a layer of highly elastic tissue, forming the caudicle, is attached; the other end being attached to the pedicel of the rostellum. The exterior grains of pollen are more angular, have ticker walls, and are yellower than the interior grains. In the early bud the two pollen-masses are enveloped in two conjoined membranous sacks, which are soon penetrated by the two produced ends of the pollen-masses and by their caudicles; and then the ends of the caudicles adhere to the pedicel. Before the flower expands the membranous sacks including the two pollen-masses open, and leave them resting naked on the back of the rostellum.
In Monachanthus the two membranous sacks containing the rudimentary pollen-masses on
the contrary never open; they easily separate from each other and from the anther. The tissue of which they are formed is thick and pulpy. Like most rudimentary parts they vary greatly in size and form. The included and therefore useless pollen-masses are not one-tenth of the bulk of the pollen-masses of the male; they are flask-shaped (p, Fig. XXVIII.), with the lower and produced end greatly exaggerated and almost penetrating through the exterior or membranous sack. The flask is closed, and there is no fissure along the lower surface. The exterior pollen-grains are square and have thicker walls than the interior grains, just as in the proper male pollen; and, what is very curious, each cell has its nucleus. Now, R. Brown has stated* that in the early stages of the formation of the pollen-grains in ordinary Orchids a minute areola or nucleus is often visible; so that the rudimentary pollen-grains of the Monachanthus apparently have retained—as is so general with rudiments in the animal kingdom—an embryonic character. Lastly, at the base, within the flask of pollen, there
* 'Transactions of the Linnæan Soc.' Vol. xvi. p. 711.
is a little mass of brown elastic tissue,—that is, a vestige of a caudicle,—which runs far up the pointed end of the flask, but does not (at least in some of the specimens) come to the surface, and could not have been attached to any part of the rostellum. These rudimentary caudicles are, therefore, utterly useless.
We thus see that every single detail of structure which characterises the male pollen-masses is represented, with some parts exaggerated and some parts slightly modified, by the mere rudiments in the female plant. Such cases are familiar to every observer, but can never be examined without renewed interest. At a period not far distant, naturalists will hear with surprise, perhaps with derision, that grave and learned men formerly maintained that such useless organs were not remnants retained by the principle of inheritance at corresponding periods of early growth, but were specially created and arranged in their proper places like dishes on a table (this is the comparison of a distinguished naturalist) by an Omnipotent hand "to complete the scheme of nature."
We now come to the third form, Myanthus
barbatus (Fig. XXVIII., B), often borne on the same plant with the two preceding forms. Its flowers, in external appearance, but not in essential structure, is the most different of all three. It generally stands in a reversed position, compared with Catasetum and Monachanthus, that is, with the labellum downwards. The labellum is fringed in an extraordinary manner with long papillæ; it has a quite insignificant medial cavity, at the hinder margin of which a curious curved and flattened horn projects. The other petals and sepals are spotted and elongated, with the two lower sepals alone reflexed. The antennæ are not so long as in the male C. tridentatum, and they project symmetrically on each side of the horn-like projection at the base of the labellum, with their tips, which are not roughened with papillæ, almost entering the medial cavity. The stigmatic chamber is of nearly intermediate size between that of the male and female forms: it is lined with utriculi charged with brown matter. The straight and well-furrowed ovarium is nearly twice as long as in Monachanthus, but it is not so thick where it joins
the flower; the ovules are not so numerous as in the female form, but are opaque and pulpy after having been kept in spirits, and resemble them in all respects. I believe, but dare not, as in the case of the Monachanthus, speak positively, that I saw the nucleus projecting from the testa. The pollinia are about a quarter of the size of those of the male Catasetum, but have a perfectly well developed disc and pedicel. The pollen-masses were lost in the specimens examined by me; but fortunately Mr. Reiss has given, in the Linnæan Transactions, a drawing of them, showing that they are due proportional size and have the proper folded or cleft structure: so that there can hardly be a doubt that they were functionally perfect. As we thus see that both male and female organs are apparently perfect, Myanthus barbatus may be considered as the hermaphrodite form of the same species, of which the Catasetum is the male and Monachanthus the female.
It is not a little singular that the hermaphrodite Myanthus should resemble in its whole structure much more closely the male form of two distinct species (namely, C.
saccatum, and more especially C. callosum) than either its own male or female form.
Finally, the genus Catasetum is interesting in an unusual degree in several respects. The separation of the sexes is unknown in other Orchids, excepting as we shall see probably in the allied genus Cycnoches and in the before-given case of Acropera. In Catasetum we have three sexual forms, generally borne on separate plants, but sometimes mingled together; and these three forms are wonderfully different from each other, much more different than, for instance, a peacock is from a peahen. But the appearance of these three forms on the same plant now ceases to be an anomaly, and can no longer be viewed as an unparalleled instance of variability.
Still more interesting is this genus in its mechanism for fertilisation. We see a flower patiently waiting with its antennæ stretched forth in a well-adapted position, ready to give notice whenever an insects puts its head into the cavity of the labellum. The female Monachanthus, not having pollinia to eject, is destitute of antennæ. In the male and hermaphrodite forms, namely Catasetum tri-
dentatum and Myanthus, the pollinia lie doubled up, like a spring, ready to be instantaneously shot forth when the antennæ are touched; the disc end is always projected foremost, and is coated with viscid matter which quickly sets hard and firmly affixes the hinged pedicel to the insect's body. The insect flies from flower to flower, till at last it visits a female or hermaphrodite plant: it then inserts one of the masses of pollen into the stigmatic cavity. When the insect flies away the elastic caudicle, made weak enough to yield to the viscidity of the stigmatic surface, breaks, and leaves behind the pollen-mass; then the pollen-tubes slowly protrude, penetrate the stigmatic canal, and the act of fertilisation is completed. Who would have been bold enough to have surmised that the propagation of a species should have depended on so complex, so apparently artificial, and yet so admirable an arrangement?
I have seen two other genera belonging to the sub-family of Catasetidæ, namely Mormodes and Cycnoches; but the latter arrived in a broken condition.
Mormodes ignea.—To show how difficult it sometimes is to understand the manner of fertilisation of Orchids, I may state that I had carefully examined twelve flowers,* trying various experiments and recording the results, before I could at all make out the meaning and action of the several parts. It was plain that the pollinia were ejected, as in Catasetum, but how each part of the flower played its proper part I could not even conjecture. I had given up the case as hopeless, until, summing up my observations, the explanation presently to be given, and subsequently proved by repeated experiments to be correct, suddenly occurred to me.
The flower presents an extraordinary appearance, and its mechanism is more curious even than its appearance (Fig. XXIX.). The base of the column is bent baclwards, at right angles to the ovarium or footstalk, and then resumes an upright position to near its summit, where it is again bent. It is, also, twisted in a unique manner, so that its front
* I must express my cordial thanks to Mr. Rucker, of West Hill, Wandsworth, for having lent me a plant of this Mormodes with two fine spikes, bearing an abundance of flowers, and for having allowed me to keep the plant for a considerable time.
Fig. XXIX.
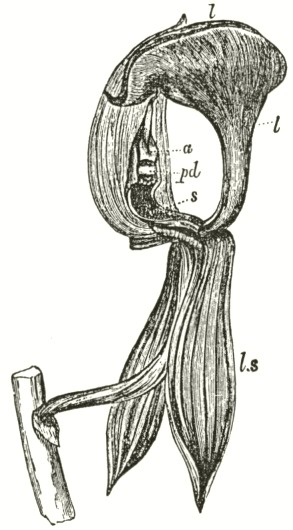
MORMODES IGNEA.
Lateral view of flower, with the upper sepal and the near upper petal cut off.
N.B. The labellum in the drawing is a little lifted up, to show the depression in its surface, which ought to be pressed close down on the bent summit of the column.
a. anther.
pd. pedicel of pollinium.
s. stigma.
l. labellum.
l s. lower sepal.
surface, including the anther, rostellum, and the upper part of the stigma, faces laterally to either the right or left hand in the flowers on the opposite sides of the spike. The twisted stigmatic surface, in an equally peculiar manner, extends down to the base of the column: at its upper end it forms a deep cavity, beneath the protuberant rostellum (pd in the drawing), in which the large viscid disc of the pollinium is lodged.
The anther-case (a in the drawing) is elongated and triangular, closely resembling that of Catasetum: it does not extend up to the apex of the column. This apex consists of a thin flattened filament, which in the bud is straight, but before the flower expands becomes much more bent by the pressure of the labellum. A group of spiral vessels runs up the column as far as the summit of the anther-case; they are then reflexed and run some way down the anther-case. The point of reflexion forms a short thread-like hinge by which the notched top of the anther-case is articulated to the column close beneath its bent summit. The hinge, although less than
a pin's head in size, is of paramount importance, for it is sensitive and conveys the stimulus from a touch to the disc of the pollinium: it serves also as a guide in the act of ejection. As it conveys the necessary stimulus to the disc, one may suspect that part of the tissue of the rostellum, which lies in close contact with the filament of the anther, runs up to this point; but I could detect no difference in structure in comparison with the same parts in Catasetum. The cellular tissue round the hinge is gorged with fluid, for a large drop exudes when the anther is torn off during the ejection of the pollinium. This gorged condition may perhaps facilitate the ultimate rupture of the hinge.
The pollinium does not differ much from that of Catasetum (see Fig. XXVI. D): it lies curved round the rostellum, which is less protuberant than in that genus. The upper and broad end of the pedicel, however, extends beneath the pollen-masses, and they are attached by rather weak caudicles to a medial crest on its upper surface.
The viscid surface of the large disc
lies in contact with the roof of the stigmatic cavity, so that it cannot be touched by insects. The anterior end of the disc is furnished with a small dependent curtain (dimly shown in Fig. XXIX.); and this, before the act of ejection, is continuously joined on each side to the upper margins of the stigmatic depression. The pedicel is united to the posterior end of the disc; but when the disc is freed, the lower part of the pedicel becomes doubly bent, so that it then appears as if attached to the centre of the disc by a hinge.
The labellum is truly remarkable: it is narrowed at its base into a nearly cylindrical footstalk, and its sides are so much reflexed as almost to meet at the back. After rising up perpendicularly it forms an arch, over and behind the summit of the column, against which it is firmly pressed. The labellum at this point (even in the bud) is depressed into a slight cavity, which receives the ultimately bent summit of the column. This slight depression manifestly represents the large hollow, with thick fleshy walls, in the labellum of the several species of Catasetum
and other Vandæ, which serves to attract insects; here, by a singular change of function, it merely keeps the labellum in its proper position on the summit of the column. In the drawing (Fig. XXIX.) the labellum is represented as forcibly raised so as to show the depression and the bent filament. The labellum in its natural position may almost be compared to a huge cocked-hat, supported by a footstalk and placed on the head of the column.
The twisting of the column, which I have seen in no other Orchid, causes all the important organs of fructification to face to the left in the flowers on the left side of the spike, and to face to the right in all those on the right side. So that two flowers taken from opposite sides of the spike and held in the same relative position are seen to be twisted in opposite directions. One single flower, which was crowded by the others, was barely twisted, so that its column faced the labellum. The labellum is also slightly twisted: for instance, in the flower figured which faces to the left, the midrib of the labellum first bends to the right hand, and
then back again, but in a lesser degree, to the left, so as to press against the posterior surface of the summit of the column. The twisting of all the parts of the flowers commences in the bud.
The position thus acquired by the several organs is of the highest importance; for if the column and labellum had not been twisted laterally, the pollinia, when shot forth, would have struck the overarching labellum and been thrown back, as actually occurred with the single abnormal flower having a nearly straight column. If all the organs had not been twisted in opposite directions on the two sides of the crowded spike, so as to face always to the outside, there would not have been a clear space for the ejection of the pollinia and their adhesion to insects.
When the flower is mature the three sepals hang down, but the two upper petals remain nearly upright. The bases of the sepals, and especially of the two upper petals, are thick and swollen and have a yellowish tint; they are so gorged, when quite mature, with fluid, that, when punctured by a fine glass tube, the
fluid rises to some height in it by capillary attraction. These swollen bases, as well as the footstalk of the labellum, have a decidedly sweet and pleasant taste; and I can hardly doubt that they are attractive to insects, for no free nectar is secreted.
I will now endeavour to show how all the parts of the flower are co-ordinated and act together. The pollinium, as in Catasetum, lies bowed round the rostellum; in that genus when freed it merely straightens itself with force, in this Mormodes something more takes place. If the reader will look forward to Fig. XXX., he will see a section of the flower-bud of another species of Mormodes, which differs only in the shape of the anther and in the viscid disc having a much deeper dependent curtain. Now let him suppose the pedicel of the pollinium to be so elastic that when freed it not only straightens itself, but suddenly bends backwards with a reversed curvature, so as to form an irregular hoop; he will then see that the exterior surface of the curtain, which is not viscid, will lie on the anther-case, and the viscid surface of the disc will be on the outside of the hoop.
This is exactly what takes place with our present Mormodes. But the pollinium assumes with such force its reversed curvature (aided, apparently, by a transverse curling outwards of the two margins of the pedicel), that it instantaneously rebounds from the protuberant face of the rostellum. As the two pollen-masses adhere, at first, rather firmly to the anther-case, the latter is torn off by its base to be rebound; and as the thin hinge at the summit of the anther-case does not at first break, the pollinium with the anther-case is instantly swung upwards like a pendulum: but in the course of the upwards swing the hinge yields and the whole body is projected perpendicularly up in the air, an inch or two above and close in front of the terminal part of the labellum. When no object is in the way, and the pollinium falls down, it generally alights and sticks, though not firmly, in the fold on the crest of the labellum, directly over the column. I witnessed repeatedly all that has been here described.
The curtain of the disc, which, after the pollinium has formed itself into a hoop, lies
on the anther-case, is of considerable service in preventing the viscid matter from the disc adhering to the anther, and thus permanently retaining the pollinium in the form of a hoop. This would have been fatal, as we shall presently see, to a subsequent movement in the pollinium necessary to the fertilisation of the flower. In some of my experiments, when the free action of the parts was checked, this did occur, and the pollinium, with the anther, remained glued together in the shape of an irregular hoop.
I have already stated that the minute hinge by which the anther-case is articulated to the column, a little way beneath its bent filamentary apex, is sensitive to the touch. I tried four times and found that I could touch with some force any other part; but when I gently touched this point with the finest needle, instantaneously the membrane which unites the disc to the edges of the cavity in which it is lodged, ruptured, and the pollinium was shot upwards and alighted on the crest of the labellum as just described.
Now let us supposse an insect to alight on the crest of the labellum (and no other con-
venient landing-place is afforded), and then to lean over in front of the column to gnaw or suck the bases of the petals swollen with sweet fluid. The weight and movements of the insect would press and move the labellum together with the bent underlying summit of the column; and the latter, pressing on the hinge in the angle, would cause the ejection of the pollinium, which would infallibly strike the head of the insect and adhere to it. I tried by placing my gloved finger on the summit of the labellum, with the tip just projecting beyond its margin, and then gently moving my finger it was really beautiful to see how instantly the pollinium was projected upwards, and how accurately the whole viscid surface of the disc struck my finger and firmly adhered to it. Nevertheless, I doubt whether the weight and movement of an insect would suffice to thus act directly on the sensitive point; but look at the drawing and see how probable it is that an insect leaning over would place its front legs over the edge of the labellum on the summit of the anther-case, and would thus touch the sensitive point; the pollinium
would then be ejected, and the viscid disc would certainly strike and adhere to the insect's head.
Before proceeding, it will be worth while to mention some of the early trials which I made. I pricked deeply the column in different parts and the stigma, and cut off the petals, and even the labellum, without causing the ejection of the pollinium; once, however, in cutting rather roughly through the thick footstalk of the labellum, the ejection ensued, no doubt caused by the filamentary summit of the column being disturbed. When I gently prised up the anther-case at its base or at one side, the pollinium was ejected, but then the sensitive hinge must necessarily have been bent. When the flower has long remained expanded and is nearly ready for spontaneous ejection, a slight jar on any part of the flower causes the action. Pressure on the thin pedicel, and therefore on the underlying protuberant rostellum at the basal edge of the anther-case, is followed by the ejection of the pollinium; but this is not surprising, as the stimulus from a touch on the sensitive hinge has to be conveyed through this part
of the rostellum to the disc. In Catasetum slight pressure on this point does not cause the ejection; but in this genus the protuberant part of the rostellum does not lie in the line of conveyed stimulus between the antennæ and the disc. A drop of chloroform, of spirits of wine, and of boiling water placed on this part of the rostellum produced no effect; nor, to my surprise, did exposure of the whole flower to vapour of chloroform.
Seeing that this part of the rostellum was sensitive to pressure, and that the flower was laterally widely open, and being pre-occupied with the case of Catasetum, I at first felt convinced that insects entered the lower part of the flower and touched the rostellum. Accordingly I pressed the rostellum with variously-shaped objects, but the viscid disc never once well adhered to the object. If I used a thick needle, the pollinium, when ejected, formed a hoop round it with the viscid surface outside; if I used a broad flat object, the pollinium struggled against it and sometimes coiled itself up spirally, but the disc either did not adhere at all or very imperfectly. At the close of
the twelfth trial I was in despair. The strange position of the labellum, perched on the summit of the column, ought to have shown me that here was the place for experiment. I ought to have scorned the notion that the labellum was thus placed for no good purpose: I neglected this plain guide, and for a long time completely failed to understand the structure of the flower.
We have seen that when the pollinium is freely ejected upwards it adheres by the whole viscid surface of the disc to any object projecting beyond the edge of the labellum directly over the column. When thus attached, it forms an irregular hoop, with the torn-off anther-case still covering the pollen-masses and lying in close contact with the disc, but protected from adhering to it by the curtain. Whilst in this position the projecting and bowed pedicel would effectually prevent the pollen-masses from being placed on the stigma of a flower, even supposing the anther-case to have fallen off. Now let us suppose the pollinium to be attached to an insect's head, and observe what takes place. The pedicel when first separated by the act
of ejection from the rostellum is quite wet on its inferior surface: as this surface dries, the pedicel slowly straightens itself, and when perfectly straight the anther-case readily drops off. The pollen-masses are now naked, and they are attached by easily ruptured caudicles to the end of the pedicel, at the right distance and on that surface which would naturally be placed in contact with the viscid stigma when the insect visited another flower, so that every detail of structure is now perfectly well adapted for the act of fertilisation.
When the anther-case drops off, it has performed its triple function; namely, its hinge as an organ of sense, its weak attachment to the column as a guide causing the pollinium to be at first swung perpendicularly upwards, and its lower margin, together with the curtain of the disc, as a protection to the pollen-masses from being permanently glued to the viscid disc.
From observations made on fifteen flowers, the straightening of the pedicel takes from about twelve to fifteen minutes. The first movement causing the act of ejection is due to elasticity; the second slow movement,
which counteracts the first, is due to the drying of the outer and convex surface; but this movement differs from that observed in the pollinia of so many Vandeæ and Ophreæ, for, when the pollinium of this Mormodes is placed in water, it does not recover the hoop-like form which it had acquired by elasticity.
Mormodes ignea is an hermaphrodite. The pollinia are well developed. The curiously elongated stigmatic surface is extremely viscid and abounds with innumerable utriculi, the contents of which shrink and become coagulated after immersion for less than an hour in spirits of wine. When placed in spirits for a day, the utriculi are so acted on that they disappear, and this I have noticed in no other Orchid. The ovules, after exposure to spirits for a day or two, present the usual semi-opaque, pulpy appearance common to all hermaphrodite and female Orchids. From the unusual length of the stigmatic surface I expected that, if the pollinia were not ejected from the excitement of a touch, the anther-case would detach itself, and that the pollen-masses would swing downwards and
fertilise their own flower. Accordingly, I left four flowers untouched; after remaining expanded from eight to ten days, the elasticity of the pedicel conquered the force of attachment and the pollinia were spontaneously ejected, but they were always wasted.
Although Mormodes ignea is an hermaphrodite, yet it must be, in fact, as truly dicious as Catasetum as far as the concurrence of two individuals is concerned in the act of reproduction; for, as it takes from twelve to fifteen minutes, after the act of ejection, before the pedicel of the pollinium straightens itself and the anther-case drops off, it is almost certain that an insect with the pollinium attached to its head would within this time leave one plant and fly to another.
A second species of Mormodes, sent me, unnamed, by Mr. Veitch, is very different from M. ignea both in general appearance and in structure. The yellow petals and sepals are reflexed; the thick labellum is singularly shaped, with its upper surface convex, like a shallow basin turned upside
down. The thin column is of extraordinary length, and arches over the labellum. The appearance of the flower is like that of Cycnoches represented on the cover of this volume; but the column in the drawing is too much foreshortened.
The specimens unfortunately arrived broken; but some large flower-buds, of which a section is here given, show the essential structure. We see the elastic pedicel of the pollinium bowed as in the last species; but at the period of growth here represented, the pedicel was still united to the rostellum, the future line of separation being shown by a layer of hyaline tissue, indistinct towards the upper end of the disc. The disc is of gigantic size, and its lower end is produced into a great fringed curtain, which hangs in front of the stigmatic chamber. The adhesion of this disc, when mature, to any object is surprisingly strong. The margins of the stigmatic chamber on each side are slightly protuberant; and these protuberances, like the antennæ in Catasetum, are continuous with the rostellum. The anther is widely different in shape from that in the last species and in Catasetum, and apparently
would retain the pollen-masses with more force.
Fig. XXX.
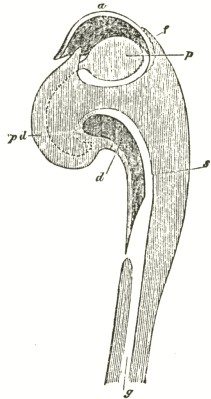
SECTION OF THE FLOWER-BUD OF A MORMODES.
a. anther.
f. filament of anther.
p. pollen-mass.
pd. pedicel of pollinium, barely separated from the rostellum.
d. disc of pollinium.
s. stigmatic chamber.
g. stigmatic canal leading to the ovarium.
From information given me by Mr. Veitch it appears that, when the end of the column is touched, some movement takes place; and, from the analogy of Catasetum, it is probable that the protuberances on the margin of the
stigmatic chamber are sensitive. These protuberances may be provisionally considered as nascent antennæ. It is obvious, from the section given, that the disc, as long as its viscid surface rests against the roof of the stigmatic chamber, cannot adhere to any object; but Mr. Veitch informs me that, when he has touched the end of the column, the disc has adhered to his fingers. These facts perhaps suffice to show what takes place under nature. A large insect visits the thick and fleshy labellum, which is overarched by the column, and touches with its back the sensitive and protuberant edges of the stigmatic chamber: when thus excited the disc alone is flirted out and adheres to the insect's back; the insect flies away, drags the pollen-masses from under the anther, and carries them to another flower. The insect, standing in the same position, inserts the pollen-masses into the stigmatic chamber, and the flower is fertilised.
When I received the flowers of this second species of Mormodes and of Cycnoches, I did not examine their ovaria and stigmas, for I knew nothing at that time about the sexual
forms of Catasetum. Mormodes ignea is an hermaphrodite; if the second species is an hermaphrodite, it cannot be fertilised, owing to the size of the curtain of the disc, until its own pollinium has been removed. With respect to Cycnoches, it is known, from an account published by Lindley,* that C. ventricosum produces on the same scape flowers with a simple labellum, others with a much segmented labellum, and others in an intermediate condition. From the analogous differences in the labellum of the sexes in Catasetum, we may believe that we here see the male, female, and hermaphrodite forms of Cycnoches.
Of Lindley's fourth and sixth Tribes, viz. The Ophreæ and Neotteæ, plenty of British forms have been described. Of the fifth Tribe, the Arethuseæ, I have not seen any living flowers. Judging from statements with respect to three distantly allied forms in this tribe, mechanical aid is requisite for their fertilisation. Irmisch makes this remark
* 'Vegetable Kingdom,' 1853, p. 177. Lindley has also published in the 'Botanical Register,' fol. 1951, a similar case of the production of two forms on the scape in another species of Cycnoches.
with respect to Epipogium aphyllum.* Mr. Rodgers, of Sevenoaks, informs me that in his hot-house species of Limodorum did not set their fruit without aid; and this is likewise well known to be the case with the Vanilla. This latter genus is cultivated for its aromatic pods in Tahiti, Bourbon, and the East Indies; but does not fruit without artificial aid.† This fact shows that some insect in its own American home is specially adapted for its fertilisation; and that the insects of the above-named tropical regions, where the Vanilla flourishes, either do not visit the flowers, though they secrete an abundance of nectar, or do not visit them in the proper method.
We have now arrived at Lindley's last and seventh Tribe, including only one genus, Cypripedium, which differs from all other Orchids far more than any other two do from each other. An enormous amount of
* 'Beiträge zur Biologie der Orchideen,' 1853, s. 55.
† For Bourbon see 'Bull. Soc. Bot. de France,' tom. i. 1854, p. 290. For Tahiti see H. A. Tilley, 'Japan, the Amour, &c.,'1861, p. 375. For the East Indies see Morren in Annals and Mag. of Nat. Hist., 1839, vol. iii. p. 6.
extinction must have swept away a multitude of intermediate forms, and left this single genus, now widely disseminated, as a record of a former and more simple state of the great Orchidean Order. Cypripedium possesses no rostellum; all three stigmas being fully developed, but confluent. That anther, which is present in all other Orchids, is here rudimentary, and is represented by a singular shield-like projecting body, deeply notched or hollowed out on its lower margin. There are two fertile anthers which belong to an inner whorl, represented in ordinary Orchids by various rudiments. The pollen-grains do not consist of three or four united granules, as in all other genera, excepting the degraded Cephalanthera. The grains are not united into waxy masses, nor tied together by elastic threads, nor furnished with a caudicle. The labellum is of large size, and as in all other Orchids is a compounded organ.
The following remarks apply to the four species which I have seen, namely, C. barbatum, purpuratum, insigne, and venustum. The manner of fertilisation is here widely different from that in all the many
before-given cases. The labellum is folded round the short column, so that its edges nearly meet along the dorsal surface; and its broad extremity is folded over and backwards in a peculiar manner, so as to form a
Fig. XXXI.
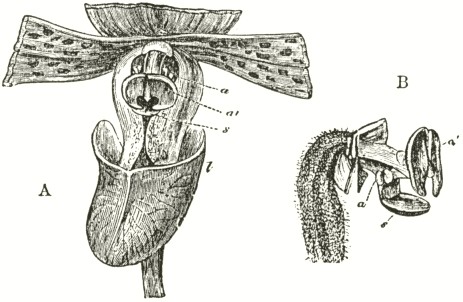
CYPRIPEDIUM.
a. anther.
a'. rudimentary, shield-like anther.
s. stigma.
l. labellum.
A. Flower viewed from above, exhibiting the dorsal surface, with the sepals and petals, excepting the labellum, partly cut off. The labellum is slightly depressed, so that the dorsal surface of the stigma is brought outside; the edges of the labellum thus become a little separated, with the toe depressed.
B. Side view of column, with all the sepals and petals removed.
sort of shoe, which closes up at the end of the flower. Hence arises the English name of Ladies'-slipper. In the position in which the
flower grows, and as it is here represented, the dorsal surface, with the edges of the labellum almost meeting, is uppermost. The stigmatic surface is slightly protuberant, and is not viscid; it fronts the basal surface of the labellum. The margin of the upper and dorsal surface of the stigma can be barely distinguished between the edges of the labellum and through the notch in the rudimentary, shield-like anther (a'); but in the drawing (s, Fig. A) this margin is brought outside the edges of the labellum by their depression; the toe also of the labellum is a little bent down, so that the flower is represented as rather more open than it really is. The edges of the pollen-masses of the two lateral anthers (a) can be seen lying down within the labellum and projecting a little beyond the column. The grains of pollen are immersed in, and coated by, viscid fluid, which is so glutinous that it can be drawn out into threads. As the two anthers stand behind and above the lower convex surface (see Fig. B) of the stigma, it is impossible that the glutinous pollen can get on to this, the fertile surface, without mechanical aid.
An insect could reach the extremity of the labellum, or the toe of the slipper, through the longitudinal dorsal slit; but according to all analogy the basal portion in front of the stigma would be the most attractive part. Now, as the flower is closed at the end, owing to the toe of the labellum being upturned, and as the dorsal surface of the stigma, together with the large shield-like rudimentary anther, almost close the basal part of the medial slit, two convenient passages alone are left for an insect to reach with its proboscis the lower part of the labellum; namely, directly over and close outside the two lateral anthers. If an insect where thus to act, and it could hardly act in any other way, it would infallibly get its proboscis smeared with the glutinous pollen, as I found to occur with a bristle thus inserted. When the bristle smeared with pollen was pushed further into the flower, especially if pushed in by the little notch outside the anther, some of the glutinous pollen was generally left on the slightly convex stigmatic surface. The proboscis of an insect would effect this latter operation better than a bristle, owing to its flexibility
and power of movement. Thus an insect would place either the flower's own pollen on to the stigma, or, flying away, would carry the pollen to another flower. Which of these two contingencies commonly occurs, will depend on whether the insect first inserts its proboscis directly over the anther, or outside by the little notch.
We thus see how important, or rather how necessary for the fertilisation of the plant, is the curious slipper-like shape of the labellum, in leading insects to insert their probosces by the lateral passages close to the anthers. The upper, shield-like, rudimentary anther is equally, and in the same manner, necessary.
The economy shown by nature in her resources is striking: in all Orchids seen by me, excepting Cypripedium, the stigma is more or less concave and is viscid, so as to secure the dry pollen, brought to it by means of the viscid matter secreted by the modified stigma, called the rostellum. In Cypripedium alone the pollen is glutinous, and takes on itself the function of viscidity, which in other Orchids belongs to both the true stigma and
the modified stigma or rostellum. The stigma itself, on the other hand, in Cypripedium entirely loses its viscidity, and at the same time becomes slightly convex, so as more effectually to rub off the glutinous pollen adhering to an insect's proboscis. Thus the act of fertilisation is completed, and there is no superfluity in the means employed.
SECRETION OF NECTAR.
Many exotic Orchids secrete plenty of nectar in our hot-houses. I have found the horn-like nectaries of Aerides filled with fluid; and Mr. Rodgers, of Sevenoaks, informs me that he has taken crystals of sugar of considerable size from the nectary of A. cornutum. In nearly all the flowers of Angræcum distichum sent me from Kew, insects had bitten holes through the nectaries, so as to get more readily at the nectar: if insects were invariably to follow this bad habit in the plant's native African home, undoubtedly it would soon become extinct, for it would never produce a seed. The nectar-secreting organs present great diver-
sity in structure and position in the various genera; but apparently always form part of the base of the labellum. In Dendrobium chrysanthum the nectary consists of a shallow saucer; in Evelyna of two large united cellular balls; in Bolbophyllum cupreum of a medial furrow. In Cattleya the nectary penetrates the ovarium; in Angræcum sesquipedale it attains, as we have seen, the astonishing length of above eleven inches; but I will not detail the several cases. The nectar-secreting apparatus of Coryanthes, however, is so remarkable, as described by M. Ménière,* that it must not be quite passed over: two little horns near the strap-like junction of the labellum with the base of the column, secrete so much limpid nectar, with a slightly sweet taste, that it slowly drops down. M. Ménière estimates the total quantity secreted by a single flower at about an English ounce. But the remarkable point is that the deeply-hollowed end of the labellum hangs some way down, exactly beneath the two little horns, and catches the drops as
* 'Bulletin de la Soc. Bot. de France,' tom ii. 1855, p. 351.
they fall, just like a bucket suspended some way beneath a dripping spring of water.
Although the secretion of nectar is of the highest importance to Orchids by attracting insects, which are indispensable to their fertilisation, yet it would seem that the secretion acts also, at least in some cases, as an excretion. One may suspect this to be the case with the Coryanthes from the immense quantity secreted; but the bracteæ of some Orchids have been observed* to secrete nectar; and as these lie outside the flower, the secretion cannot serve any useful purpose by attracting insects. Mr. Rodgers informs me that he has observed much nectar secreted at the base of the footstalks of the flowers in Vanilla. It is in perfect accordance with the scheme of nature, as worked out by natural selection, that matter excreted to free the system from superfluous or injuring substances
* J. G. Kurr, 'Ueber die Bedeutung der Nektarien,' 1833, s. 28, on the authority of Treviranus and Curt. Sprengel. The calyx of certain species of Iris also (idem, s. 25) secretes nectar. I have observed much nectar secreted by the stipules of Vicia sativa and faba, which is eagerly collected by bees. Glands on the under side of the leaves of the common laurel also secrete nectar, which, though the drops are extremely minute, is sought for by various insects.
should be utilised for purposes of the highest importance. To give an example in strong contrast with flowers and honey, the larvæ of certain beetles (Cassidæ, &c.) use their own excrement to make an umbrella-like projection for their tender bodies.
In Cypripedium the slipper-like labellum seems well adapted to hold and collect nectar; but I have found no such collection in the four previously named species; nor does C. calceolus ever secrete nectar according to Kurr.* The labellum, however, in these four species is studded with hairs; and I have almost always noticed on their tips little drops of slightly viscid fluid, which if sweet would certainly suffice to attract insects; this viscid fluid when dried forms a brittle crust, but I could not perceive in it any traces of crystallisation.
It may be remembered that in the first chapter evidence was given proving that nectar is never found within the spur-like nectary of several species of Orchis, but that between its two membranes there is an abundant supply of fluid. In all the species
* 'Bedeutung der Nektarien,' 1833, s. 29.
thus characterised the viscid matter of the disc of the pollinium sets hard in a minute or two, and it would be an advantage to the plant if an insect were delayed in getting the nectar by having to puncture the nectary at several points, so that time should be allowed for the setting of the viscid matter. On the other hand, in all the Ophreæ which have nectar ready stored within the nectary the viscid matter does not rapidly set hard, and there would be no advantage in insects being delayed.
In the case of cultivated exotic Orchids which have a nectary, without nectar, it is of course impossible to feel sure that it would be empty under more natural conditions. Nor have I made many comparative observations on the rate of setting of the viscid matter of the disc in exotic forms. Nevertheless it seems that certain Vandeæ are in the same predicament as our British species of Orchis; thus Calanthe masuca has a very long nectary, which in all the specimens examined was internally quite dry, and was inhabited by powdery Cocci; but in the intercellular spaces between its two coats there was much fluid;
in this species the viscid matter of the disc, after its surface had been disturbed, entirely lost its adhesiveness in two minutes. In an Oncidium the disc, similarly disturbed, became dry in one minute and a half; in an Odontoglossum in two minutes; neither of these Orchids have free nectar. On the other hand, in Angræcum sesquipedale, which has free nectar stored within the lower end of its nectary, the disc of the pollinium, removed from the plant, and with its surface disturbed, was strongly adhesive after forty-eight hours.
Sarcanthus teretifolius offers a more curious case. The disc quite lost its viscidity, after the removal of the pollinium from the rostellum, in less than three minutes. Hence it might have been expected that fluid would have occurred in the intercellular spaces of the nectary, but none within the nectary; nevertheless there was fluid in both places, so that we here see the two conditions of the nectar-secreting organs combined in the same flower. It might perhaps have been thought that insects would have rapidly sucked the free nectar and neglected that between the two coats. But I strongly suspect that insects are
delayed in sucking the free nectar, so as to allow the viscid matter to set hard, by totally different means. In this Orchid the labellum with its nectary is an extraordinary organ. I wished to have had a drawing of its structure; but found that it was as hopeless as to give one of the wards of a complicated lock: even the skilful Bauer, with numerous figures and sections on a large scale, hardly makes the structure intelligible. So complicated is the passage that I failed in repeated attempts to pass a bristle from the outside of the flower into the nectar-receptacle; or in a reversed direction from the cut-off end of the nectar-receptacle to the outside. No doubt an insect with a voluntarily flexible proboscis could wind it through the passages, and thus get the nectar; but in effecting this, some delay would be caused; and time would be allowed for the curious square disc to become securely semented to the insect's head or body.
As in Epipactis the cup at the base of the labellum serves as a nectar-receptacle, I expected to find that the analogous cup in Stanhopea, Acropera, &c., would serve for the same purpose; but I could never find a drop
of nectar in it. According, also, to M. Ménière,* this is never the case in these genera, nor in Gongora, Cirrhæa, and others. In Catasetum tridentatum, and in its female the Monachanthus , we see that the upturned cup cannot possibly serve as a nectar-receptacle. What then attracts insects to these flowers? That they must be attracted is certain; more especially in the case of Catasetum, in which the sexes stand on separate plants. In many genera of Vandeæ there is no trace of any nectar-secreting organ or receptacle; but in all these cases, as far as I have seen, the labellum is either thick and fleshy or is furnished with excrescences. The labellum, for instance, of Oncidium and of Odontoglossum offers all sorts of singular protuberances. In Calanthe we have (Fig. XXIV.) a cluster of odd little spherical warts on the labellum, together with an extremely long nectary, which does not include nectar; in Eulophia viridis the short nectary is in the same condition, and the labellum is covered with longitudinal and fimbriated ridges. In some also of the Ophreæ, which have no nectary, the labellum, as in the Fly Ophrys, has two
* 'Bulletin Bot. Soc. de France,' tom ii. 1855, p. 352.
shining protuberances at its base, placed beneath the two pouches. Lindley has remarked that the use of these strange and diversified excrescences is quite unknown.
From the position, relative to the viscid disc of the pollinium, which these excrescences hold, and from the absence of nectar, it seems to me highly probable that they afford food, and thus attract either Hymenoptera or flower-feeding Coleoptera. I mention this belief because a close examination of the flowers of the Vandeæ, which in their native country have had their pollinia removed, would soon settle this point. There is no more inherent improbablity in a flower being habitually fertilised by an insect coming to feed on the labellum, than in seeds being habitually disseminated by birds attracted by the sweet pulp in which they are embedded. But I am bound to state that Dr. Percy had the thick and furrowed labellum of a Warrea analysed for me, by fermentation over mercury, and it gave no evidence of containing more saccharine matter than the other petals. On the other hand, the thick labellum of Catasetum, and even the bases of the upper petals in Mormodes
ignea, had, as previously stated, a slightly sweet, rather pleasant, and nutritious taste.
We have now done with exotic Orchids. To me the study has been most interesting of these wonderful and often beautiful productions, so unlike common flowers, with all their many adaptations, with parts capable of movement, and other parts endowed with something so like, though no doubt really different from, sensibility. The flowers of Orchids, in their strange and endless diversity of shape, may be compared with the great vertebrate class of Fish, or still more appropriately with tropical Homopterous insects, which seem to us in our ignorance as if modelled by the wildest caprice.
CHAPTER VII.
Homologies of Orchid-flowers—The great modification which they have undergone—Gradation of organs, of the rostellum, of the pollen-masses—Formation of the caudicle—Genealogical affinities—Mechanism of the movement of the pollinia—Uses of the petals—Production of seed—Importance of trifling details of structure—Cause of the vast diversity of structure for the same general purpose—Cause of the perfection of the contrivances in Orchids—Summary on insect-agency—Nature abhors perpetual self-fertilisation.
THE theoretical structure of few flowers has been so largely discussed as that of Orchids; nor is this surprising, seeing how unlike they are to common flowers. No group of organic beings can be well understood until their homologies are made out; that is, until the general pattern, or, as it is often called, the ideal type, of the several members of the group is intelligible. No one member may now exist exhibiting the full pattern; but this does not make the subject less important to the naturalist,—probably makes it more important for the full understanding of the group.
The homologies of any being, or group of beings, can be most surely made out by tracing their embryological development when that is possible; or by discovery of organs in a rudimentary condition; or by tracing, through a long series of beings, a close gradation from one part to another, until the two parts, or organs, employed for widely different functions, and most unlike each other, can be joined by a succession of short links. No instance is known of a close gradation between two organs, unless they be homologically one and the same organ.
The importance of the science of Homology rests in its giving us the key-note of the possible amount of difference in plan within any group; it allows us to class under proper heads the most diversified organs; it shows us gradations which would otherwise have been overlooked, and thus aids us in our classification; it explains many monstrosities; it leads to the detection of obscure and hidden parts, or mere vestiges of parts, and shows us the meaning of rudiments. Besides these practical uses, to the naturalist who believes in the gradual modification of organic
beings, the science of Homology clears away the mist from such terms as the scheme of nature, ideal types, archetypal patterns or ideas, &c.; for these terms come to express real facts. The naturalist, thus guided, sees that all homological parts or organs, however much diversified, are modifications of one and the same ancestral organ; in tracing existing gradations he gains a clue in tracing, as far as that is possible, the probable course of modification during a long line of generations. He may feel assured that, whether he follows embryological development, or searches for the merest rudiments, or traces gradations between the most different beings, he is pursuing the same object by different routes, and is tending towards the knowledge of the actual progenitor of the group, as it once grew and lived. Thus the subject of Homology gains largely in interest.
Although this subject, under whatever aspect it be viewed, will always be most interesting to the student of nature, it is very doubtful whether the following details on the homological nature of the flowers of Orchids will be endured by the general reader. If,
indeed, he should care to see how much light, through far from perfect, homology throws on a subject, this will, perhaps, be nearly as good an instance as could be given. He will see how curiously a flower may be moulded out of many separate organs,—how perfect the cohesion of primordially distinct parts may become,—how organs may be used for purposes widely different from their proper function,—how other organs may be entirely suppressed, or leave mere useless emblems of their former existence. Finally, he will see how enormous has been the total amount of change from the simple parental or typical structure which these flowers have undergone.
Robert Brown first clearly discussed the homologies of Orchids,* and left, as might be expected, little to be done. Guided by the general structure of monocotyledonous plants, and by various considerations, he propounded the doctrine that the flower properly consists of three sepals, three petals, six anthers in two whorls or circles (of which only one anther belonging to the outer whorl is perfect in all
* I believe his latest views are given in his celebrated paper, read Nov. 1-15, 1831, in the 'Linnæan Transactions,' vol. xvi. p. 685.
common forms), and of three pistils, with one of them modified into the rostellum. These fifteen organs are arranged as usual, alternately, three within three, in five whorls. Of the existence of three of the anthers in two whorls, R. Brown offers no sufficient evidence, but believes that they are combined with the labellum, whenever that organ presents crests or ridges. In these views Brown is followed by the greatest living authority on Orchids, namely, Lindley.
Brown traced the spiral vessels in the flower by making transverse sections,* and only occasionally, as far as it appears, by longitudinal sections. As spiral vessels are developed at a very early period of growth, which always gives much value to an organ in making out homologies; and as they are apparently of high functional importance, though their function is not well known, it
* 'Linn. Transac.' Vol. xvi. p. 696-701. Link in his 'Bemerkungen über der Bau der Orchideen' ('Botanische Zeitung,' 1849, s. 745) seems to have also trusted to transverse sections. Had he traced the vessels upwards I cannot believe that he would have disputed Brown's view of the nature of the two anthers in Cypripedium. Brongniart in his admirable paper ('Annales des Sciences Nat.' Tom. xxiv. 1831) incidentally shows the course of some of the spiral vessels.
appeared to me, guided also by the advice of Dr. Hooker, to be worth while to trace upwards all the spiral vessels from the six groups surrounding the ovarium. Of the six ovarian groups of vessels, I will call (though not correctly) that under the labellum the anterior group; that under the upper sepal the posterior group; and the two groups on both sides of the ovarium the antero-lateral and postero-lateral groups.
The result of my dissections is given in the following diagram. The fifteen little circles represent so many groups of spiral vessels, in every case traced down to one of the six large ovarian groups. They alternate in five whorls, as represented; but I have not attempted to give the actual distances at which they stand apart. In order to guide the eye, the three central groups running to the three pistils are connected by a triangle.
Five groups of vessels run into the three sepals and two upper petals; three enter the labellum; and seven run up the great central column. These vessels are arranged, as may be seen, in rays proceeding from the axis of the flower; and all on the same ray invari-
ably run into the same ovarian group: thus the vessels supplying the upper sepal, the
Fig. XXXII.
SECTION OF THE FLOWER OF AN ORCHID.
The little circles show the position of the spiral vessels.
S S. Stigmas; S r, stigma modified into the rostellum.
A 1. Fertile anther of the outer whorl; A 2 A 3, anthers of the same whorl combined with the lower petal, forming the labellum.
a 1 a 2. Rudimentary anthers of the inner whorl (fertile in Cypripedium), generally forming the clinandrum; a 3, third anther of the same whorl, when present, forming the front of the column.
fertile anther (A 1), and the upper pistil or stigma (i.e. rostellum S r), all unite and form the posterior ovarian group. Again, the
vessels supplying one of the lower sepals, the corner of the labellum, and one of the two stigmas (S), unite and form the antero-lateral group; and so with all the other vessels.
Hence, if the existence of groups of spiral vessels can be trusted, and Dr. Hooker informs me that he has never known them to speak falsely, the flower of an Orchid certainly consists of fifteen organs, in a much modified and confluent condition. We see three stigmas, with the two lower ones generally confluent, and with the upper one modified into the rostellum. We see six stamens, arranged in two whorls, with one alone (A 1) generally fertile. In Cypripedium, however, two stamens of the inner whorl (a 1 and a 2) are fertile, and in other Orchids these two are represented in various ways more plainly than the remaining stamens. The third stamen of the inner whorl (a 3), when its vessels can be traced, forms the front of the column: Brown thought that it often formed a medial excrescence, or ridge, cohering to the labellum; or, in the case of Glossodia,* a filamentous
* See Brown's observations under Apostasia in Wallich's 'Plantæ Asiaticæ rariores,' 1830, p. 74.
organ, freely projecting in front of the labellum. The former conclusion does not agree with my dissections; about Glossodia I know nothing. The two infertile stamens of the outer whorl (A 2, A 3) were believed by Brown to be sometimes represented by lateral excrescences on the labellum; I find these vessels invariably present in the labellum of every Orchid examined,—even when the labellum is very narrow, or quite simple, as in Malaxis, Herminium and Habenaria.
We thus see that an Orchid-flower consists of five simple parts, namely, three sepals and two petals; and of two compounded parts, namely, the column and the labellum. The column is formed of three pistils, and generally of four stamens, all completely confluent. The labellum is formed of one petal and two petaloid stamens of the outer whorl, likewise completely confluent. I may remark, as making this fact more probable, that in the allied Marantaceæ the stamens, even the fertile stamens, are often petaloid, and partially cohere. This view of the nature of the labellum explains its large size, its frequently tripartite form, and especially its manner of
coherence to the column, unlike that of the other petals.* As rudimentary organs vary much, we can thus also probably understand the variability, which Dr. Hooker informs me is characteristic of the excrescences on the labellum. In some Orchids which have a spur-like nectary, the two sides are apparently formed by the two modified stamens; thus in Gymnadenia conopsea (but not in Orchis pyramidalis), the vessels, proceeding from the antero-lateral ovarian group, run down the sides of the nectary; those from the anterior group run down the exact middle of the nectary, then returning up the opposite side form the mid-rib of the labellum. The extension of these lateral elements of the nectary apparently explains the tendency, as in Calanthe, Orchis morio, &c, to the bifurcation of its extremity.
The number, position, and course of the spiral vessels exhibited in the diagram (Fig. XXXII.), were observed in some Vandeæ and Epidendreæ. † In the Malaxeæ all were
* Link remarks on the manner of coherence of the labellum to the column in his 'Bemerkungen' in 'Bot. Zeitung,' 1849, s. 745.
† It may be advisable to give a few details on the flowers which I dissected; but I looked to special points, such as the course
observed excepting a 3, which is the most difficult of all to trace, and which apparently
of the vessels in the labellum, in many cases not worth here giving. In the Vandeæ I traced all the vessels in Catasetum tridentatum and saccatum; the great group of vessels going to the rostellum separate (as likewise in Mormodes) from the posterior ovarian group, beneath the bifurcation supplying the upper sepal and fertile anther; the anterior ovarian group runs a little way along the labellum before bifurcating, and sending a group (a3) up the front of the column; the vessels proceeding from the postero-lateral group run up the back of the column, on each side of those running to the fertile anther, and do not go to the edges of the clinandrum. In Acropera luteola the base of the column, where the labellum is attached, is much produced, and the vessels of the whole anterior ovarian group are similarly produced; those (a3) going up the front of the column are abruptly reflected back; the vessels at the point of reflexion are curiously hardened, flattened, and produced into odd crests and points. In an Oncidium I traced the vessels Sr to the viscid gland of the pollinium. In the Epidendreæ I traced all the vessels in a Cattleya; and all in Evelyna carivata except a3, which I did not search for. In the Malaxeæ I traced all in Liparis pendula except a3, which I do not believe is present. In Malaxis paludosa I traced nearly all the vessels. In Cypripedium barbatum and purpuratum I traced all except a3, which I am nearly sure does not exist. In the Neotteæ I traced in Cephalanthera grandiflora all the vessels, excepting that to the aborted rostellum and those to the two auricles a1 and a2, which were certainly absent. In Epipactis I traced all excepting a1, a2, and a3, which are certainly absent. In Spiranthes autumnalis the vessel Sr runs to the bottom of the fork of the rostellum: there are no vessels to the membranes of the clinandrum in this Orchid nor in Goodyera. In none of the Ophreæ do the vessels a1, a2, and a3 occur. In Orchis pyramidalis I traced all the others, including two to the two separate stigmas: in this species the contrast between the vessels of the labellum and of the other sepals and petals is striking, as in the latter the vessels do not
is oftenest absent. In the Cypripedeæ, again, all were traced except a3,* which, I feel pretty sure, was here really absent: in this tribe the stamen (A1) is represented by a conspicuous shield-like rudiment, and a1 and a2 are developed into two fertile anthers. In the Ophreæ and Neotteæ all were traced, with
branch, whilst the labellum has three vessels, the lateral ones running of course into the antero-lateral ovarian group. In Gymnadenia conopsea I traced all the vessels; but I am not sure whether the vessels supplying the sides of the upper sepal do not, as in the allied Habenaria, wander from their proper course and enter the postero-lateral ovarian group: the vessel Sr, going to the rostellum, enters the little folded crest of membrane, which projects between the bases of the anther-cells. Lastly, in Habenaria chlorantha I traced all the vessels, excepting of course the three of the inner staminal whorl, and I looked carefully for a3: the vessel supplying the fertile anther runs up the connective membrane between the two anther-cells, but does not bifurcate: the vessel to the rostellum runs up to the top of the shoulder or ledge beneath the connective membrane of the anther, but does not bifurcate and extend to the two widely-separated viscid discs.
* From Irmisch's ('Beiträge zur Biologie der Orchideen,' 1853, s. 78 and 42) description of the development of the flower-bud of Cypripedium, it would appear that there is a tendency to the formation of a free filament in front of the labellum, as in the case of Glossodia before mentioned; and this will perhaps account for the absence of spiral vessels, proceeding from the anterior ovarian group, and coalescing with the column. In Uropedium, a genus which A. Brongniart ('Annal. des. Sc. Nat.,' 3rd series, Bot. tom. xiii. P. 114) considers closely allied to, and even perhaps a monstrosity of, Cypripedium, a third fertile anther occupies this same exact position.
the important exception of the vessels belonging to the three stamens (a1, a2, and a3) of the inner whorl. In Cephalanthera grandiflora, however, I clearly saw a 3 proceeding from the anterior ovarian group, and running up the front of the column: this anomalous member of the Neotteæ has no rostellum, and the vessel marked Sr in the diagram was entirely absent, though seen in every other Orchid.
Although in no true Orchid, excepting Cypripedium, the two anthers (a1 and a2) of the inner whorl are fully developed, their rudiments are generally present and are often utilised; for they generally form the membranous sides of the clinandrum, or cup on the summit of the column, which includes and protects the pollen-masses. These rudiments thus aid their fertile brother-anther. In the young flower-bud of Malaxis paludosa, the close resemblance between the membranes of the clinandrum and the fertile anther, in shape, texture, and in the height to which the spiral vessels extend, is most striking: it is impossible to doubt that in these two membranes we see two rudimen-
tary anthers. In Evelyna, one of the Epidendreæ, the clinandrum was similarly formed, as were the horns of the clinandrum in Masdevallia, which likewise serve to keep the labellum at the proper distance from the column. In Liparis pendula and some other forms, these two rudimentary anthers formed not only the clinandrum, but likewise wings, projecting on each side of the entrance to the stigmatic cavity, and serving as guides for the insertion of the pollen-masses. In Acropera and Stanhopea, as far as I could make out, the membranous borders of the column, down to its base, were also thus formed; but in other cases, as in Cattleya, the wing-like borders of the column seemed to be simple developments of the two pistils. In this latter genus, as well as in Catasetum, these same two rudimentary stamens, judging from the position of the vessels, served chiefly to strengthen the back of the column; and the strengthening of the front of the column is the sole function of the third stamen (a3) of the inner whorl, in those cases in which I observed it. This third stamen runs up the middle of the column to the lower edge, or lip, of the stigmatic cavity.
I have said that in the Ophreæ and Neotteæ the spiral vessels, marked a1, a2, a3 in the diagram, are entirely absent, and I looked carefully for them; but in nearly all the members of these two tribes, two small papillæ, or auricles as they have been often called, stand in exact the position which the two first of these three anthers would have occupied, had they been developed. Not only do they stand in this position, but the column in some cases, as in Cephalanthera, has on each side a prominent ridge, running from them to the bases or mid-ribs of the two upper petals; that
is, in the proper position of the filaments of these two stamens. It is, again, impossible to doubt that the membranes of the clinandrum in Malaxis are formed by these two anthers in a rudimentary and modified condition. Now, from the perfect clinandrum of Malaxis, through that of Spiranthes, Goodyera, Epipactis latifolia, and E. palustris (see Fig. XIV. and XIII.), to the minute and slightly flattened auricles in the genus Orchis, a perfect gradation can be traced. Hence I conclude that these auricles are doubly rudimentary; that is, that they are rudiments of the membranous sides of the clinandrum, these membranes themselves being rudiments of the two anthers so often referred to. The absence of spiral vessels running to the auricles by no means seems sufficient to overthrow these several arguments on their much-disputed nature; that such vessels may quite disappear, we have proof in Cephalanthera grandiflora, in which the rostellum and its vessels are completely aborted.
Finally, then, with respect to the six stamens or anthers which ought to be represented in every Orchid: the three belonging to the outer whorl are always present, with the upper one generally fertile, and the two lower ones invariably petaloid and forming part of the labellum; the three stamens of the inner whorl are less plainly developed, especially the lower one, a3, which, when it can be detected, serves only to strengthen the column, and, in some rare cases, according to Brown, forms a separate projection, or filament; the upper two anthers of this inner whorl are fertile in Cypripedium, and in other cases are generally represented either by mem-
branous expansions, or by minute auricles without spiral vessels. These auricles, however, are sometimes quite absent, as in some species of Ophrys.
On this view of the homologies of Orchid-flowers, we can understand the existence of the conspicuous central column,—the large size, generally tripartite form, and peculiar manner of attachment of the labellum,—the origin of the clinandrum,—the relative position of the single fertile anther in most Orchids, and of the two fertile anthers in Cypripedium,—the position of the rostellum, as well as of all the other organs,—and, lastly, the frequent occurrence of a bilobed stigma, and the occasional occurrence of two distinct stigmas.
I have encountered only one case of difficulty on the foregoing views, namely in Habenaria, and in the allied Bonatea. These flowers have undergone such an extraordinary amount of distortion, owing to the wide separation of their anther-cells and of the two viscid discs of the rostellum, that any anomaly in them is the less surprising. The anomaly relates only to the vessels supplying the sides
of the upper sepal and of the two upper petals; the vessels running into their mid-ribs and into all the other more important organs persue the same identical course as in all the other Ophreæ. The vessels on the sides of the upper sepal, instead of uniting with the mid-rib, and entering the posterior ovarian group, diverge and enter the postero-lateral groups: again, the vessels on the anterior side of the upper petals, instead of uniting with the mid-rib and entering the postero-lateral ovarian groups, diverge, or wander from their proper course, and enter the antero-lateral groups.
This anomaly is so far of importance, as it throws some doubt on the view which I have taken of the labellum being always an organ compounded of one petal and two petaloid stamens; for if any one were to assume that from some unknown cause the lateral vessels of the lower petal in an early progenitor of the Orchidean order had wandered from their proper course into the antero-lateral ovarian groups, and that this structure had been inherited by all existing Orchids, even by those with the smallest and simplest labellums, I could answer only as follows; but the answer
is, I think, satisfactory. From the analogy of other monocotyledonous plants, we might expect the hidden presence of fifteen organs in the flowers of Orchids, arranged alternately in five whorls; and in Orchid-flowers we do find fifteen groups of vessels exactly thus arranged. Hence there is a strong probability that the vessels, A 2 and A 3, which enter the sides of the labellum, not in one or two cases, but in all Orchids seen by me, and which occupy the precise position which they would have occupied had they supplied two normal stamens, do really represent modified and petaloid stamens, and are not lateral vessels of the lower petal which have wandered from their proper course. In Habenaria and Bonatea,* on the other hand,
* In Bonatea speciosa, of which I have examined only dry specimens sent me by Dr. Hooker, the vessels from the sides of the upper sepal enter the postero-lateral ovarian group, exactly as in Habenaria. The two upper petals are divided down to their bases, and the vessels supplying the anterior segment and those supplying the anterior portion of the posterior segment unite and then run, as in Habenaria, into the antero-lateral (and therefore wrong) groups. The anterior segments of the two upper petals cohere with the labellum, making it, in a most unusual manner, five-segmented. The two wonderfully protuberant stigmas also cohere to the upper surface of the labellum; and the lower sepals apparently also cohere to its under side.
the vessels from the sides of the upper sepal and of the two upper petals, which enter the wrong ovarian groups, cannot possibly represent any now lost and once distinct organs.
We have now finished with the general homologies of the flowers of Orchids. It is interesting to look at one of the magnificent exotic species, or, indeed, at one of our humblest forms, and observe how profoundly it has been modified, as compared with all ordinary flowers,—with its usually great labellum, formed of one petal and two petaloid stamens,—with its singular pollen-masses, presently to be referred to,—with its column formed of seven cohering organs, of which three alone perform their proper function, namely, one anther and two generally confluent stigmas,—with the third stigma incapable of fertilisation and modified into the wonderful rostellum,—with three of the anthers no longer capable of producing pollen, but serv-
Consequently a section of the base of the labellum divides one lower petal, two petaloid anthers, portions of the two upper petals, and apparently of the two lower sepals and the two stigmas: altogether the section passes through the whole of or through portions of no less than seven or nine organs. The base of the labellum is here as complex an organ as the column of other Orchids.
ing either to protect the pollen of the fertile anther, or to strengthen the column, or existing as mere rudiments, or entirely suppressed. What an amount of modification, change of function, cohesion, and abortion do we here see! Yet hidden in that column, with its surrounding petals and sepals, we know that there are fifteen groups of vessels, arranged three within three, in alternating order, which probably have been preserved to the present time from having been developed in each flower at a very early period of growth, before the shape or existance of this or that part signified to the well-being of the plant.
Can we, in truth, feel satisfied by saying that each Orchid was created, exactly as we now see it, on a certain "ideal type;" that the Omnipotent Creator, having fixed on one plan for the whole Order, did not please to depart from this plan; that He, therefore, made the same organ to perform diverse functions—often of trifling importance compared with their proper function—converted other organs into mere purposeless rudiments, and arranged all as if they had to stand separate, and then made them cohere? Is it not a more
simple and intelligible view that all Orchids owe what they have in common to descent from some monocotyledonous plant, which, like so many other plants of the same division, possessed fifteen organs, arranged alternately three within three in five whorls; and that the now wonderfully changed structure of the flower is due to a long course of slow modification,— each modification having been preserved which was useful to each plant, during the incessant changes to which the organic and the inorganic world has been exposed?
On the gradation of Organs.—The rostellum, the pollinia, the labellum, and, in a lesser degree, the column, are the most remarkable points in the structure of Orchids. Of the two latter parts enough has been already said. No organ like the rostellum exists in any other flower. If the homologies in Orchids had not been pretty well known, those who believe in the separate creation of each being might have advanced this as a case of a perfectly new organ specially created, and which could not have been developed by successive slow modifications of any pre-existing
part. But, as Robert Brown long ago remarked, it is not a new organ. It is impossible to look at the two groups of spiral vessels (Fig. XXXII.) running from the mid-ribs of the two lower sepals to the two, sometimes quite distinct, lower stigmas, and then look at the third group of vessels running from the mid-rib of the upper sepal to the rostellum, which occupies exactly the position of the third stigma, and doubt its homological nature. There is every reason to believe that the whole of this upper stigma, and not merely a part, has been converted into the rostellum; for there are plenty of cases of two stigmas, but not one instance of three stigmatic surfaces being present in those Orchids which have a rostellum. On the other hand, in Cypripedium and Apostasia (the latter ranked by Brown in the Orchidean order), which are destitute of a rostellum, the stigmatic surface is trifid.
As we know only those plants which are now living, it is impossible to follow all the gradations by which the upper stigma has been converted into the rostellum; but let us see what are the facilities and indications of
such a change having been effected. The change of function has not been so great as it at first appears. The function of the rostellum is to secrete a quantity of viscid matter; it has lost that of fertility, but this loss is so common with plants that it is hardly worth notice. The stigmas of Orchids, as well as of most other plants, secrete viscid matter, the use of which in all cases is to retain the pollen when brought by any means to its surface. Now, if we look to one of the simplest rostellums,—for instance, to that of Cattleya or Epidendrum,—we find a thick layer of viscid matter, not distinctly separated from the viscid surface of the two confluent stigmas: its action is simply to smear and affix to the back of a retreating insect the pollen-masses, which are thus dragged out of the anther and transported to another flower, where they are retained by the almost equally viscid stigmatic surface. So that the office of the rostellum is still to secure the pollen-masses, but indirectly by means of their attachment to an insect's body.
The viscid matter of the rostellum and of the stigma appears to have nearly the same
character: that of the rostellum generally has the peculiar property of quickly drying or setting hard; that of the stigma, when removed from the plant, apparently dried more quickly than gum-water of about equal tenacity. This tendency to dry is the more remarkable, as Gärtner* found that drops of the stigmatic secretion from Nicotiana did not dry in two months. The viscid matter of the rostellum in many Orchids when exposed to the air changes colour with remarkable quickness, and becomes brownish-purple; and I have noticed a similar but slow change of colour in the viscid secretion of the stigmas of some Orchids, as of Cephalanthera grandiflora. When the viscid disc of an Orchis, as Bauer and Brown have also observed, is placed in water, minute particles are expelled with violence in a peculiar manner; and I have observed exactly the same fact in the layer of viscid matter covering the stigmatic utriculi in an unopened flower of Mormodes ignea.
In order to compare the minute structure of the rostellum and stigma, I examined young flower-buds of Epidendrum cochleatum and
* 'Beiträge zur Kenntnis der Befruchtung,' 1844, s. 236.
floribundum, which, when mature, have a simple rostellum. The posterior surface was the same in both organs: the rostellum at this early age consisted of a mass of nearly orbicular cells, containing spheres of brown matter, which resolve themselves into the viscid matter: the stigma was covered with a thinner layer of similar cells, and beneath them were the coherent spindle-formed utriculi. The utriculi are believed to be connected with the penetration of the pollen-tubes; and their absence in the rostellum probably accounts for its infertility. As I do not find the exterior layer of nearly orbicular cells on the stigma in the bud-state (which apparently secrete the viscid matter), mentioned by more experienced observers, I cannot help feeling some doubt on the subject; though I have no other reason to doubt my accuracy. If the structure of the rostellum, in one of the simplest Orchids, and of the stigma, be as I have described them, their only difference is, that in the rostellum the layer of cells, which secretes the viscid matter, is thicker, and the utriculi have disappeared.
Hence, during a course of slow modific-
ation, it is quite conceivable that the upper stigma, whilst still in some degree fertile or capable of penetration by the pollen-tubes, might secrete a superfluity of viscid matter; and that insects smeared with this might remove and transport the pollen-masses to the stigmas of other flowers. In this case an incipient rostellum would have been formed.
The following details on the rostellum and pollinia will have no interest to any one, unless he cares much about the structure of Orchids, or wishes to see how far very different states of the same organ can be graduated together within the limits of the same order. In the several Tribes, the rostellum presents a marvellous amount of diversity of structure; but most of the differences can be connected without very wide breaks. One of the most striking diferences is, that either the whole anterior surface to some depth or only the central portion becomes viscid, in the latter case the surface retaining, as in Orchis, a membranous condition. But these two states so graduate into each other, that it is scarcely possible to draw any line of separation: thus, in Epipactis, the exterior surface undergoes a
vast change from its early cellular condition, and becomes converted into a highly elastic and tender membrane, which is in itself slightly viscid, and allows the underlying viscid matter readily to exude; yet it acts as a membrane, with its under surface lined with much more viscid matter: in Habenaria chlorantha the exterior surface is highly viscid, but still closely resembles, under the microscope, the exterior membrane of Epipactis. Lastly, in some species of Oncidium, &c., the viscid exterior surface differs, as far as appearance under the microscope goes, from the underlying viscid layer only in colour; but it must have some essential difference; for I find that, until this very thin exterior layer is disturbed, the underlying matter remains viscid; whilst, after it has been disturbed, it rapidly sets hard. The gradation in the state of the surface of the rostellum is not surprising, for in all cases in the bud the surface is cellular; so that an early condition has only to be more or less perfectly retained.
The nature of the viscid matter differs remarkably in different Orchids: in Listera
it sets hard almost instantaneously, more quickly than plaster of Paris; in Malaxis and Angræcum it remains fluid and viscid for several days; but these two states pass into each other by many gradations: in an Oncidium I have observed the viscid matter to dry in a minute and a half; in some species of Orchis in two or three minutes; in Epipactis in ten minutes; in Gymnadenia in two hours; and in Habenaria in over twenty-four hours. After the viscid matter of Listera has set hard, neither water nor weak spirits of wine has any effect on it; whereas that of Habenaria bifolia, after being kept in spirits, and after having been dried for several months, when moistened was as sticky as ever; the viscid matter in some species of Orchis, when remoistened, presented an intermediate condition.
One of the most important differences in the state of the rostellum is, whether or not the pollinia are congenitally attached to it. I do not allude to those cases in which the upper surface of the rostellum becomes viscid, as in Malaxis and some Epidendrums, and adheres without mechanical aid to the pollen-masses;
for these cases present no difficulty, and can be graduated together. But I refer to the so-called congenital attachment of the pollinia by their caudicles. It is not, however, strictly correct to speak of congenital attachment, for the pollinia at an early period are invariably free, and become attached either earlier or later in different Orchids. No actual gradation is at present known in the process of attachment; but it can be shown to depend on very simple conditions and modifications. In the Epidendreæ the pollinia consist of a ball of waxy pollen, with a long caudicle (formed of elastic threads with adherent pollen-grains), which never becomes spontaneously attached to the rostellum. Cymbidium giganteum, on the other hand, has a congenitally attached caudicle, but its structure is identically the same, with the sole difference, that the elastic threads near its base adhere to, instead of simply lying on, the upper lip of the rostellum.
In an allied form, the Oncidium unguiculatum, I examined the development of the caudicles. At an early period the pollen-masses are enclosed im membranous cases;
which soon rupture at one point. At this early period, within the cleft of each pollen-mass, a layer of cellular matter may be detected, with the cells of rather large size, including remarkably opaque matter. The contained matter may be traced undergoing all the stages of development into the translucent substance forming the threads of the caudicles. As this change progresses, the cells themselves disappear. The threads at the one end finally adhere to the waxy pollen, and at the other end, whilst still in a semi-developed state, they protrude through the small opening of the membranous case. and adhere to the rostellum, against which the anther is pressed. So that the adhesion of the caudicle to the back of the rostellum seems to depend solely on the early rupturing of the anther-case, and on the slight protrusion of the caudicles, before they have become fully developed and hardened.
In all Orchids a portion of the rostellum is, in fact, removed by insects when the pollinia are removed; for the viscid matter, though conveniently spoken of as a secretion, is a part of the rostellum in a modified condition.
But in those Orchids, which have their caudicles at an early period attached to the rostellum, a membranous or solid portion of its exterior surface in an unmodified condition is likewise removed. In the Vandeæ this portion is sometimes of considerable size (forming the disc and pedicel of the pollinium), and gives to their pollinia their most remarkable character; but the differences in the shape and size of the removed portions of the rostellum can be finely graduated together, even within the Vandeæ; and still more closely by commencing with the minute oval atom of membrane to which the caudicle of Orchis adheres, passing thense to that of Habenaria bifolia, to that of H. chlorantha with its drum-like pedicel, and thense through many forms to the great disc and pedicel of Catasetum.
In all the cases in which a portion of the exterior surface of the rostellum is removed attached to the caudicles, definite and often complicated lines of separation are formed, or are at least prepared by being weak, in order to allow of the easy separation of the removeable portions. But the formation of the lines
of weakness does not differ much from certain definite portions of the exterior surface of the rostellum assuming a condition intermediate between that of unaltered membrane and of viscid matter, which has been already alluded to. The actual separation of the lines depends in many, perhaps in all, cases on the excitement of a touch; and how this is effected is at present inexplicable. But sensitiveness to touch in the stigma (and the rostellum, as we know, is a modified stigma), and indeed in all the organs of vegetation, is not a very rare attribute of many plants.
In Listera and Neottia, when the rostellum is touched, even by a human hair, two points rupture, and the included viscid matter is instantaneously expelled. Here we have a case towards which as yet no gradation is known. But Dr. Hooker has shown that the structure of the rostellum is at first cellular (as in other Orchids), and that the viscid matter originally developed within these cells is contained, apparently in a state of tension, in the loculi, ready to be expelled as soon as the exterior surface ruptures.
The last and conspicuous difference in the
state of the rostellum which I will mention is the existence, in many Ophreæ, of two widely-separated viscid discs, sometimes included in two separate pouches. Here it at first appears as if there were two rostella; but there is never more than one medial group of spiral vessels. In the Vandeæ we can see how a single viscid disc and single pedicel might become divided into two; for in some Stanhopeas the heart-shaped disc shows a trace of a tendency to division; and in Angræcum we have two distinct discs and two pedicels, either standing close together or removed a little way apart.
It might be thought that a similar gradation from a single rostellum into what appears like two distinct rostella was still more plainly shown in the Ophreæ; for we have the following series,—in Orchis pyramidalis a single disc enclosed in a single pouch; in Aceras two discs touching and affecting each other's shapes, but not actually joined; in Orchis latifolia and maculata two quite distinct discs with the pouch still showing plain traces of division; and, lastly, in Ophrys we have two perfectly distinct pouches, in-
cluding of course two perfectly distinct discs. But this series does not indicate the former steps by which a single rostellum has become divided into two distinct organs; but shows, on the contrary, how the rostellum, after having been anciently divided into two organs, has now in several cases been reunited into a single organ.
This conclusion is founded on the nature of the little medial crest (sometimes called the rostellate process) between the bases of the anther-cells (see Fig. I., B and D). In both divisions of the Ophreæ (those with naked discs and those with discs enclosed in a pouch), whenever the two discs come into close juxtaposition, the medial crest or process appears.* On the other hand, when the two discs stand widely separate, the summit of the rostellum between them is smooth, or nearly smooth. In the Frog Orchis (Peristylus viridis) the overarching summit is bent like the
* Professor Babington ('Manual of British Botany,' 3rd edit.) uses the existence of this "rostellate process" as a character to separate Orchis, Gymnadenia, and Aceras from the other genera of Ophreæ. The group of spiral vessels, properly belonging to the rostellum, runs up, and even into, the base of this crest or process.
roof of a house; and here we see the first stage of the formation of the folded crest. In Herminium, however, which has two separate and large discs, a crest, or solid ridge, is rather more plainly developed than might have been expected. In Gymnadenia conopsea, Orchis maculata, and others, the crest consists of a hood of thin membrane; in O. mascula the two sides of the hood have partly adhered; and in O. pyramidalis and in Aceras it has been converted into a solid ridge. These facts are intelligible only on the view, that, whilst the two discs were gradually, during a long line of generations, brought together, the intermediate portion or summit of the rostellum became more and more arched, until a folded crest, and finally a solid ridge, was formed.
Whether we compare together the state of the rostellum in the various Orchid-tribes, or compare the rostellum with the pistil and stigma of ordinary flowers, the differences are wonderfully great. A simple pistil of a common plant consists of a cylinder surmounted by a small viscid surface. Now, see what a contrast the rostellum of Catasetum,
when dissected from the other elements of the column, presents; and as I traced all the vessels in this Orchid, the drawing may be trusted as approximately accurate. The whole organ has lost its normal function of fertility. Its shape is most singular, with its upper end
Fig. XXXIII.
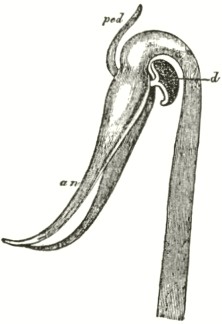
ROSTELLUM OF CATASETUM.
an, antenæ of the rostellum.
d, viscid disc.
ped, pedicel of rostellum, to which the pollen-masses are attached.
thickened, bent over and produced into two long tapering and sensitive antennæ, hollow within like adder's fangs. Behind and between the bases of these antennæ we see the large viscid disc, attached to the pedicel, which
differs in structure from the underlying portion of the rostellum, and is separated from it by a layer of hyaline tissue, that spontaneously dissolves. The disc, attached to the surrounding parts by membrane which ruptures when excited by a touch, consists of strong upper tissue, with an underlying elastic cushion, coated with viscid matter; and this again in most Orchids is overlaid by a film of a different nature. What an amount of specialisation of parts do we here behold! Yet we have seen in the comparatively few Orchids described in this volume, so many and such plainly-marked gradations in the structure of this organ, and such plain facilities for the original conversion of the upper pistil into the rostellum, that it becomes far from incredible, if we had every Orchid which has ever existed throughout the world, that every gap in the existing chain, and every gap in many lost chains, would be amply filled up by a series of easy transitions.
We now come to the last great peculiarity in Orchids, namely, their pollen-masses. The anther opens early, and often deposits the
naked masses of pollen on the back of the rostellum. This action is prefigured in Canna, a member of the family nearest allied to Orchids, in which the pollen is deposited on the pistil, close beneath the stigma. In the state of the pollen there is great diversity: in the anomalous Cypripedium single grains are embedded in a glutinous fluid; in all other Orchids (except the degraded Cephalanthera) each grain consists of generally four united granules.* These compound grains are tied
* In several cases I have observed four tubes emitted from the four granules. In some semi-monstrous flowers of Malaxis paludosa, and of Aceras anthropophora, and in perfect flowers of Neottia nidus-avis, I have observed pollen-tubes emitted from the pollen-grains, whilst still within the anther and not in contact with the stigma. I have thought this worth mentioning as R. Brown (in 'Linn. Transact.' vol. xvi. p. 729) states, apparently with some surprise, that the pollen-tubes were emitted from the pollen, whilst still within the anther, in a decaying flower of Asclepias. These cases show that the protruding tubes are, at least at first, formed at the expense of the contents of the pollen-grains.
Having alluded to the monstrous flowers of Aceras, I will add that I examined several, always the lowest on the spike; in these the labellum was hardly developed, and was pressed close against the stigma. The rostellum was not developed, so that the pollinia had not viscid discs; but the most curious feature was, that the two anther-cells had become, apparently in consequence of the position of the rudimentary labellum, widely separated, and were joined by a connective membrane, almost as broad as that of Habenaria chlorantha!
together by elastic threads, or are cemented together into so-called waxy masses by some unknown substance. The waxy masses thus found are numerous in the Ophreæ; and graduate in the Epidendreæ and Vandeæ from eight, to four, to two, and, by the cohesion of the two, into a single mass. In the Epidendreæ we have both kinds of pollen within the same anther, namely, waxy masses, and caudicles consisting of elastic threads, with numerous compound grains adhering to them.
I can throw no light on the nature of the cohesion of the pollen in these waxy masses; when they are placed in water for three or four days, the compound grains readily fall apart; but the granules forming each grain still firmly cohere, so that the nature of the cohesion in the two cases is different. The elastic threads by which the packets of pollen are tied together in the Ophreæ, and which run far up inside the waxy masses of the Vandeæ, are also of a different nature; for they are acted on by chloroform, and by long immersion in spirits of wine; but these fluids have no particular action on the cohesion of the waxy masses. In several Epidendreæ
and Vandeæ the exterior pollen-grains of these masses differ from the interior grains, in being larger, and in having yellower and much thicker walls. So that in the contents of a single anther-cell we see a surprising degree of differentiation of structure in the pollen, namely, granules cohering by fours, apparently due to their manner of early development, and the compound grains partly tied together by threads and partly cemented together, with the exterior grains different from the interior grains.
In the Vandeæ, the caudicle, composed of fine coherent threads, is developed from the semi-fluid contents of a layer of cellular membrane. As I find that chloroform has a peculiar and energetic action on the caudicles of all Orchids, and likewise on the glutinous matter enveloping the pollen-grains in Cypripedium, and which can easily be drawn out into threads, one may suspect that in this simpler Orchid we see the primordial condition of the elastic threads by which the pollen-grains in so many other more highly-developed Orchids are tied together.*
* Auguste de Saint Hilaire states ('Leçons de Botanique, ' &c.,
The caudicle, when largely developed and destitute of pollen-grains, is the most striking peculiarity of the pollinia. In some Neotteæ, especially in Goodyera, we see it in a nascent condition, projecting just beyond the pollen-mass, with the threads only partially confluent. By tracing in the Vandeæ the gradation from the ordinary naked condition of the caudicle, through Lycaste in which it is almost naked, through Calanthe, to Cymbidium giganteum, in which it is covered with pollen-grains, it seems probably that its ordinary condition has been arrived at by the modification of a
1841, p. 447) that the elastic threads exist in the early bud, after the pollen-grains have been partly formed, as a thick creamy fluid. He adds that his observations on Ophrys apifera have shown him that this fluid is secreted by the rostellum, and is slowly forced drop by drop into the anther. Had not so eminent an authority made this statement I should not have noticed it. It is certainly erroneous. In buds of Epipactis latifolia I opened the anther, whilst perfectly closed and free from the rostellum, and found the pollen-grains united by elastic threads. Cephalanthera grandiflora has no rostellum to secrete the thick fluid, yet the pollen-grains are thus united. In a monstrous specimen of Orchis pyramidalis the auricles, or rudimentary anthers on each side of the proper anther, had become partly developed, and they stood quite on one side of the rostellum and stigma; yet I found in one of these auricles a distinct caudicle (which necessarily had no disc at its extremity), and this caudicle could not possibly have been secreted from the stigma. I could give additional evidence, but it would be superfluous.
pollinium like that of one of the Epidendreæ; namely, by the abortion of the pollen-grains which primordially adhered to separate elastic threads, and by the cohesion of these threads.
In the Ophreæ we have better evidence than that offered by mere gradation, that the long, rigid and naked caudicle has been thus partly formed. I had often observed a cloudy appearance in the middle of the translucent caudicle; and on carefully opening that of Orchis pyramidalis, I found in several specimens in the centre, fully half-way down between the packets of pollen and the viscid disc, several pollen-grains (consisting, as usual, of four united granules), lying quite loose. These grains, from their embedded position, could never by any possibility have been left on the stigma of a flower, and were absolutely useless. Those who can persuade themselves that purposeless organs have been specially created, will think little of this fact. Those, on the contrary, who believe in the slow modification of organic beings, will feel no surprise that the process of change should not always have been perfectly efficient,—that, during and after the many inherited
stages of the abortion of the lower pollen-grains and of the coherence of the elastic threads, there should still exist a tendency to the production of a few pollen-grains where they were formerly developed, and that these should consequently be left entangled within the now coherent threads of the caudicle. They will look at the little clouds formed by the loose pollen-grains within the caudicles of Orchis pyramidalis, as good evidence that the pollen-masses of its parent-form was originally like that of Epipactis or Goodyera, and that the grains slowly disappeared from the lower parts of the mass, leaving the elastic threads naked and ready to cohere into a true caudicle.
As the caudicle, whether longer or shorter in the several species, plays an important part in the fertilisation of the flower, it apparently might have been formed from a nascent condition, as in Epipactis, by the continued preservation of varying increments in its length, each beneficial in relation to other changes in the structure of the flower. But we may conclude from the facts given, that this has not been the sole means,—that the caudicle owes much
of its length to the abortion of the lower pollen-grains. That it has subsequently in some cases been largely increased in length by natural selection, is highly probable; for in Bonatea speciosa the caudicle is actually more than thrice as long as the elongated mass of pollen-grains; and it is improbable that so lengthy a mass of slightly cohering grains should have existed, as an insect could not have safely transported and applied to the stigma a pollen-mass of this shape and size.
We have hitherto considered the gradations in the state of the same organ. To any one with much more knowledge than I possess, it would be an interesting subject to trace, in this great and closely-connected order, the gradations, as far as possible, between the several species and groups of species. To make a perfect gradation, all the extinct forms which have ever existed, along many lines of descent converging to the common progenitor of the order, would have to be called into life. It is due to their absence, and to the consequent wide gaps in the series, that we are enabled to divide the existing species into
definable groups, such as genera, families, and tribes. If there had been no extinction, there would still have been great lines, or branches, of special development,—the Vandeæ, for instance, would still, as a great body, have been distinguishable from the great body of the Ophreæ; but ancient and intermediate forms, very different probably from their present descendants, would have rendered it utterly impossible to separate by distinct characters the one great body from the other.
I will venture on only a few remarks. Cypripedium, in having three stigmas developed, and therefore in not having a rostellum, in having two fertile anthers with a large rudiment of a third, and in the state of its pollen, seems a remnant of the order whilst in a simpler condition. Apostasia is a related genus, placed by Brown amongst Orchids, but by Lindley in a small distinct family. These broken groups do not indicate to us the structure of the common parent-form of all Orchids, but they probably serve to show the state of the order in ancient times, when none of the forms had become so widely differentiated from each other and from other
plants, as are the existing Orchids, especially the Vandeæ and Ophreæ; and when, consequently, the order made a nearer approach in all its characters, than at present, to such allied groups as the Marantaceæ.
With respect to other Orchids, we can see that an ancient form, like one of the Pleurothallidæ, some of which have waxy pollen-masses with a minute caudicle, might give rise, by the entire abortion of the caudicle, to the Dendrobidæ, and by an increase of the caudicle to the Epidendreæ. Cymbidium shows us how simply a form like one of our present Epidendreæ could be modified into one of the Vandeæ. The Neotteæ stand in nearly a similar relation to the Ophreæ, which the Epidendreæ do to the Vandeæ. In certain genera of the Neotteæ we have the pollen-grains cemented into packets, tied together by elastic threads, which project and form a nascent caudicle. But this caudicle does not protrude from the lower end of the pollinium as in the Ophreæ, nor does it always protrude from the extreme upper end in the Neotteæ; so that we can see that a transition in this respect would be far from impossible.
In Spiranthes, the back of the rostellum, lined with viscid matter, is alone removed; the front part is membranous, and ruptures like the pouch-formed rostellum of the Ophreæ. An ancient form, combining most of the characters, but in a less developed state, of Goodyera, Epipactis, and Spiranthes, would, by further slight modifications, give rise to the whole tribe of the Ophreæ.
Hardly any question in Natural History is more vague and difficult to decide than what forms within any large group ought to be considered as the highest;* for all are excellently adapted to their conditions of life. If we look to traces of successive modification, with differentiation of parts and consequent complexity of structure, as the standard of comparison, the Ophreæ and Vandeæ will stand the highest. Are we to lay much stress on the size and beauty of the flower, and on the size of the whole plant? if so, the Vandeæ
* The fullest and the most able discussion on this difficult subject is by Professor H. G. Bronn in his 'Entwickelungs-Gesetze der Organischen Welt, ' 1858. I have read the French translation published in 1861, Supplement, 'Comptes Rendus,' tom. ii. p. 520 et seq. This great work was crowned by the French Academy of Sciences.
are pre-eminent. They have, also, rather more complex pollinia, with the pollen-masses often reduced to two. The rostellum, on the other hand, in the Ophreæ, has apparently been more modified from its primordial stigmatic nature than in the Vandeæ. In all the Ophreæ the anthers of the inner whorl are almost enterily suppressed,—the auricles—mere rudiments of rudiments—being retained: these anthers have, therefore, undergone a greater amount of modification; but can such suppression be considered as a sign of highness? I should doubt whether any member of the Orchidean order has been more profoundly modified in its whole structure than Bonatea speciosa, one of the Ophreæ. So again, within this tribe, nothing can be more perfect than the contrivances in Orchis pyramidalis for its fertilisation. Yet an ill-defined feeling tells me to rank the magnificent Vandeæ as the highest. When we look within this tribe at the elaborate mechanism of Catasetum for the ejection and transportal of the pollinia, with the sensitive rostellum so wonderfully modified, with the sexes borne on distinct plants, we may
perhaps give to this genus the palm of victory.
A few miscellaneous points, which could not elsewhere have been conveniently introduced, deserve to be noticed. First, for the mechanism by which the pollinia in so many Orchids undergo a movement of depression, when removed from their places of attachment and exposed for a few seconds to the air. This is always due to the contraction of a portion, sometimes, as in Orchis, to an exceedingly minute portion, of the exterior surface of the rostellum, which has retained a membranous condition. This membrane, as we have seen, is likewise sensitive to a touch. In one Maxillaria the middle of the pedicel, and in Habenaria the whole drum-like pedicel, contracts. The point of contraction in all other cases seen by me is either close to the surface of attachment of the caudicle to the disc, or at the point where the pedicel is united to the disc; but both the disc and pedicel are parts of the exterior surface of the rostellum. In these remarks I do not refer to the movements simply due to the elasticity of the pollinia in the Vandeæ.
The long strap-formed disc of Gymnadenia conopsea is well adapted to show the mechanism of the movement of depression. The whole pollinium, both in its upright and depressed (but not closely depressed) position, has been shown by Fig. X. The disc, highly magnified, in its uncontracted condition, is seen from above in the upper figure here given, with the caudicle removed; and in the lower figure we have a longitudinal section of the uncontracted disc, together with the base of the attached and
Fig. XXXIV.
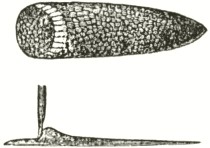
Disc of Gymnadenia conopsea.
upright caudicle. At the broad end of the disc there is a deep crescent-shaped depression, bordered by a slight ridge formed of elongated cells. The end of the caudicle is attached to the steep sides of this depression and ridge. Now, when the disc is exposed to the air for about thirty seconds, the ridge contracts and sinks flat down; in sinking, it drags with it the caudicle. When placed again in water the ridge rises, and when re-exposed to the air it sinks, but each time with
somewhat enfeebled power. With each sinking and rising of the caudicle, the whole pollinium is depressed and elevated.
That the power of movement lies exclusively in the surface of the rostellum is well shown by the saddle-shaped disc of Orchis pyramidalis; for I removed under water the attached caudicles, as well as the layer of viscid matter from its under surface, and immediately that it was exposed to the air the proper contraction ensued. The disc is here formed of several layers of minute cells (and I believe this to be the case with the disc of the Gymnadenia), which are best seen in specimens kept in spirits of wine, for their contents are thus rendered more opaque. The cells in the flaps of the saddle are a little elongated. As long as the saddle is kept damp its upper surface is nearly flat, but when exposed to the air (see Fig. III. E) the surface contracts immediately beneath the point of attachment of the truncated end of each caudicle, and becomes oblique; and two valleys are likewise formed in front of the two caudicles. By this contraction the caudicles are thrown down, almost in the
same way as if trenches were dug in front of two upright poles, and the ground at the same time undermined beneath them. As far as I could perceive, an analogous contraction causes the depression of the pollinia in Orchis mascula.
Some pollinia which had been gummed on card for several months, when placed in water, underwent the movement of depression; and a fresh pollinium, when alternately damped and exposed to the air, can be made to rise and sink several times. Before I had ascertained these facts, which seem to show that the movement is hygrometric, I thought that it was a vital action, and tried vapour of chloroform and prussic acid, and immersion in laudanum; but these reagents did not check the movement. Nevertheless, there are considerable difficulties in understanding how the movement can be simply hygrometric. The flaps of the saddle in Orchis pyramidalis (see Fig. III. D) curl completely inwards in nine seconds, which is surprisingly quick for the action of mere evaporation; and it is the under surface which curls inwards and ought to dry so quickly; but this cannot
happen, as it is covered with a thick layer of viscid matter: the edges, however, of the saddle might become slightly dry in the nine seconds. When the saddle-formed disc is placed in spirits of wine it contracts energetically, and when placed in water opens again. This does not look as if the action was simply hydrometric. Whether the contraction is hygrometric, or is due to endosmose, or to some other unknown cause, the movements of depression in the pollinia thus produced are admirably regulated in each species, so that the pollen-masses, when transported by insects from flower to flower, should assume a position fitted to strike the stigmatic surface.
These movements would be quite useless, unless the pollinia first became attached to the insect in a uniform position relatively to the flower, so as to become after the movement of depression invariably directed towards the stigma; and this necessitates that insects should be led to visit all the flowers of the same species in a uniform manner. Hence I must say a few words on the sepals and petals. Their primary function, no doubt, is to protect the organs of fructification in
the bud. Even after the flower has fully expanded, the upper sepal and two upper petals often continue the same office. We cannot doubt that this protection is of service, when we see in Stelis the sepals so neatly closing and reprotecting the flower after its expansion; in Masdevallia the sepals soldered together, with two little windows alone left open; and when we see, in the open and exposed flowers of the Bolbophyllum, that the mouth of the stigmatic chamber after a time closes. Analogous facts in Malaxis, Cephalanthera, &c., could be given. But the hood formed by the upper sepal and two upper petals, besides affording protection, evidently forms a guide, compelling insects to visit the flower in front. I do not believe that C. K. Sprengel's view,* that the bright and conspicuous colour of the flower serves to attract insects from a distance, is a fanciful notion;
* I am aware that this author's curious work, with its curious title of 'Das Entdeckte Geheimniss der Natur,' has often been spoken lightly of. No doubt he was an enthousiast, and probably carried some of his ideas to an extreme length. I feel sure, from my own observations, that his work contains a large body of truth. Many years ago Robert Brown, to whose judgement all botanists defer, spoke highly of it to me, and remarked that only those who knew little of the subject would laugh at this work.
though some Orchids have singular inconspicuous and greenish flowers,—perhaps in order to escape some danger. Many of these inconspicuous flowers are, however, strongly scented, which would equally well serve to attract insects.
But the labellum is by far the most important of the external envelopes of the flower. It secretes, and often collects in a receptacle, nectar; or is fleshy, and is furnished with excrescences, which probably are attractive to insects. Unless the flowers were by some means rendered attractive, they would be cursed with perpetual sterility. The labellum always stands in front of the rostellum, and its outer portion often serves, as I have seen, as a landing-place for the necessary visitors: in Epipactis palustris this part is flexible and elastic, and apparently compels insects in retreating to brush against the rostellum: in Cypripedium this end is folded over like the end of a slipper, and compels insects to insert their probosces over and near the anthers. In the older flowers of Spiranthes the labellum moves from the column, and leaves a wider passage for the safe introduc-
tion of the pollinia, when attached to the proboscis of a bee. In certain exotic Orchids the labellum suddenly moves and catches insects as if in a box. In Mormodes ignea it is perched on the summit of the column, and here the insects alight and touch the sensitive hinge of the anther. The labellum is often deeply channelled, or has guiding ridges, or is pressed closely against the column, and in a multitude of cases it approaches closely enough to render the flower tubular. By these several means insects are forced to brush against the rostellum. We must not, however, suppose that every detail of structure in the labellum is of use: in some instances, as in Sarcanthus, part of its extraordinary shape seems due to its having been developed in the bud in close apposition to the curiously shaped rostellum.
In Listera ovata the labellum stands far from the column, but its base is narrow, so that insects are led to stand exactly beneath the middle of the rostellum: in other cases, as in Stanhopea, Phalænopsis, &c., the labellum is furnished with upturned lobes, which manifestly act as lateral guides. In some cases, as in Malaxis, the two upper petals are
curled backwards so as to be out of the way; in other cases, as in Acropera, Masdevallia, and some Bolbophyllums, these upper petals plainly serve as lateral guides, compelling insects to visit the flower, or to insert their probosces directly in front of the rostellum. In other cases, wings from the margins of the clinandrum, or from the sides of the column, serve as lateral guides both in the withdrawal of the pollinia, and in their subsequent insertion into the stigmatic cavity. So that there can be no doubt that the petals and sepals and rudimentary anthers do good service in several ways, besides in affording protection to the bud.
The final end of the whole flower, with all its parts, is the production of seed; and these are produced by Orchids in vast profusion. Not that this is anything to boast of in the order; for the production of an almost infinite number of eggs, or seeds, is undoubtedly a sign of lowness of organisation. That a plant, not an annual, should escape destruction at some period of its life simply by the production of a vast number of seeds or seedlings, shows a poverty of contrivance, or a want of
some fitting protection against some danger. I was curious to estimate the number of seeds produced by Orchids; so I took a ripe capsule of Cephalanthera grandiflora, and arranged the seeds as equably as I could in a narrow hillock, on a long ruled line; and then counted the seeds in a length, accurately measured, of one-tenth of an inch. They were 83 in number, and this would give the whole capsule 6020 seeds; and for the four capsules borne by the plant 24,000 seeds. Estimating in the same manner the smaller seeds in Orchis maculata, I found the number nearly the sime, viz. 6200; and, as I have often seen above 30 capsules on the same plant, the total amount will be 186,300,—a prodigious number for a small plant to bear. As this Orchid is perennial, and cannot in most places be increasing in number, one seed alone of this large number, once in every few years, produces a mature plant. I examined many seeds of the Cephalanthera, and very few seemed bad.
To give an idea what the above figures really mean, I will briefly show the possible rate of increase of O. maculata: an acre of
land would hold 174,240 plants, each having a space of six inches square, which is rather closer than they could flourish together; so that, allowing twelve thousand bad seeds, an acre would be thickly clothed by the progeny of a single plant. At the same rate of increase, the grandchildren would cover a space slightly exceeding the island of Anglesea; and the great grandchildren of a single plant would nearly (in the proportion of 47 to 50) clothe with one uniform green carpet the entire surface of the land thoughout the globe.
What checks this unlimited multiplication cannot be told. The minute seeds within their light coats are well fitted for wide dissemination; and I have several times observed seedlings in my orchard, and in a newly-planted wood, which must have come from some little distance. Yet it is notorious that Orchids are sparingly distributed; for instance, this district is highly favourable to the order, for within a mile of my house nine genera, including thirteen species, grow; but of these one alone, Orchis morio, is sufficiently abundant to make a conspicuous feature in the vegetation; as is O. maculata
in a lesser degree in open woodlands. Most of the other species, though not deserving to be called rare, are sparingly distributed; yet, if their seeds or seedlings were not largely and habitually destroyed, any one of them would, as we have just seen, immediately cover the whole land.
I have now nearly finished this too lengthy volume. It has, I think, been shown that Orchids exhibit an almost endless diversity of beautiful adaptations. When this or that part has been spoken of as contrived for some special purpose, it must not be supposed that it was originally always formed for this sole purpose. The regular course of events seems to be, that a part which originally served for one purpose, by slow changes becomes adapted for widely different purposes. To give an instance: in all the Ophreæ, the long and nearly rigid caudicle manifestly serves for the application of the pollen-grains to the stigma, when the pollinium attached to an insect is transported from flower to flower; and the anther opens widely that the pollinium may be easily withdrawn; but in the Bee Ophrys, the caudicle, by a slight increase in
length, and decrease in thickness, and by the anther opening a little more widely, becomes specially adapted for the very different purpose of self-fertilisation, through the combined aid of the gravity of the pollen-mass and the vibration of the flower. Every gradation between these two states would be possible,—of which we have seen partial proof in O. arachnites.
Again the elasticity of the pedicel of the pollinium in some Vandeæ is adapted to free the pollen-masses out of their anther-cases; but by further slight modifications, the elasticity of the pedicel becomes specially adapted to shoot out the pollinia to a distance. The great cavity in the labellum of many Vandeæ serves to attract insects, but in Mormodes ignea it is greatly reduced in size, and only serves to keep the labellum in its proper position on the summit of the column. From the analogy of many plants we may infer that a spur-like nectary is primarily adapted to secrete and hold a store of nectar; but in many Orchids it has so far lost this function, as only to contain fluid between its two coats. In those Orchids, in which the nectary con-
tains both free nectar and fluid in the intercellular spaces, we can see how a passage from one state to the other could have been effected, namely, by less and less nectar being secreted from the inner membrane, and more and more being retained within the intercellular spaces. Other analogous cases could be given.
Although an organ may not have been originally formed for some special purpose, if it now serves for this end we are justified in saying that it is specially contrived for it. On the same principle, if a man were to make a machine for some special purpose, but were to use old wheels, springs, and pulleys, only slightly altered, the whole machine, with all its parts, might be said to be specially contrived for that purpose. Thus throughout nature almost every part of each living being has probably served, in a slightly modified condition, for diverse purposes, and has acted in the living machinery of many ancient and distinct specific forms.
In my examination of Orchids, hardly any fact has so much struck me as the endless diversity of structure,—the prodigality of resources,—for gaining the very same end,
namely, the fertilisation of one flower by the pollen of another. The fact to a certain extent is intelligible on the principle of natural selection. As all the parts of a flower are co-ordinated, if slight variations in any one part are preserved from being beneficial to the plant, then the other parts will generally have to be modified in some corresponding manner. But certain parts may not vary at all, or may not vary in the simplest corresponding manner, and those variations, whatever their nature may be, which will bring all the parts into more perfect harmony with each other, will be seized on and preserved by natural selection.
To give a simple illustration: in many Orchids the ovarium (but sometimes the footstalk) becomes for a period twisted, causing the labellum to hang downwards, so that insects can easily visit the flower; but from slow changes in the form and position of the petals, or from new sorts of insects visiting the flower, it might become advantageous to the plant that the labellum should resume its normal upward position, as is actually the case with Malaxis paludosa; this change, it is obvious, might be simply effected by the continued
selection of varieties which had their ovarium a little less twisted; but if the plant only afforded varieties with the ovarium more twisted, the same end could be attained by their selection until the flower had turned completely round on its axis: this seems to have occurred with the Malaxis, for the labellum has acquired its present upward position, and the ovarium is twisted to excess.
Again, we have seen that in most Vandeæ there is a plain relation between the depth of the stigmatic chamber and the length of the pedicel, by which the pollen-masses are inserted; now if the chamber became slightly less deep from any change in the form of the column or any other unknown cause, the shortening of the pedicel would be the simplest corresponding change; but if the pedicel did not happen to vary in lenth, and the slightest tendency to an upward curvature from elasticity as in Phalænopsis, or to a backward hygrometric movement as in one of the Maxillarias, would be preserved, and the tendency would be continually augmented by selection; thus the pedicel, as far as its action is concerned, would be modified in the same manner
as if it had been shortened. Such processes carried on during many thousand generations in various ways, with the several parts of the flower, would create an endless diversity of coadapted structures for the same general purpose. This view affords, I believe, the key which partly solves the problem of the vast diversity of structure adapted for closely analogous ends in many large groups of organic beings.
The more I study nature, the more I become impressed with ever-increasing force with the conclusion, that the contrivances and beautiful adaptations slowly acquired through each part occasionally varying in a slight degree in many ways, with the preservation or natural selection of those variations which are beneficial to the organism under the complex and ever-varying conditions of life, transcend in an incomparable degree the contrivances and adaptations which the most fertile imagination of the most imaginative man could suggest with unlimited time at his disposal.
The use of each trifling detail of structure is far from a barren search to those who believe
in natural selection. When a naturalist casually takes up an organic being, and does not study its whole life (imperfect though that study will ever be), he naturally doubts whether each trifling point can be of any use, or indeed whether it be due to any general law. Some naturalists believe that numberless structures have been created for the sake of mere variety and beauty,—much as a workman would make a set of different patterns. I, for one, have often and often doubted whether this or that detail of structure could be of any service; yet, if of no good, these structures could not have been modelled by the natural preservation of useful variations; such details could only be vagely accounted for by the direct action of the conditions of life, or the mysterious laws of correlation of growth.
To give nearly all the instances of trifling details of structure in the flowers of Orchids, which are certainly of high importance, would be to recapitulate a great portion of this volume. But I will recall to the reader's memory a few cases. I do not here refer to the fundamental framework of the plant, such as the remnants of the fifteen primary organs
arranged alternately in the five worls; for nearly all those who believe in the modification of organic beings will admit that their presence is due to inheritance from a remote parent-form. A series of facts with respect to the use of the variously shaped and placed petals and sepals has just been enumerated. So, again, the importance of the slight differences in the shape of the caudicle of the pollinium of the Bee Ophrys, compared with that of the other species of the genus, has just been referred to: to this might be added the doubly-bent caudicle of the Fly Ophrys: indeed, the important relation of the length and shape of the caudicle, with reference to the position of the stigma, might be cited throughout whole tribes. The solid projecting knob of the anther in Epipactis palustris, which does not include pollen, when moved by insects, liberates the pollen-masses. In Cephalanthera grandiflora, the upright position of the flower, and its almost closed condition, protect from disturbance the slightly coherent pillars of pollen. The length and elasticity of the filament of the anther in certain species of Dendrobium apparently serves for self-fertilisation, if in-
sects fail to transport the pollen-masses. The slight forward inclination of the crest of the rostellum in Listera prevents the anther-case being caught when the viscid matter explodes. The elasticity of the lip of the rostellum in Orchis causes it to spring up again when one pollen-mass is removed, thus keeping ready for action the second viscid disc, which otherwise would be wasted. The two nectar-secreting spots in the Frog Orchis, beneath the viscid discs at the base of the labellum, and the medial nectary in front of the stigma, are apparently necessary for the fertilisation of the flower. No one who had not studied Orchids would have suspected that these and many other small details of structure were of the highest importance to each species; and that consequently, if the species were exposed to new conditions of life, and the structure of the several parts varied ever so little, such small details of structure might be modified by natural selection. These cases afford a good lesson of caution with respect to the importance, in other organic beings, of apparently trifling particulars of structure.
It may naturally be inquired, why do
Orchids exhibit so many perfect contrivances? From the observations of C. K. Sprengel, and from my own, I am sure that many other plants offer, in the means of fertilisation, analogous adaptations of high perfection; but it seems that they are really more numerous with Orchids than with most other plants. To a certain extent the inquiry can be answered. As each ovule requires at least one, probably more than one, pollen-grain,* and as the seeds produced by Orchids are so inordinately numerous, we can see that large masses of pollen would have to be left on the stigma of each flower. Even in the Neotteæ, which have granular pollen, with the grains tied together only by weak threads, I have observed that considerable masses of pollen are generally left on the stigmas. Hence we can perhaps understand the use of the grains cohering in large waxy masses, as they do in so many tribes, so as to prevent waste in the act of transportal. Most plants produce pollen enough to fertilise several flowers, even when each flower has several stigmas. But as the two
* Gärtner, 'Beiträge zur Kenntniss der Befruchtung, ' 1844, s. 135.
confluent stigmas of Orchids require so much pollen, its elaboration, if proportional in amount to that in most other plants, would have been extravagant in the highest degree, and exhausting to the individual. To save this waste and exhaustion, special and admirable contrivances were necessary for safely placing the pollen-masses on the stigma; and thus we can partially understand why Orchids have been more highly endowed in this respect than most other plants.
The simple fact that many Vandeæ have only two pollen-masses, and that, from their coherence, some Malaxeæ have only a single pollen-mass, necessitates that extraordinary pains should have been taken in their fertilisation, otherwise these plants would have been barren. The existence of a single pollen-mass, so that all the pollen-grains from one flower cannot possibly fertilise more than a single stigma, occurs, I believe, in no other plants. The case is partially analogous with that of seeds: many flowers produce a multitude of seeds, several profuce a single seed; very many flowers produce a countless number of pollen-grains; some Orchid-flowers produce,
as far as the power of fertilising other flowers is concerned, a single pollen-mass, though really consisting of a multitude of pollen-grains.
Notwithstanding that such care has been taken that the pollen of Orchids should not be wasted, we see that throughout the vast Orchidean order,—including, according to Lindley,* 433 genera, and probably about 6000 species,—the act of fertilisation is almost invariably left to insects. This assertion can hardly be considered rash, after the examination of so many British and exotic genera scattered through the main Tribes, which generally have a nearly uniform structure. In all plants in which insects play an important part in the act of fertilisation, there will be a good chance of pollen being carried from one flower to another. But in Orchids we have seen numerous adaptations,—such as the movements of the pollinia after their removal in order the acquire a proper position,—the slow movement of the labellum or rostellum to allow the entrance of the pollen-masses,—the separation of the sexes in some
* 'Gardener's Chronicle,' March 1, 1862, p. 192.
instances—which render it certain that in these cases the pollen of one flower or of one plant is habitually transported to another flower or plant. As this transportal increases the risk of loss, it necessitates and still further explains the extraordionary care bestowed on the contrivances of fertilisation.
Self-fertilisation is a rare event with Orchids. In Cephalanthera grandiflora it occurs, but in a very simple degree; and the early penetration of the stigma by the flower's own pollen-tubes seems to be fully as much determined by the support thus given to the pillars of pollen, as by the production of a small proportion of seed: certainly the fertilisation of this Orchid is aided by insects. In some species of Dendrobium self-fertilisation apparently occurs, but only if insects accidentally fail in removing the flower's own single pollen-mass. In Cypripedium, the Frog Orchis, and perhaps in a few other cases, it will depend on the manner (at present unknown) in which insects first insert their probosces by the one or the other entrance,—whether the flower's own pollen, or that of another flower, is habitually placed on the stigma; but in
these cases there will assuredly always be a good chance of the stigma being fertilised by pollen brought from another flower. In the Bee Ophrys alone, as far as I have seen, there are special and perfectly efficient contrivances for self-fertilisation, combined, however, in the most paradoxical manner, with manifest adaptations for the occasional transport by insects of the pollinia from one flower to another, as in the other species of the same genus.
Considering how precious the pollen of Orchids evidently is, and what care has been bestowed on its organisation and on the accessory parts;—considering that the anther always stands close behind or above the stigma, self-fertilisation would have been an incomparably safer process than the transportal of the pollen from flower to flower. It is an astonishing fact that self-fertilisation should not have been an habitual occurrence. It apparently demonstrates to us that there must be something injurious in the process. Nature thus tells us, in the most emphatic manner, that she abhors perpetual self-fertilisation. This conclusion seems to be of high
importance, and perhaps justifies the lengthy details given in this volume. For may we not further infer as probable, in accordance with the belief of the vast majority of the breeders of our domestic productions, that marriage between near relatives is likewise in some way injurious,—that some unknown great good is derived from the union of individuals which have been kept distinct for many generations?
INDEX.
A.
Aceras anthropophora, 19.
Aceras, monstrous flowers of, 324.
Acontia luctuosa with pollen-masses, 37.
Acropera, structure of flower, 203.
Acropera, movements of pollinia, 189.
Acropera, vessels of, 296.
Adaptations, apparently of small importance, 351.
Adaptations, diversity of, 348.
Adaptations, how far special, 346.
Aerides, nectary of, 276.
Aerides, movement of pollinia, 189, 192.
Angræcum, nectary of, 276.
Angræcum, pollinia of, 186, 188.
Angræcum sesquipedale, 197.
Antennæ of the rostellum in Catasetum, 220, 225.
Anthers, rudimentary, 299.
Apostasia, 308.
Apostasia, relations of, 331.
Arethuseæ, 269.
B.
Babington, Prof., on the rostellum, 320.
Baillon, M., on Catasetum, 229.
Barkeria, 164.
Bateman, Mr., obligations to, 114, 158, 197.
Bauer, Mr., on pollen-grains of Cephalanthera, 104.
——on pollen-masses of Bletia, 162.
——on utriculi of stigma, 206.
Bee Ophrys, 63.
Bentham, Mr., on monstrous flowers of Orchis pyramidalis, 47.
Bletia, 164.
Bolbophyllum, 169.
Bonatea speciosa, 87.
Bonatea speciosa, pollinia of, 330.
Bonatea speciosa, vessels of, 302.
Bond, Mr. F., on moths with attached pollinia, 35.
Brongniart, M., on Catasetum, 236.
——on secretion of nectar, 51.
——on Uropedium, 297.
——on vessels of Orchids, 290.
Bronn, Prof., on the grade of organic beings, 333.
Brown, R., on the fertilisation of Orchids, 3.
——on the viscidity of the stigma, 17.
——on Ophrys apifera, 65.
——on the utriculi of the stigma, 206.
——on homologies of Orchids, 289.
——on the rostellum of Orchids, 308.
——on Apostasia, 331.
Butterfly orchis, 83.
Butterflies with attached pollinia, 36.
C.
Calæna, 172.
Calanthe, structure of flower, 195.
Calanthe, nectary of, 280, 283.
Carpenter, Dr., on the forms of Catasetum, 236.
Catasetidæ, 211.
Catasetum callosum, 231.
Catasetum saccatum, 214.
Catasetum tridentatum, 231.
Catasetum tridentatum a male Orchid, 237.
Catasetum, spiral vessels of, 296.
Catasetum, curious form of the rostellum, 322.
Cattleya, structure of flower, 160.
Cattleya, vessels of, 296.
Caudicles of pollinia in the Vandeæ, 182.
Caudicles, development of, 315.
Caudicle, structure of, 327.
Cephalanthera grandiflora, 104.
Cephalanthera grandiflora, how structure might have been acquired, 155.
Cephalanthera grandiflora, vessels of, 296.
Cephalanthera grandiflora, number of seed, 344.
Clinandrum, vessels of, 298.
Clogyne, 164.
Contrivances apparently of small importance, 351.
Contrivances, how far special, 346.
Coryanthes, nectary of, 277.
Cycnoches, 268.
Cymbidium, 188.
Cymbidium, structure of pollinia, 314, 327.
Cypripedium, structure of flower, 270.
Cypripedium, secretion from hairs, 279.
Cypripedium, vessels of, 296.
Cypripedium, pollen of, 326.
Cypripedium, relations of, 331.
D.
Dendrobium, structure of, 172.
Dendrobidæ, relations of, 332.
Dickie, Prof., obligations to, 152.
Disc, viscidity of, in the Ophreæ, 51.
—— ——in the Vandeæ, 281.
Disc of Catasetum, viscidity of, 229.
Discs double in Ophreæ, 319.
Diversity of structure for same purpose, 348.
Ducts of Orchids, 290.
E.
Epidendreæ, 159.
Epidendreæ, relations of, 332.
Epidendrum, structure of, 165.
——minute structure of rostellum, 311.
Epipactis latifolia, structure of, 102.
——transplanted, 38.
Epipactis palustris, structure of, 95.
Epipactis, vessels of, 296.
Epipogium, 270.
Eulophia, 189.
Eulophia, nectary of, 283.
Evelyna, 164.
Evelyna, nectary of, 277.
Evelyna, vessels of, 296.
F.
Fertilisation, summary on, 357.
Fertility of English Orchids, 39.
Flowers, use of external envelopes, 339.
Fly Ophrys, 54.
Frog orchis, structure of flowers, 76.
——secretion of nectar, 52, 77.
G.
Galeandra, 188.
Gärtner on viscid matter of stigma, 310.
Glossodia, 293.
Goodyera repens, 112.
Goodyera, vessels of, 296.
Goodyera discolor, 114, 209.
Gordon, Rev. G., obligations to, 112.
Gradation of organs, 307.
Gray, Prof. Asa, on structure of Spiranthes, 123.
Gymnadenia albida, 82.
Gymnadenia conopsea, structure of, 79.
——secretion of nectar, 50, 52.
——vessels of, 297.
—— movements of the pollinia, 336.
H.
Habenaria bifolia, 88.
Habenaria chlorantha, 83.
Habenaria viridis, 76.
Habenaria, secretion of nectar, 50, 52.
Habenaria, vessels of, 297, 302.
Herminium monorchis, 74.
Homologies of Orchids, 286.
Hooker, Dr., on Listera, 3.
——obligations to, 158.
——on structure of Listera ovata, 139.
Horwood, Mr., assistance from, 158.
I.
Insects, frequency of visits to Orchids, 39.
Irmisch on flower-bud of Cypripedium, 297.
——Neottia, 153.
——Epipogium, 270.
K.
Krünitz, secretion of nectar, 45.
Kurr, on nectar secreted from bracteæ, 278.
L:
Labellum, easily vibratile, 171.
——Sarcanthus, 281.
——excrescences of, 283.
——cup of, not secreting nectar in the Vandeæ, 283.
——vessels of, 294, 302.
——homologies of, 302.
——important uses of, 341.
Lælia, 164.
Laurel, secretion of nectar from leaves, 278.
Lepidoptera with attached pollinia, 36.
Limodorum, 270.
Lindley, Dr., obligations to, 158.
——arrangements of Orchids, 159.
——on forms of Catasetum, 236.
——on forms of Cycnoches, 269.
——on homologies of Orchids, 290.
——on Apostasia, 331.
——on number of genera and species, 357.
Link on homologies of Orchids, 290, 295.
Liparis, vessels of, 296.
Listera cordata, 152.
Listera ovata, structure of, 139.
Lycaste, 188.
——pollen of, 327.
M.
Malaxeæ, 166.
Malaxis paludosa transplanted, 38.
Malaxis paludosa, 130.
Malaxis, vessels of, 296, 298.
Malaxis, clinandrum of, 298.
Malden, Rev. B. S., obligations to, 43, 78.
Male flowers of Acropera, 203; of Catasetum, 237.
Marshall, Mr., on sterility of transplanted Orchids, 38.
Masdevallia fenestrata, 168.
Maxillaria, movement of pollinia, 189, 191.
Ménière, M., on insects visiting Orchids, 35.
——on movements of Catasetum, 225, 229.
——on secretion of nectar, 277, 283.
Miltonia, pollinia of, 186, 188.
Mycrostylis Rhedii, 135,167.
Monachanthus viridis, 239.
More, Mr. A. G., on fertility of Bee Ophrys, 67.
———on Epipactis palustris, 95, 99.
Mormodes ignea, 249.
Mormodes, sp. ... ? 265.
Moths with attached pollinia, 36.
Moths, intellect of, 46.
Movements of the pollinia, 92, 335.
Musk orchis, 74.
Myanthus barbatus, 239, 245.
N.
Nectar, secretion of, by British Ophreæ, 45, 50.
—— —in foreign Orchids, 276, 280.
Nectar secreted from bracteæ, &c., 278.
Nectary of British Ophreæ, 45, 50.
Nectary cut off to test intellect of moths, 47.
Nectary, length of, in Angræcum, 198.
Nectary, vessels of, 294.
Neottia nidus-avis, 152.
Neotteæ, relations of, 332.
Nevill, Lady Dorothy, obligations to, 158.
Nicotiana, stigma of, 310.
O.
Odontoglossum, 189.
Oliver, Prof., obligations to, 158.
Oncidium, pollinia of, 185, 189, 192.
Ophreæ, relations of, 332.
Ophreæ, vessels of, 300.
Ophrys apifera, structure of, 63.
Ophrys apifera, fertility of, 66.
Ophrys arachnitis, 72.
Ophrys aranifera, 60.
Ophrys muscifera, structure of, 55.
—— fertility of, 59.
—— transplanted, 38.
Ophrys fusca, 19.
O. fusca, sterility of, 43.
Orchis latifolia, 19.
O. latifolia, fertility of, 42.
——secretion of nectar, 45.
Orchis maculata, 19.
O. maculata transplanted, 38.
——number of seed, 344.
Orchis mascula, 9, 34.
O. mascula, movement of pollinia, 16.
Orchis morio, 19, 33.
——fertility of, in cold season, 39.
——secretion of nectar, 45, 48.
Orchis pyramidalis, 20.
——secretion of nectar, 46, 49.
—— fertility of, in different stations, 39.
——monstrous flowers of, 47.
—— vessels of, 296.
——movements of the pollinia, 26, 337.
Orchis ustulata, 31.
Oxenden, Mr. G. C., obligations to, 31.
P.
Parfitt, Mr., on attached pollinia, 36.
Parker, Mr. R., obligations to, 158.
Pedicel of pollinium, 317.
Percy, Dr., analysis of labellum, 284.
Peristylus viridis, 76.
——,secretion of nectar, 52, 77.
Petals, uses of, 339.
Phaius, 164.
Phalænopsis, viscidity of stigma, 183.
Phalænopsis, movements of pollinia, 194.
Pleurothallis, 166.
Pollen-masses, rudimentary in Monachanthus, 241.
——, gradation of, 323.
Pollen-tubes emitted within the anther, 324.
Pollinia, summary on their movements, 91, 335.
Pollinia of the Vandeæ, 185.
Pollinia of Catasetum, ejection of, 218.
Pollinia of Mormodes, ejection of, 256.
Pollinia, attachment to the rostellum, 314.
Pollinia, gradation of, 323.
R.
Rodgers, Mr., obligations to, 158.
——on Myanthus, 236.
Rodriguezia, 189, 193.
Rostellum, a single organ in the Ophreæ, 54, 319.
Rostellum of the Vandeæ, 179.
Rostellum aborted, 301.
Rostellum, gradation of, 307, 312.
Rostellum, minute structure of, 311.
Rostellum, crest of, in the Ophreæ, 320.
Rostellum complex in Catasetum, 322.
Rucker, Mr., obligations to, 158, 214, 231, 249.
S.
Saccolabium, viscidity of stigma, 183.
——, movements of pollinia, 189.
Saint-Hilaire, Aug., on pollen of Orchids, 326.
Sarcanthus, pollinia of, 185, 189.
——, movements of pollinia, 189.
——, labellum of, 281.
Schomburgk, Sir. R., on Catasetum, 236.
Secretion of nectar, 44, 276, 280.
Seed, number of, 343.
Self-fertilisation, summary on, 358.
Sepals, uses of, 339.
Sexes of Orchids, 203, 236.
Smith, Mr. F., on Bombus with attached pollen-masses, 164.
Smith, Sir James, on position of flowers in Malaxis, 131.
Smith, Mr. G., on bees visiting the Bee Ophrys, 68.
Sophronitis, 164.
Spiranthes autumnalis, 116.
—— ——, vessels of, 296.
Sprengel, C. K., sterility of Orchis militaris, 44.
——, secretion of nectar by Orchis, 45.
—— on transportal of pollinia of Epipactis, 103.
—— on Listera, 139, 146.
——, value of work, 340.
Stamens, rudimentary, 299.
Stanhopea, pollinia of, 185.
Stelis, 167.
Sterility of English Orchids, 39.
Stigma, viscidity of, in the Vandeæ, 183.
——, utriculi of, 206, 237, 264.
——, state of, in Catasetum, 237.
——, gradation of, 309, 312.
——, minute structure of, 311.
Structure, diversity of, for same purpose, 348.
Structures of apparently small importance, 351.
T.
Turnbull, Mr. G. H., of Down, obligations to, 158.
U.
Uropedium, 297.
Utriculi of stigma, 206, 237, 264.
V.
Vanda, 189.
Vandeæ, 178.
——, relations of, 332.
Vanilla, fertilisation of, 270.
——, secretion of nectar, 278.
Veitch, Mr., obligations to, 158, 214.
Vessels, spiral, of Orchids, 290.
Vicia, secretion of nectar from stipules, 278.
Viscidity of disc in British Ophreæ, 51.
—— ——Catasetum, 229.
—— ——Vandeæ, 281.
Viscidity of rostellum and stigma, 309, 314.
W.
Wallis, Mr., obligations to, 130.
Warrea, nectary of, 284.
——, pollinia of, 188.
Weddell, Dr., on hybrids of Aceras, 19.
Westwood, Prof., on bees visiting Orchids, 35.
Z.
Zygopetalum, 188.
LONDON:
PRINTED BY WILLIAM CLOWES AND SONS, STAMFORD STREET,
AND CHARING CROSS.
ALBEMARLE STREET, LONDON.
December, 1861.
MR. MURRAY'S
GENERAL LIST OF WORKS.
ABBOTT'S (REV. J.) Philip Musgrave; or, Memoirs of a Church of England Missionary in the North American Colonies. Post 8vo. 2s.
ABERCROMBIE'S (JOHN) Enquiries concerning the Intellectual Powers and the Investigation of Truth Sixteenth Edition. Fcap. 8vo. 6s. 6d.
— Philosophy of the Moral Feelings. Twelfth Edition. Fcap. 8vo 4s.
— Pathological and Practical Researches on the Diseases of the Stomach, &c. Third Edition. Fcap. 8vo. 6s.
ACLAND'S (REV. CHARLES) Popular Account of the Manners and Customs of India. Post 8vo. 2s.
ADOLPHUS'S (J. L.) Letters from Spain, in 1856 and 1857. Post 8vo. 10s. 6d.
ÆSCHYLUS. (The Agamemnon and Choephorœ.) Edited, with Notes. By Rev. W. PEILE, D.D. Second Edition. 2 Vols. 8vo. 9s. each
ÆSOP'S FABLES. A New Translation. With Historical Preface. By Rev. THOMAS JAMES. With 100 Woodcuts, by TENNIEL and WOLF. 38th Thousand. Post 8vo. 2s. 6d.
AGRICULTURAL (THE) JOURNAL. Of the Royal Agricultural Society of England. 8vo. 10s. Published half-yearly.
AIDS TO FAITH: a Series of Theological Essays. 8vo. 9s.
CONTENTS.
Rev. H. L. MANSEL—On Miracles.
BISHOP OF CORK—Christian Evidences.
REV. DR. MCCAUL—On Prophecy.
Rev. F. C. COOK—Ideology and Subscription.
Rev. DR. MCCAUL—Mosaic Record of Creation.
Rev. GEORGE RAWLINSON—The Pentateuch.
BISHOP OF GLOUCESTER AND BRISTOL—Doctrine of the Atonement.
Rev. HAROLD BROWNE—On Inspiration.
DEAN OF EXETER—Scripture and its Interpretation.
AMBER-WITCH (THE). The most interesting Trial for Witch-craft ever known. Translated from the German by LADY DUFF GORDON. Post 8vo. 2s.
ARTHUR'S (LITTLE) History of England. By LADY CALLCOTT. 100th Thousand. With 20 Woodcuts. Fcap.8vo. 2s. 6d.
AUNT IDA'S Walks and Talks; a Story Book for Children. By a LADY. Woodcuts. 16mo. 5s.
AUSTIN'S (JOHN) PROVINCE OF JURISPRUDENCE DETERMINED; or, Philosophy of Positive Law. Second Edition. 8vo. 15s.
— Lectures on Jurisprudence. Being a Continuation of the "Province of Jurisprudence Determined." 8vo. (Nearly Ready.)
— (SARAH) Fragments from German Prose Writers. With Biographical Notes. Post 8vo. 10s.
B
ADMIRALTY PUBLICATIONS; Issued by direction of the Lords Commissioners of the Admiralty:—
A MANUAL OF SCIENTIFIC ENQUIRY, for the Use of Travellers in General. By Various Hands. Edited by Sir JOHN F. HERSCHEL, Bart. Third Edition, revised by Rev. ROBERT MAIN. Woodcuts. Post 8vo. 9s.
AIRY'S ASTRONOMICAL OBSERVATIONS MADE AT GREENWICH. 1836 to 1847. Royal 4to. 50s. each.
— ASTRONOMICAL RESULTS. 1848 to 1858. 4to. 8s. each.
— APPENDICES TO THE ASTRONOMICAL OBSERVATIONS.
1836.—I. Bessel's Refraction Tables. 8s.
II. Tables for converting Errors of R.A. and N.P.D. into Errors of Longitude and Ecliptic P.D. 8s.
1887.—I. Logarithms of Sines and Cosines to every Ten Seconds of Time. 8s.
II. Table for converting Sidereal into Mean Solar Time. 8s.
1842.—Catalogue of 1439 Stars. 8s.
1845.—Longitude of Valentia. 8s.
1847.—Twelve Years' Catalogue of Stars. 14s.
1851.—Maskelyne's Ledger of Stars. 6s.
1852.—I. Description of the Transit Circle. 5s.
II. Regulations of the Royal Observatory. 2s.
1853.—Bessel's Refraction Tables. 3s.
1854.—I. Description of the Zenith Tube. 3s.
II. Six Years' Catalogue of Stars. 10s.
1856.—Description of the Galvanic Apparatus at Greenwich Observatory. 8s.
— MAGNETICAL AND METEOROLOGICAL OBSERVATIONS. 1840 to 1847. Royal 4to. 50s. each.
— ASTRONOMICAL, MAGNETICAL, AND METEOROLOGICAL OBSERVATIONS, 1848 to 1859. Royal 4to. 50s. each.
— ASTRONOMICAL RESULTS. 1859. 4to.
— MAGNETICAL AND METEOROLOGICAL RESULTS. 1848 to 1859. 4to. 8s. each.
— REDUCTION OF THE OBSERVATIONS OF PLANETS, 1750 to 1830. Royal 4to. 50s.
— LUNAR OBSERVATIONS. 1750 to 1830. 2 Vols. Royal 4to. 50s.. each.
— 1831 to 1851. 4to. 20s.
BERNOULLI'S SEXCENTENARY TABLE. London, 1779. 4to.
BESSEL'S AUXILIARY TABLES FOR HIS METHOD OF CLEARING LUNAR DISTANCES. 8vo.
— FUNDAMENTA ASTRONOMIÆ: Regiomontii,1818. Folio.60s.
BIRD'S METHOD OF CONSTRUCTING MURAL QUADRANTS. London, 1768. 4to. 2s. 6d.
— METHOD OF DIVIDING ASTRONOMICAL INSTRUMENTS. London, 1767. 4to. 2s. 6d.
COOK, KING, AND BAYLY'S ASTRONOMICAL OBSERVATIONS. London 1782. 4to. 21s.
EIFFE'S ACCOUNT OF IMPROVEMENTS IN CHRONOMETERS. 4to. 2s.
ENCKE'S BERLINER JAHRBUCH, for 1830. Berlin, 1828. 8vo. 9s.
GROOMBRIDGE'S CATALOGUE OF CIRCUMPOLAR STARS. 4to. 10s.
HANSEN'S TABLES DE LA LUNE. 4to. 20s.
BARRISON'S PRINCIPLES OF HIS TIME-KEEPER. PLATES. 1767. 4to. 5s.
[page] 3 PUBLISHED BY MR. MURRAY.
ADMIRALTY PUBLICATIONS—Continued.
HUTTON'S TABLES OF THE PRODUCTS AND POWERS OF NUMBERS. 1781. Folio. 7s. 6d.
LAX'S TABLES FOR FINDING THE LATITUDE AND LONGITUDE. 1821. 8vo. 10s.
LUNAR OBSERVATIONS at GREENWICH. 1788 to 1819. Compared with the Tables, 1821. 4to. 7s. 6d.
MASKELYNE'S ACCOUNT OF THE GOING OF HARRISON'S WATCH. 1767. 4to. 2s. 6d.
MAYER'S DISTANCES of the MOON'S CENTRE from the PLANETS. 1822, 3s.; 1823, 4s. 6d. 1824 to 1835, 8vo. 4s. each.
— THEORIA LUNÆ JUXTA SYSTEMA NEWTONIANUM. 4to. 2s. 6d.
— TABULÆ MOTUUM SOLIS ET LUNÆ. 1770. 4to. 5s.
— ASTRONOMICAL OBSERVATIONS MADE AT GOTTINGEN, from 1756 to 1761. 1826. Folio. 7s. 6d.
NAUTICAL ALMANACS, from 1767 to 1864. 8vo. 2s. 6d. each.
— SELECTIONS FROM THE ADDITIONS up to 1812. 8vo. 5s. 1834-54. 8vo. 5s.
— SUPPLEMENTS, 1828 to 1838, 1837 and 1838. 8vo. 2s. each.
— TABLE requisite to be used with the N.A. 1781. 8vo. 5s.
POND'S ASTRONOMICAL OBSERVATIONS. 1811 to 1835. 4to. 21s. each.
RAMSDEN'S ENGINE for DIVIDING MATHEMATICAL INSTRUMENTS. 4to. 5s.
— ENGINE for DIVIDING STRAIGHT LINES. 4to. 5s.
SABINE'S PENDULUM EXPERIMENTS to DETERMINE THE FIGURE. OF THE EARTH. 1825. 4to. 40s.
SHEPHERD'S TABLES for CORRECTING LUNAR DISTANCES. 1772. Royal 4to. 21s.
— TABLES, GENERAL, of the MOON'S DISTANCE from the SUN, and 10 STARS. 1787. Folio. 5s. 6d.
TAYLOR'S SEXAGESIMAL TABLE. 1780. 4to. 15s.
— TABLES OF LOGARITHMS. 4to. 3l.
TIARK'S ASTRONOMICAL OBSERVATIONS for the LONGITUDE of MADEIRA. 1822. 4to. 5s.
— CHRONOMETRICAL OBSERVATIONS for DIFFERENCES of LONGITUDE between DOVER, PORTSMOUTH, and FALMOUTH. 1823. 4to. 5s.
VENUS and JUPITER: OBSERVATIONS of, compared with the TABLES. London, 1822. 4to. 2s.
WALES' AND BAYLY'S ASTRONOMICAL OBSERVATIONS. 1777. 4to. 21s.
WALES' REDUCTION OF ASTRONOMICAL OBSERVATIONS MADE IN THE SOUTHERN HEMISPHERE. 1764–1771. 1788. 4to. 10s. 6d.
BABBAGE'S (CHARLES) Economy of Machinery and Manufactures. Fourth Edition. Fcap. 8vo. 6s.
— Ninth Bridgewater Treatise. 8vo. 9s. 6d.
— Reflections on the Decline of Science in England, and on some of its Causes. 4to. 7s. 6d.
B 2
BAIKIE'S (W. B.) Narrative of an Exploring Voyage up the Rivers Quorra and Tshadda in 1854. Map. 8vo. 16s.
BANKES' (GEORGE) STORY OF CORFE CASTLE, with documents relating to the Time of the Civil Wars, &c. Woodcuts. Post 8vo. 10s. 6d.
BASSOMPIERRE'S Memoirs of his Embassy to the Court of England in 1626. Translated with Notes. 8vo. 9s. 6d.
BASTIAT'S (FREDERIC) Harmonies of Political Economy. Translated, with a Notice of his Life and Writings, by P. J. STIRLING. 8vo. 7s. 6d.
BARROW'S (SIR JOHN) Autobiographical Memoir, including Reflections, Observations, and Reminiscences at Home and Abroad. From Early Life to Advanced Age. Portrait. 8vo. 16s.
—Voyages of Discovery and Research within the Arctic Regions, from 1818 to the present time. Abridged and arranged from the Official Narratives. 8vo. 15s.
— (SIR GEORGE) Ceylon; Past and Present. Map. Post 8vo. 6s. 6d.
— (JOHN) Naval Worthies of Queen Elizabeth's Reign, their Gallant Deeds, Daring Adventures, and Services in the infant state of the British Navy. 8vo. 14s.
— Life and Voyages of Sir Francis Drake. With numerous Original Letters. Post 8vo. 2s.
BEES AND FLOWERS. Two Essays. By Rev. Thomas James. Reprinted from the "Quarterly Review." Fcap. 8vo. 1s. each.
BELL'S (SIR CHARLES) Mechanism and Vital Endowments of the Hand as evincing Design. Sixth Edition. Woodcuts. Post 8vo. 6s.
BENEDICT'S (JULES) Sketch of the Life and Works of Felix Mendelssohn-Bartholdy. Second Edition. 8vo. 2s. 6d.
BERTHA'S Journal during a Visit to her Uncle in England. Containing a Variety of Interesting and Instructive Information. Seventh Edition. Woodcuts. 12mo.
BIRCH'S (SAMUEL) History of Ancient Pottery and Porcelain: Egyptian, Assyrian, Greek, Roman, and Etruscan. With 200 Illustrations. 2 Vols. Medium 8vo. 42s.
BLUNT'S (REV. J. J.) Principles for the proper understanding of the Mosaic Writings, stated and applied, together with an Incidental Argument for the truth of the Resurrection of our Lord. Being the HULSEAN LECTURES for 1832. Post 8vo. 6s. 6d.
— Undesigned Coincidences in the Writings of the Old and New Testament, an Argument of their Veracity: with an Appendix containing Undesigned Coincidences between the Gospels, Acts, and Josephus. Sixth Edition. Post 8vo. 7s. 6d.
— History of the Church in the First Three Centuries. Third Edition. Post 8vo. 7s. 6d.
— Parish Priest; His Duties, Acquirements and Obligations. Third Edition. Post 8vo. 7s. 6d.
— Lectures on the Right Use of the Early Fathers. Second Edition. 8vo. 15s.
— Plain Sermons Preached to a Country Congregation. Second Edition. 3 Vols. Post 8vo. 7s. 6d. each.
— Literary and Clerical Essays. 8vo. 12s.
[page] 5 PUBLISHED BY MR. MURRAY.
BLACKSTONE'S COMMENTARIES on the Laws of England. Adapted to the present state of the law. By R. MALCOLM KERR, LL.D. Third Edition, corrected to 1861. 4 Vols. 8vo. 63s.
— For STUDENTS. Being those Portions which relate to the BRITISH CONSTITUTION and the RIGHTS OF PERSONS. Post 8vo. 9s.
BOOK OF COMMON PRAYER. With 1000 Illustrations of Borders, Initials, and Woodcut Vignettes. Medium 8vo. 21s.
BOSWELL'S (JAMES) Life of Dr. Johnson. Including the Tour to the Hebrides. Edited by Mr. CROKER. Portraits. Royal 8vo. 10s.
BORROW'S (GEORGE) Lavengro; The Scholar—The Gipsy—and the Priest. Portrait. 3 Vols. Post 8vo. 30s.
—Romany Rye; a Sequel to Lavengro. Second Edition. 2 Vols. Post 8vo. 21s.
—Bible in Spain; or the Journeys, Adventures, and Imprisonments of an Englishman in an Attempt to circulate the Scriptures in the Peninsula. 3 Vols. Post 8vo. 27s; or Popular Edition, 16mo, 3s. 6d.
— Zincali, or the Gipsies of Spain; their Manners, Customs, Religion, and Language. 2 Vols. Post 8vo. 18s; or Popular Edition, 16mo, 3s. 6d.
BRAY'S (MRS.) Life of Thomas Stothard, R.A. With Personal Reminiscences. Illustrated with Portrait and 60 Woodcuts of his chief works. 4to.
BREWSTER'S (SIR DAVID) Martyrs of Science, or the Lives of Galileo, Tycho Brahe, and Kepler. Fourth Edition. Fcap. 8vo. 4s. 6d.
— More Worlds than One. The Creed of the Philosopher and the Hope of the Christian. Eighth Edition. Post 8vo. 6s.
— Stereoscope: its History, Theory, Construction, and Application to the Arts and to Education. Woodcuts. 12mo. 5s. 6d.
— Kaleidoscope: its History, Theory, and Construction, with its application to the Fine and Useful Arts. Second Edition. Woodcuts. Post 8vo. 5s. 6d.
BRITISH ASSOCIATION REPORTS. 8vo. York and Oxford, 1831-32, 13s. 6d. Cambridge, 1833, 12s. Edinburgh, 1834, 15s. Dublin, 1835, 13s. 6d. Bristol, 1836, 12s. Liverpool, 1837, 16s. 6d. Newcastle, 1838, 15s. Birmingham, 1839, 13s. 6d. Glasgow, 1840, 15s. Plymouth, 1841, 13s. 6d. Manchester, 1842, 10s. 6d. Cork, 1843, 12s. York, 1844, 20s. Cambridge, 1845, 12s. Southampton, 1846, 15s. Oxford, 1847, 18s. Swansea,1848, 9s Birmingham, 1849, 10s. Edinburgh, 1850, 15s. Ipswich, 1851, 16s. 6d. Belfast, 1832, 15s. Hull, 1853, 10s. 6d. Liverpool, 1854, 18s. Glasgow, 1855, 15s.; Cheltenham, 1856, 18s.; Dublin, 1857, 15s.; Leeds, 1858, 20s.; Aberdeen, 1859, 15s. Oxford, 1860. 15s.
BRITISH CLASSICS. A New Series of Standard English Authors, printed from the most correct text, and edited with elucidatory notes. Published occasionally in demy 8vo. Volumes, varying in price.
Already Published.
GOLDSMITH'S WORKS. Edited by PETER CUNNINGHAM, F.S.A. Vignettes. 4 Vols. 30s.
GIBBON'S DECLINE AND FALL OF THE ROMAN EMPIRE. Edited by WILLIAM SMITH, LL.D. Portrait and Maps. 8 Vols. 60s.
JOHNSON'S LIVES OF THE ENGLISH POETS. Edited by PETER CUNNINGHAM, F.S.A. 3 Vols. 22s. 6d.
BYRON'S POETICAL WORKS. Edited, with Notes. 6 vols. 45s.
In Preparation.
WORKS OF POPE. With Life, Introductions, and Notes, by REV. WHITWELL ELWIN. Portrait.
HUME'S HISTORY OF ENGLAND. Edited, with Notes.
LIFE AND WORKS OF SWIFT. Edited by JOHN FORSTER.
BROUGHAM'S (LORD) Address at the Social Science Association, Dublin. August, 1861. Revised, with Notes. 8vo. 1s.
BROUGHTON'S (LORD) Journey through Albania and other Provinces of Turkey in Europe and Asia. to Constantinople, 1809—10. Third Edition. Maps and Woodcuts. 2 Vols. 8vo. 30s.
— Visits to Italy. Third Edition. 2 vols. Post 8vo. 18s.
BUBBLES FROM THE BRUNNEN OF NASSAU. By an Old MAN. Sixth Edition. 16mo. 5s.
BUNBURY'S (C. J. F.) Journal of a Residence at the Cape of Good Hope; with Excursions into the Interior, and Notes on the Natural History and Native Tribes of the Country. Woodcuts. Post 8vo. 9s.
BUNYAN (JOHN) and Oliver Cromwell. Select Biographies. By ROBERT SOUTHEY. Post 8vo. 2s.
BUONAPARTE'S (NAPOLEON) Confidential Correspondence with his Brother Joseph, sometime King of Spain. Second Edition. 2 vols. 8vo. 26s.
BURGHERSH'S (LORD) Memoir of the Operations of the Allied Armies under Prince Schwarzenberg and Marshal Blucher during the latter end of 1813—14. 8vo. 21s.
— Early Campaigns of the Duke of Wellington in Portugal and Spain. 8vo. 8s. 6d.
BURGON'S (Rev. J. W.) Memoir of Patrick Fraser Tytler. Second Edition. Post 8vo. 9s.
— Letters from Rome, written to Friends at Home. Illustrations. Post 8vo. 12s.
BURN'S (LIEUT.-COL.) French and English Dictionary of Naval and Military Technical Terms. Third Edition. Crown 8vo. 15s.
BURNS' (ROBERT) Life. By JOHN GIBSON LOCKHART. Fifth Edition. Fcap. 8vo. 3s.
BURR'S (G. D.) Instructions in Practical Surveying, Topographical Plan Drawing, and on sketching ground without Instruments. Third Edition. Woodcuts. Post 8vo. 7s. 6d.
BUTTMAN'S LEXILOGUS; a Critical Examination of the Meaning of numerous Greek Words, chiefly in Homer and Hesiod. Translated by Rev. J. R. FISHLAKE. Fifth Edition. 8vo. 12s.
[page] 7 PUBLISHED BY MR. MURRAY.
BUXTON'S (SIR FOWELL) Memoirs. With Selections from his Correspondence. By his Son. Portrait. Fifth Edition. 8vo. 16s. Abridged Edition, Portrait. Fcap. 8vo. 2s. 6d.
BYRON'S (LORD) Life, Letters, and Journals. By THOMAS MOORE. Plates. 6 Vols. Fcap. 8vo. 18s.
— Life, Letters, and Journals. By THOMAS MOORE. Portraits. Royal 8vo. 9s.
— Poetical Works. Portrait. 6 Vols. 8vo. 45s.
— Poetical Works. Plates. 10 Vols. Fcap. 8vo. 30s.
— Poetical Works. 8 Vols. 24mo. 20s.
— Poetical Works. Plates. Royal 8vo. 9s.
— Poetical Works. Portrait. Crown 8vo. 6s.
— Childe Harold. With 80 Engravings. Square 8vo. 21s.
— Childe Harold. With 30 Vignettes. 12mo. 6s.
— Childe Harold. 16mo. 2s. 6d.
— Childe Harold. Vignettes. 16mo. 1s.
— Childe Harold. Portrait. 16mo. 6d.
— Tales and Poems. 24mo. 2s. 6d.
— Miscellaneous. 2 Vols. 24mo. 5s.
— Dramas and Plays. 2 Vols. 24mo. 5s.
— Don Juan and Beppo. 2 Vols. 24mo. 5s.
— Beauties. Selected from his Poetry and Prose. Portrait, Fcap. 8vo. 3s. 6d.
CARNARVON'S (LORD) Portugal, Gallicia, and the Basque Provinces. From Notes made during a Journey to those Countries. Third Edition. Post 8vo. 3s. 6d.
— Address on the Archæology of Berkshire. Second Edition. Fcap. 8vo. 1s.
— Recollections of the Druses of Lebanon. With Notes on their Religion. Third Edition. Post 8vo. 5s. 6d.
CAMPBELL'S (LORD) Lives of the Lord Chancellors and Keepers of the Great Seal of England. From the Earliest Times to the Death of Lord Eldon in 1838. Fourth Edition. 10 Vols. Crown 8vo. 6s. each.
— Lives of the Chief Justices of England. From the Norman Conquest to the Death of Lord Tenterden. Second Edition. 3 Vols. 8vo. 42s.
— Shakspeare's Legal Acquirements Considered. 8vo. 5s. 6d.
— Life of Lord Chancellor Bacon. Fcap. 8vo. 2s. 6d.
— (GEORGE) Modern India. A Sketch of the System of Civil Government. With some Account of the Natives and Native Institutions. Second Edition. 8vo. 16s.
— India as it may be. An Outline of a proposed Government and Policy. 8vo. 12s.
— (THOS.) Short Lives of the British Poets. With an Essay on English Poetry. Post 8vo. 3s. 6d.
CALVIN'S (JOHN) Life. With Extracts from his Correspondence. By THOMAS H. DYER. Portrait. 8vo. 15s.
CALLCOTT'S (LADY) Little Arthur's History of England. 100th Thousand. With 20 Woodcuts. Fcap. 8vo. 2s. 6d.
CARMICHAEL'S (A. N.) Greek Verbs. Their Formations, Irregularities, and Defects. Second Edition. Post 8vo. 8s. 6d.
CASTLEREAGH (THE) DESPATCHES, from the commencement of the official career of the late Viscount Castlereagh to the close of his life. Edited by the MARQUIS OF LONDONDERRY. 12 Vols. 8vo. 14s. each.
CATHCART'S (SIR GEORGE) Commentaries on the War in Russia and Germany, 1812-13. Plans. 8vo. 14s.
— Military Operations in Kaffraria, which led to the Termination of the Kaffir War. Second Edition. 8vo. 12s.
CAVALCASELLE (G. B.). Notices of the Early Flemish Painters; Their Lives and Works. Woodcuts. Post 8vo. 12s.
CHAMBERS' (G. F.) Handbook of Descriptive and Practical Astronomy. Illustrations. Post 8vo. 12s.
CHANTREY (SIR FRANCIS). Winged Words on Chantrey's Woodcocks. Edited by JAS. P. MUIRHEAD. Etchings. Square 8vo. 10s. 6d.
CHARMED ROE (THE); or, The Story of the Little Brother and Sister. By OTTO SPECKTER. Plates. 16mo. 5s.
CLAUSEWITZ'S (CARL VON) Campaign of 1812, in Russia. Translated from the German by LORD ELLESMERE. Map. 8vo. 10s. 6d.
CLIVE'S (LORD) Life. By REV. G. R. GLEIG, M.A. Post 8vo. 3s. 6d.
COBBOLD'S (REV. R. H.) Pictures of the Chinese drawn by themselves. With 24 Plates. Crown 8vo. 9s.
COLCHESTER (LORD). The Diary and Correspondence of Charles Abbott, Lord Colchester, Speaker of the House of Commons, 1802-1817. Edited by HIS SON. Portrait. 3 Vols. 8vo. 42s.
COLERIDGE'S (SAMUEL TAYLOR) Table-Talk. Fourth Edition. Portrait. Fcap. 8vo. 6s.
— (HENRY NELSON) Introductions to the Greek Classic Poets. Third Edition. Fcap. 8vo. 5s. 6d.
— (SIR JOHN) on Public School Education, with especial reference to Eton. Third Edition. Fcap. 8vo. 2s.
COLONIAL LIBRARY. [See Home and Colonial Library.]
COOKERY (DOMESTIC). Founded on Principles of Economy and Practical Knowledge, and adapted for Private Families. New Edition. Woodcuts. Fcap. 8vo. 5s.
CORNWALLIS (THE) Papers and Correspondence during the American War,—Administrations in India,—Union with Ireland, and Peace of Amiens. Edited by CHARLES Ross. Second Edition. 3 Vols. 8vo. 63s.
CRABBE'S (REV. GEORGE) Life, Letters, and Journals. By his SON. Portrait. Fcap. 8vo. 3s.
— Poetical Works. With his Life. Plates. 8 Vols. Fcap.8vo. 24s.
— Poetical Works. Plates. Royal 8vo. 7s.
[page] 9 PUBLISHED BY MR. MURRAY.
CURZON'S (HON. ROBERT) Visits to the Monasteries of the Levant. Fourth Edition. Woodcuts. Post 8vo. 15s.
— ARMENIA AND ERZEROUM. A Year on the Frontiers of Russia, Turkey, and Persia. Third Edition. Woodcuts. Post 8vo. 7s. 6d.
CUNNINGHAM'S (ALLAN) Life of Sir David Wilkie. With his Journals and Critical Remarks on Works of Art. Portrait. 3 Vols. 8vo. 42s.
— Poems and Songs. Now first collected and arranged, with Biographical Notice. 24mo. 2s. 6d.
— (CAPT. J. D.) History of the Sikhs. From the Origin of the Nation to the Battle of the Sutlej. Second Edition. Maps. 8vo. 15s.
CROKER'S (J. W.) Progressive Geography for Children. Fifth Edition. 18mo. 1s. 6d.
— Stories for Children, Selected from the History of England. Fifteenth Edition. Woodcuts. 16mo. 2s. 6d.
— Boswell's Life of Johnson. Including the Tour to the Hebrides. Portraits. Royal 8vo. 10s.
— LORD HERVEY'S Memoirs of the Reign of George the Second, from his Accession to the death of Queen Caroline. Edited with Notes. Second Edition. Portrait. 2 Vols. 8vo. 21s.
— Essays on the Early Period of the French Revolution. Reprinted from the Quarterly Review. 8vo. 15s.
— Historical Essay on the Guillotine. Fcap. 8vo. 1s.
CROMWELL (OLIVER) and John Bunyan. By ROBERT SOUTHEY. Post 8vo. 2s.
CROWE'S (J. A.) Notices of the Early Flemish Painters; their Lives and Works. Woodcuts. Post 8vo. 12s.
CURETON (REV. W.) Remains of a very Ancient Recension of the Four Gospels in Syriac, hitherto unknown in Europe. Discovered, Edited, and Translated. 4to. 24s.
CURTIUS' (PROFESSOR) Greek Grammar. Translated from the German. Edited by DR. WM. SMITH. Post 8vo. (In the Press.)
DARWIN'S (CHARLES) Journal of Researches into the Natural History and Geology of the Countries visited during a Voyage round the World. Tenth Thousand. Post 8vo. 9s.
— Origin of Species by Means of Natural Selection; or, the Preservation of Favoured Races in the Struggle for Life. Seventh Thousand. Post 8vo. 14s.
— Various Contrivances by which British and Foreign Orchids are Fertilised through Insect Agency, and as to good of Intercrossing. Woodcuts. Post 8vo. (Nearly Ready.)
DAVY'S (SIR HUMPHRY) Consolations in Travel; or, Last Days of a Philosopher. Fifth Edition. Woodcuts. Fcap. 8vo. 6s.
— Salmonia; or, Days of Fly Fishing. With some Account of the Habits of Fishes belonging to the genus Salmo. Fourth Edition. Woodcuts. Fcap. 8vo. 6s.
DELEPIERRE'S (OCTAVE) History of Flemish Literature and its celebrated Authors. From the Twelfth Century to the present Day. 8vo. 9s.
DENNIS' (GEORGE) Cities and Cemeteries of Etruria. Plates. 2 Vols. 8vo. 42s.
DIXON'S (HEPWORTH) Story of the Life of Lord Bacon; from unpublished Papers. Fcap. 8vo. (In the Press.)
DOG-BREAKING; the Most Expeditious, Certain, and Easy Method, whether great excellence or only mediocrity be required. By LIEUT.-COL. HUTCHINSON. Third Edition. Woodcuts. Post 8vo. 9s.
DOMESTIC MODERN COOKERY. Founded on Principles of Economy and Practical Knowledge, and adapted for Private Families. New Edition. Woodcuts. Fcap. 8vo. 5s.
DOUGLAS'S (GENERAL SIR HOWARD) Treatise on the Theory and Practice of Gunnery. Fifth Edition. Plates. 8vo. 21s.
— Treatise on Military Bridges, and the Passages of Rivers in Military Operations. Third Edition. Plates. 8vo. 21s.
— Naval Warfare with Steam. Second Edition. 8vo. 8s. 6d.
— Modern Systems of Fortification, with special reference to the Naval, Littoral, and Internal Defence of England. Plans. 8vo. 12s.
DRAKE'S (SIR FRANCIS) Life, Voyages, and Exploits, by Sea and Land. By JOHN BARROW. Third Edition. Post 8vo. 2s.
DRINKWATER'S (JOHN) History of the Siege of Gibraltar, 1779-1783. With a Description and Account of that Garrison from the Earliest Periods. Post 8vo. 2s.
DU CHAILLU'S (PAUL B.) EQUATORIAL AFRICA, with Accounts of the Manners and Customs of the People, and of the Chase of the Gorilla, the Nest-building Ape, Chimpanzee, Crocodile, &c. Tenth Thousand. Illustrations. 8vo. 21s.
DUDLEY'S (EARL OF) Letters to the late Bishop of LIandaff. Second Edition. Portrait. 8vo. 10s. 6d.
DUFFERIN'S (LORD) Letters from High Latitudes, being some Account of a Yacht Voyage to Iceland, &c., in 1856. Fourth Edition. Woodcuts. Post 8vo. 9s.
DURHAM'S (ADMIRAL SIR PHILIP) Naval Life and Services. By CAPT. ALEXANDER MURRAY. 8vo. 5s. 6d.
DYER'S (THOMAS H.) Life and Letters of John Calvin. Compiled from authentic Sources. Portrait. 8vo. 15s.
— New History of Modern Europe, from the taking of Constantinople by the Turks to the close of the War in the Crimea. Vols.1 & 2. 8vo. 30s.
EASTLAKE'S (SIR CHARLES) Italian Schools of Painting. From the German of KUGLER. Edited, with Notes. Third Edition. Illustrated from the Old Masters. 2 Vols. Post 8vo. 30s.
EASTWICK'S (E. B.) Handbook for Bombay and Madras, with Directions for Travellers, Officers, &c. Map. 2 Vols. Post 8vo. 24s.
EDWARDS' (W. H.) Voyage up the River Amazon, including a Visit to Para. Post 8vo. 2s.
EGERTON'S (Hon. CAPT. FRANCIS) Journal of a Winter's Tour in India; with a Visit to Nepaul. Woodcuts. 2 Vols. Post 8vo. 18s.
ELDON'S (LORD) Public and Private Life, with Selections from his Correspondence and Diaries. By HORACE TWISS. Third Edition. Portrait. 2 Vols. Post 8vo. 21s.
ELIOT'S (HON. W. G. C.) Khans of the Crimea. Being a Narrative of an Embassy from Frederick the Great to the Court of Krim Gerai. Translated from the German. Post 8vo. 6s.
[page] 11 PUBLISHED BY MR. MURRAY.
ELLIS (REV. W.) Visits to Madagascar, including a Journey to the Capital, with notices of Natural History, and Present Civilisation of the People. Fifth Thousand. Map and Woodcuts. 8vo. 16s.
— (MRS.) Education of Character, with Hints on Moral Training. Post 8vo. 7s. 6d.
ELLESMERE'S (LORD) Two Sieges of Vienna by the Turks. Translated from the German. Post 8vo. 2s.
— Second Campaign of Radetzky in Piedmont. The Defence of Temeswar and the Camp of the Ban. From the German. Post 8vo. 6s. 6d.
— Campaign of 1812 in Russia, from the German of General Carl Von Clausewitz. Map. 8vo. 10s. 6d.
— Pilgrimage, and other Poems. Crown 4to. 24s.
— Essays on History, Biography, Geography, and Engineering. 8vo. 12s.
ELPHINSTONE'S (HON. MOUNTSTUART) History of India—the Hindoo and Mahomedan Periods. Fourth Edition. With an Index. Map. 8vo. 18s.
ENGLAND (HISTORY OF) from the Peace of Utrecht to the Peace of Versailles, 1713—83. By LORD MAHON. Library Edition, 7 Vols. 8vo. 93s.; or Popular Edition, 7 Vols. Post 8vo. 35s.
— From the First Invasion by the Romans, down to the 14th year of Queen Victoria's Reign. By MRS. MARKHAM. 118th Edition. Woodcuts. 12mo. 6s.
— Social, Political, and Industrial, in the 19th Century. By W. JOHNSTON. 2 Vols. Post 8vo. 18s.
ENGLISHWOMAN IN AMERICA. Post 8vo. 10s. 6d.
— RUSSIA. Woodcuts. Post 8vo. 10s. 6d.
EOTHEN; or, Traces of Travel brought Home from the East. A New Edition. Post 8vo. 7s. 6d.
ERSKINE'S (ADMIRAL) Journal of a Cruise among the Islands of the Western Pacific, including the Fejees, and others inhabited by the Polynesian Negro Races. Plates. 8vo. 16s.
ESKIMAUX and English Vocabulary, for Travellers in the Arctic Regions. 16mo. 3s. 6d.
ESSAYS FROM "THE TIMES." Being a Selection from the LITERARY PAPERS which have appeared in that Journal. Seventh Thousand. 2 vols. Fcap. 8vo. 8s.
EXETER'S (BISHOP OF) Letters to the late Charles Butler, on the Theological parts of his Book of the Roman Catholic Church; with Remarks on certain Works of Dr. Milner and Dr. Lingard, and on some parts of the Evidence of Dr. Doyle. Second Edition. 8vo. 16s.
FAIRY RING; A Collection of TALES and STORIES. From the German. By J. E. TAYLOR. Illustrated by RICHARD DOYLE. Second Edition. Fcap. 8vo.
FALKNER'S (FRED.) Muck Manual for the Use of Farmers. A Treatise on the Nature and Value of Manures. Second Edition. Fcap. 8vo. 5s.
FAMILY RECEIPT-BOOK. A Collection of a Thousand Valuable and Useful Receipts. Fcap. 8vo. 5s. 6d.
FANCOURT'S (COL.) History of Yucatan, from its Discovery to the Close of the 17th Century. With Map. 8vo. 10s. 6d.
FARRAR'S (REV. A. S.) Science in Theology. Sermons Preached before the University of Oxford. 8vo. 9s.
— (F. W.) Origin of Language, based on Modern Researches. Fcap. 8vo. 5s.
FEATHERSTONHAUGH'S (G. W.) Tour through the Slave States of North America, from the River Potomac to Texas and the Frontiers of Mexico. Plates. 2 Vols. 8vo. 26s.
FELLOWS' (SIR CHARLES) Travels and Researches in Asia Minor, more particularly in the Province of Lydia. New Edition. Plates. Post 8vo. 9s.
FERGUSSON'S (JAMES) Palaces of Nineveh and Persepolis Restored: an Essay on Ancient Assyrian and Persian Architecture. Woodcuts. 8vo. 16s.
— Handbook of Architecture. Being a Concise and Popular Account of the Different Styles prevailing in all Ages and Countries in the World. With a Description of the most remarkable Buildings. Fourth Thousand. With 850 Illustrations. 8vo. 26s.
FERRIER'S (T. P.) Caravan Journeys in Persia, Afghanistan, Herat, Turkistan, and Beloochistan, with Descriptions of Meshed, Balk, and Candahar, &c. Second Edition. Map. 8vo. 21s.
— History of the Afghans. Map. 8vo. 21s.
FEUERBACH'S Remarkable German Crimes and Trials. Translated from the German by Lady DUFF GORDON. 8vo. 12s.
FISHER'S (REV. GEORGE) Elements of Geometry, for the Use of Schools. Fifth Edition. 18mo. 1s. 6d.
— First Principles of Algebra, for the Use of Schools. Fifth Edition. 18mo. 1s. 6d.
FLOWER GARDEN (THE). An Essay. By REV. THOS. JAMES. Reprinted from the "Quarterly Review." Fcap. 8vo. 1s.
FORBES' (C. S.) Iceland; its Volcanoes, Geysers, and Glaciers. Illustrations. Post 8vo. 14s.
FORD'S (RICHARD) Handbook for Spain, Andalusia, Ronda, Valencia, Catalonia, Granada, Gallicia, Arragon, Navarre, &c. Third Edition. 2 Vols. Post 8vo. 30s.
— Gatherings from Spain. Post 8vo. 3s. 6d.
FORSTER'S (JOHN) Arrest of the Five Members by Charles the First. A Chapter of English History re-written. Post 8vo. 12s.
— Debates on the Grand Remonstrance, 1641. With an Introductory Essay on English freedom under the Plantagenet and Tudor Severeigns. Second Edition. Post 8vo. 12s.
— Oliver Cromwell, Daniel De Foe, Sir Richard Steele, Charles Churchill, Samuel Foote. Biographical Essays. Third Edition. Post 8vo. 12s.
FORSYTH'S (WILLIAM) Hortensius, or the Advocate: an Historical Essay on the Office and Duties of an Advocate. Post 8vo. 12s.
— History of Napoleon at St. Helena. From the Letters and Journals of SIR HUDSON LOWE. Portrait and Maps. 3 Vols. 8vo. 45s.
FORTUNE'S (ROBERT) Narrative of Two Visits to the Tea Countries of China, between the years 1843-52, with full Descriptions of the Tea Plant. Third Edition. Woodcuts. 2 Vols. Post 8vo. 18s.
— Chinese, Inland, on the Coast, and at Sea. A Narrative of a Third Visit in 1853-56. Woodcuts. 8vo. 16s.
[page] 13 PUBLISHED BY MR. MURRAY.
FRANCE (HISTORY OF). From the Conquest by the Gauls to the Death of Louis Philippe. By Mrs. MARKHAM. 56th Thousand. Woodcuts. 12mo. 6s.
FRENCH (THE) in Algiers; The Soldier of the Foreign Legion—and the Prisoners of Abd-el-Kadir. Translated by Lady DUFF GORDON. Post 8vo. 2s.
GALTON'S (FRANCIS) Art of Travel; or, Hints on the Shifts and Contrivances available in Wild Countries. Third Edition. Woodcuts. Post 8vo. 7s. 6d.
GEOGRAPHICAL (THE) Journal. Published by the Royal Geographical Society of London. 8vo.
GERMANY (HISTORY OF). From the Invasion by Marius, to the present time. On the plan of Mrs. MARKHAM. Fifteenth Thousand. Woodcuts, 12mo. 6s.
GIBBON'S (EDWARD) History of the Decline and Fall of the Roman Empire. A New Edition. Preceded by his Autobiography. Edited, with Notes, by Dr. WM. SMITH. Maps. 8 Vols. 8vo. 60s.
— (The Student's Gibbon); Being an Epitome of the Decline and Fall, incorporating the Researches of Recent Commentators. By Dr. WM SMITH. Ninth Thousand. Woodcuts. Post 8vo. 7s. 6d.
GIFFARD'S (EDWARD) Deeds of Naval Daring; or, Anecdotes of the British Navy. 2 Vols. Fcap.8vo. 5s.
GISBORNE'S (THOMAS) Essays on Agriculture. Third Edition. Post 8vo. 5s.
GLADSTONE'S (W. E.) Prayers arranged from the Liturgy for Family Use. Second Edition. 12mo. 2s. 6d.
GOLDSMITH'S (OLIVER) Works. A New Edition. Printed from the last editions revised by the Author. Edited by PETER CUNNING-HAM. Vignettes, 4 Vols. 8vo. 30s. (Murray's British Classics.)
GLEIG'S (REV. G. R) Campaigns of the British Army at Washington and New Orleans. Post 8vo. 2s.
— Story of the Battle of Waterloo. Compiled from Public and Authentic Sources. Post 8vo. 3s. 6d.
— Narrative of Sir Robert Sale's Brigade in Affghanistan, with an Account of the Seizure and Defence of Jellalabad, Post 8vo. 2s.
— Life of Robert Lord Clive. Post 8vo. 3s. d.
— Life and Letters of General Sir Thomas Munro. Post 8vo. 3s. 6d.
GORDON'S (SIR ALEX. DUFF) Sketches of German Life, and Scenes from the War of Liberation. From the German. Post 8vo. 3s. 6d.
—(LADY DUFF) Amber-Witch: the most interesting Trial for Witchcraft ever known. From the German. Post 8vo. 2s.
— French in Algiers. 1. The Soldier of the Foreign Legion. 2, The Prisoners of Abd-el-Kadir. From the French. Post 8vo. 2s.
—Remarkable German Crimes and Trials. From the German. 8vo. 12s.
GOUGER'S (HENRY) Personal Narrative of Two Years' Imprisonment in Burmah. Second Edition. Woodcuts. Post 8vo. 12s.
GRANT'S (ASAHEL) Nestorians, or the Lost Tribes; containing Evidence of their Identity, their Manners, Customs, and Ceremonies; with Sketches of Travel in Ancient Assyria, Armenia, and Mesopotamia; and Illustrations of Scripture Prophecy. Third Edition. Fcap 8vo, 6s.
GRENVILLE (THE) PAPERS. Being the Public and Private Correspondence of George Grenville, his Friends and Contemporaries, during a period of 30 years.—Including his DIARY OF POLITICAL EVENTS while First Lord of the Treasury. Edited, with Notes, by W. J. SMITH. 4 Vols. 8vo. 16s. each.
GREEK GRAMMAR FOR SCHOOLS. Abridged from Matthiæ. By the BISHOP OF LONDON. Ninth Edition, revised by Rev. J. EDWARDS. 12mo. 3s.
GREY'S (SIR GEORGE) Polynesian Mythology, and Ancient Traditional History of the New Zealand Race. Woodcuts. Post 8vo. 10s. 6d.
GROTE'S (GEORGE) History of Greece. From the Earliest Times to the close of the generation contemporary with the death of Alexander the Great. Third Edition. Portrait and Maps. 12 vols. 8vo. 16s. each.
— Plato's Doctrine on the Rotation of the Earth, and Aristotle's Comment upon that Doctrine. Second Edition. 8vo. 1s. 6d.
— (MRS.) Memoir of the Life of the late Ary Scheffer. Second Edition. Portrait. Post 8vo. 8s. 6d.
GUSTAVUS VASA (History of), King of Sweden. With Extracts from his Correspondence. Portrait. 8vo. 10s. 6d.
HALLAM'S (HENRY) Constitutional History of England, from the Accession of Henry the Seventh to the Death of George the Second. Seventh Edition. 3 Vols. 8vo. 30s.
— History of Europe during the Middle Ages. Tenth Edition. 3 Vols. 8vo. 30s.
— Introduction to the Literary History of Europe, during the 16th, 17th, and 18th Centuries. Fourth Edition. 3 Vols. 8vo. 36s.
— Literary Essays and Characters. Selected from the last work. Fcap. 8vo. 2s.
— Historical Works. Complete. Containing—History of England,—Middle Ages of Europe,—and Literary History of Europe. Cabinet Edition. 10 Vols. Post 8vo. 6s. each.
HAMILTON'S (JAMES) Wanderings in Northern Africa, Benghazi, Cyrene, the Oasis of Siwah, &c. Second Edition. Woodcuts. Post 8vo. 12s.
HAMPDEN'S (BISHOP) Philosophical Evidence of Christianity, or the Credibility obtained to a Scripture Revelation from its Coincidence with the Facts of Nature. 8vo. 9s. 6d.
HARCOURT'S (EDWARD VERNON) Sketch of Madeira; with Map and Plates. Post 8vo. 8s. 6d.
HART'S ARMY LIST. (Quarterly and Annually.) 8vo. 10s. 6d. and 21s.
HAY'S (J. H. DRUMMOND) Western Barbary, its wild Tribes and savage Animals. Post 8vo. 2s.
HEBER'S (BISHOP) Journey through the Upper Provinces of India, From Calcutta to Bombay, with a Journey to Madras and the Southern Provinces. Twelfth Edition. 2 Vols. Post 8vo. 7s.
— Poetical Works. Sixth Edition. Portrait. Fcap. 8vo. 6s.
— Parish Sermons; on the Lessons, the Gospel, or the Epistle, for every Sunday in the Year, and for Week-day Festivals. Sixth Edition. 2 Vols. Post 8vo. 16s.
— Sermons Preached in England. Second Edition. 8vo. 9s. 6d.
— Hymns written and adapted for the Weekly Church Service of the Year. Twelfth Edition. 16mo. 2s.
[page] 15 PUBLISHED BY MR. MURRAY.
HAND-BOOK—TRAVEL-TALK. In English, German, French, and Italian. 18mo. 3s. 6d.
— NORTH GERMANY, HOLLAND, BELGIUM, and the Rhine to Switzerland. Map. Post 8vo. 10s.
— SOUTH GERMANY, Bavaria, Austria, Salzberg, the Austrian and Bavarian Alps, the Tyrol, and the Danube, from Ulm to the Black Sea. Map. Post 8vo. 10s.
— PAINTING. The German, Flemish, and Dutch Schools. Edited by DR. WAAGEN. Woodcuts. 2 Vols. Post 8vo. 24s.
— SWITZERLAND, Alps of Savoy, and Piedmont. Maps. Post 8vo. 9s.
— FRANCE, Normandy, Brittany, the French Alps, the Rivers Loire, Seine, Rhone, and Garonne, Dauphiné, Provence, and the Pyrenees. Maps. Post 8vo. 10s.
— PARIS AND ITS ENVIRONS. Map. Post 8vo. (Nearly Ready.)
— SPAIN, Andalusia, Ronda, Granada, Valencia, Catalonia, Gallicia, Arragon, and Navarre. Maps. 2 Vols. Post 8vo. 30s.
— PORTUGAL, LISBON, &c. Map. Post 8vo. 9s.
— NORTH ITALY, Sardinia, Genoa, the Riviera, Venice, Lombardy, and Tuscany. Map. Post 8vo. 12s.
— CENTRAL ITALY, FLORENCE, SOUTH TUSCANY, and the PAPAL STATES. Map. Post 8vo. 10s.
— ROME AND ITS ENVIRONS. Map. Post 8vo. 9s.
— SOUTH ITALY, Naples, Pompeii, Herculaneum, Vesuvius, &c. Map. Post 8vo. 10s.
— SICILY. Map. Post 8vo. (In the Press.)
— PAINTING. The Italian Schools. From the German of KUGLER. Edited by Sir CHARLES EASTLAKE, R.A. Woodcuts. 2 Vols. Post 8vo. 30s.
— LIVES OF THE EARLY ITALIAN PAINTERS, CIMABUE to BASSANO, AND PROGRESS OF PAINTING IN ITALY. By Mrs. JAMESON. Woodcuts. Post 8vo. 12s.
— DICTIONARY OF ITALIAN PAINTERS. By A LADY. Edited by RALPH WORNUM. With a Chart. Post 8vo. 6s. 6d.
— GREECE, Ionian Islands, Albania, Thessaly, and Macedonia. Maps. Post 8vo. 15s.
— TURKEY, MALTA, ASIA MINOR, CONSTANTINOPLE, Armenia, Mesopotamia, &c. Maps. Post 8vo. (In the Press.)
— EGYPT, Thebes, the Nile, Alexandria, Cairo, the Pyramids, Mount Sinai, &c. Map. Post 8vo. 15s.
— SYRIA & PALESTINE. Maps. 2 Vols. Post 8vo. 24s.
— BOMBAY AND MADRAS. Map. 2 Vols. Post 8vo. 24s.
— DENMARK, NORWAY and SWEDEN. Maps. Post 8vo. 15s.
HAND-BOOK—RUSSIA, THE BALTIC AND FINLAND. Maps. Post 8vo. 12s.
— KENT AND SUSSEX. Map. Post 8vo. 10s.
— SURREY, HANTS, and ISLE OF WIGHT. Maps. Post 8vo. 7s. 6d.
— MODERN LONDON. A Complete Guide to the Metropolis. Map. 16mo. 5s.
— LONDON, PAST AND PRESENT. Second Edition. Post 8vo. 16s.
— WESTMINSTER ABBEY. Woodcuts. 16mo. 1s.
— ENVIRONS OF LONDON. Maps. Post 8vo. (In preparation.)
— BERKS, BUCKS, AND OXFORDSHIRE. Map. Post 8vo. 7s. 6d.
— WILTS, DORSET, AND SOMERSET. Map. Post 8vo. 7s. 6d.
— DEVON AND CORNWALL. Maps. Post 8vo. 7s. 6d.
— SOUTHERN CATHEDRALS OF ENGLAND. Winchester, Salisbury, Exeter, Wells, Chichester, Rochester, Canterbury. With 150 Illustrations. 2 Vols. Post 8vo. 24s.
— NORTH AND SOUTH WALES. Maps. 2 Vols. Post 8vo. 12s.
— FAMILIAR QUOTATIONS. From English Authors. Third Edition. Fcap. 8vo. 5s.
— ARCHITECTURE. In all Ages and Countries. By JAMES FERGUSSON. Fourth Thousand. With 850 Illustrations. 8vo. 26s.
— ARTS OF THE MIDDLE AGES. By M. JULES LABARTE. With 200 Illustrations. 8vo. 18s.
HEAD'S (SIR FRANCIS) Horse and his Rider. Fourth Thousand. Woodcuts. Post 8vo. 5s.
— Rapid Journeys across the Pampas and over the Andes. Post 8vo. 2s.
— Descriptive Essays. 2 Vols. Post 8vo. 18s.
— Bubbles from the Brunnen of Nassau. By an OLD MAN. Sixth Edition. 16mo. 5s.
— Emigrant. Sixth Edition. Fcap. 8vo. 2s. 6d.
— Stokers and Pokers; or, the North-Western. Railway. Post 8vo. 2s.
— Defenceless State of Great Britain. Post 8vo. 12s.
— Faggot of French Sticks; or, Sketches of Paris. New Edition. 2 Vols. Post 8vo. 12s.
— Fortnight in Ireland. Second Edition. Map. 8vo. 12s.
— (SIR GEORGE) Forest Scenes and Incidents in Canada. Second Edition. Post 8vo. 10s.
— Home Tour through the Manufacturing Districts of England. Third Edition. 2 Vols. Post 8vo. 12s.
— (SIR EDMUND) Shall and Will; or, Two Chapters on Future Auxiliary Verbs. Second Edition, Enlarged. Fcap. 8vo. 4s.
[page] 17 PUBLISHED BY MR. MURRAY.
HEIRESS (THE) in Her Minority; or, The Progress of Character. By the Author of "BERTHA'S JOURNAL," 2 Vols. 12mo. 18s.
HERODOTUS. A New English Version. Edited with Notes and Essays, historical, ethnographical, and geographical. By Rev. G. RAWLINSON, assisted by SIR HENRY RAWLINSON and SIR J. G. WILKINSON. Second Edition. Maps and Woodcuts. 4 Vols. 8vo. 48s.
HERVEY'S (LORD) Memoirs of the Reign of George the Second, from his Accession to the Death of Queen Caroline. Edited, with Notes, by MR. CROKER. Second Edition. Portrait. 2 Vols. 8vo. 21s.
HESSEY (REV. DR.). Sunday—Its Origin, History, and Present Obligations. Being the Bampton Lectures for 1860. Second Edition. 8vo. 16s.
HICKMAN'S (WM.) Treatise on the Law and Practice of Naval Courts-Martial. 8vo. 10s. 6d.
HILLARD'S (G. S.) Six Months in Italy. 2 Vols. Post 8vo. 16s.
HOLLWAY'S (J. G.) Month in Norway. Fcap. 8vo. 2s.
HONEY BEE (THE). An Essay. By REV. THOMAS JAMES. Reprinted from the "Quarterly Review." Fcap. 8vo. 1s.
HOOK'S (DEAN) Church Dictionary. Eighth Edition. 8vo. 16s.
— Discourses on the Religious Controversies of the Day. 8vo. 9s.
— (THEODORE) Life. By J . G. LOCKHART. Reprinted from the "Quarterly Review." Fcap. 8vo. 1s.
HOOKER'S (Dr.J. D.) Himalayan Journals; or, Notes of an Oriental Naturalist in Bengal, the Sikkim and Nepal Himalayas, the Khasia Mountains, &c. Second Edition. Woodcuts. 2 Vols. Post 8vo. 18s.
HOOPER'S (LIEUT.) Ten Months among the Tents of the Tuski; with Incidents of an Arctic Boat Expedition in Search of Sir John Franklin. Plates. 8vo. 14s.
HOPE'S (A. J. BERESFORD) English Cathedral of the Nineteenth Century. With Illustrations. 8vo. 12s.
HORACE (Works of). Edited by DEAN MILMAN. With 300 Woodcuts. Crown 8vo. 21s.
— (Life of). By DEAN MILMAN. Woodcuts, and coloured Borders. 8vo. 9s.
HOSPITALS AND SISTERHOODS. By A LADY. Fcap. 8vo. 3s. 6d.
HUME'S (DAVID) History of England, from the Invasion of Julius Cæsar to the Revolution of 1688. Abridged for Students. Correcting his errors, and continued to 1858. Twentieth Thousand. Woodcuts. Post 8vo. 7s. 6d.
HUTCHINSON (COL.) on the most expeditious, certain, and easy Method of Dog-Breaking. Third Edition. Woodcuts. Post 8vo. 9s.
HUTTON'S (H. E.) Principia Græca; an Introduction to the Study of Greek. Comprehending Grammar, Delectus, and Exercise-book, with Vocabularies. Second Edition, 12mo. 3s.
C
HOME AND COLONIAL LIBRARY. A Series of Works adapted for all circles and classes of Readers, having been selected for their acknowledged interest and ability of the Authors. Post 8vo. Published at 2s. and 3s. 6d. each, and arranged under two distinctive heads as follows:—
CLASS A.
HISTORY, BIOGRAPHY, AND HISTORIC TALES.
1. SIEGE OF GIBRALTAR. By JOHN DRINKWATER. 2s.
2. THE AMBER-WITCH. By LADY DUFF GORDON. 2s.
3. CROMWELL AND BUNYAN. By ROBERT SOUTHEY. 2s.
4. LIFE OF SIR FRANCIS DRAKE. By JOHN BARROW.
5. CAMPAIGNS AT WASHINGTON. By REV. G. R. GLEIG. 2s.
6. THE FRENCH IN ALGIERS. By LADY DUFF GORDON. 2s.
7. THE FALL OF THE JESUITS. 2s.
8. LIVONIAN TALES. 2s.
9. LIFE OF CONDE. By LORD MAHON. 3s. 6d.
10. SALE'S BRIGADE. By REV. G. R. GLEIG. 2s.
11. THE SIEGES OF VIENNA. By LORD ELLESMERE. 2s.
12. THE WAYSIDE CROSS. By CAPT. MILMAN. 2s.
13. SKETCHES OF GERMAN LIFE. By SIR A. GORDON. 3s. 6d.
14. THE BATTLE OF WATERLOO. By REV. G. R. GLEIG. 3s. 6d.
15. AUTOBIOGRAPHY OF STEFFENS. 2s.
16. THE BRITISH POETS. By THOMAS CAMPBELL. 3s. 6d.
17. HISTORICAL ESSAYS. By. LORD MAHON. 3s. 6d.
18. LIFE OF LORD CLIVE. By REV. G. R. GLEIG. 3s. 6d.
19. NORTH - WESTERN RAILWAY. By SIR F. B. HEAD. 2s
20. LIFE OF MUNRO. By REV. G. R. GLEIG. 3s. 6d.
CLASS B.
VOYAGES, TRAVELS, AND ADVENTURES.
1. BIBLE IN SPAIN. By GEORGE BORROW. 3s. 6d.
2. GIPSIES OF SPAIN. By GEORGE BORROW. 3s. 6d.
3&4. JOURNALS IN INDIA. By BISHOP HEBER. 2 Vols. 7s.
5. TRAVELS IN THE HOLY LAND. By IRBY and MANGLES. 2s.
6. MOROCCO AND THE MOORS. By J. DRUMMOND HAY. 2s.
7. LETTERS FROM THE BALTIC. By a LADY. 2s.
8. NEW SOUTH WALES. By MRS. MEREDITH. 2s.
9. THE WEST INDIES. By M. G. LEWIS. 2s.
10. SKETCHES OF PERSIA. By SIR JOHN MALCOLM. 3s. 6d.
11. MEMOIRS OF FATHER RIPA. 2s.
12 & 13. TYPEE AND OMOO. By HERMANN MELVILLE. 2 Vols. 7s.
14. MISSIONARY LIFE IN CANADA. By REV. J. ABBOTT. 2s.
15. LETTERS FROM MADRAS. By a LADY. 2s.
16. HIGHLAND SPORTS. By CHARLES ST. JOHN. 3s. 6d.
17. PAMPAS JOURNEYS. By SIR F. B. HEAD. 2s.
18. GATHERINGS FROM SPAIN. By RICHARD FORD. 3s. 6d.
19. THE RIVER AMAZON. By W. H. EDWARDS. 2s.
20. MANNERS & CUSTOMS OF INDIA. By REV. C. ACLAND. 2s.
21. ADVENTURES IN MEXICO. By G. F. RUXTON. 3s. 6d.
22. PORTUGAL AND GALLICIA. By LORD CARNARVON. 3s. 6d.
23. BUSH LIFE IN AUSTRALIA. By Rev. H. W. HAYGARTH. 2s.
24. THE LIBYAN DESERT. By BAYLE ST. JOHN. 2s.
25. SIERRA LEONE. By a LADY. 3s. 6d.
*** Each work may be had separately.
[page] 19 PUBLISHED BY MR. MURRAY.
IRBY AND MANGLES' Travels in Egypt, Nubia, Syria, and the Holy Land. Post 8vo. 2s.
JAMES' (REV. THOMAS) Fables of Æsop. A New Translation, with Historical Preface. With 100 Woodcuts by TENNIEL and WOLF. Thirty-eighth Thousand. Post 8vo. 2s. 6d.
JAMESON'S (MRS.) Lives of the Early Italian Painters, from Cimabue to Bassano, and the Progress of Painting in Italy. New Edition. With Woodcuts. Post 8vo. 12s.
JERVIS'S (CAPT.) Manual of Operations in the Field. Post 8vo. 9s. 6d.
JESSE'S (EDWARD) Visits to Spots of Interest in the Vicinity of Windsor and Eton. Woodcuts. Post 8vo. 12s.
— Scenes and Occupations of Country Life. Third Edition. Woodcuts. Fcap. 8vo. 6s.
— Gleanings in Natural History. Eighth Edition. Fcap. 8vo. 6s.
JOHNSON'S (DR. SAMUEL) Life. By James Boswell. Including the Tour to the Hebrides. Edited by the late MR. CROKER. Portraits. Royal 8vo. 10s.
— Lives of the most eminent English Poets. Edited by PETER CUNNINGHAM. 3 vols. 8vo. 22s. 6d. (Murray's British Classics.)
JOHNSTON'S (WM.) England: Social, Political, and Industrial, in 19th Century. 2 Vols. Post 8vo. 18s.
JOURNAL OF A NATURALIST. Fourth Edition. Woodcuts. Post 8vo. 9s. 6d.
JOWETT (Rev. B.) on St. Paul's Epistles to the Thessalonians, Galatians, and Romans. Second Edition. 2 Vols. 8vo. 30s.
JONES' (Rev. R.) Literary Remains. With a Prefatory Notice. By Rev. W. WHEWELL, D.D. Portrait. 8vo. 14s.
KEN'S (BISHOP) Life. By A LAYMAN. Second Edition. Portrait. 2 Vols. 8vo. 18s.
— Exposition of the Apostles' Creed. Extracted from his "Practice of Divine Love." New Edition. Fcap. 1s. 6d.
— Approach to the Holy Altar. Extracted from his "Manual of Prayer" and "Practice of Divine Love." New Edition. Fcap. 8vo. 1s. 6d.
KING'S (REV. S. W.) Italian Valleys of the Alps; a Tour through all the Romantic and less-frequented "Vals" of Northern Piedmont. Illustrations. Crown 8vo. 18s.
— (REV. C. W.) Antique Gems; their Origin, Use, and Value, as Interpreters of Ancient History, and as illustrative of Ancient Art. Illustrations. 8vo. 42s.
KING EDWARD VITH's Latin Grammar; or, an Introduction to the Latin Tongue, for the Use of Schools. Fifteenth Edition. 12mo. 3s. 6d.
— First Latin Book; or, the Accidence, Syntax, and Prosody, with an English Translation for the Use of Junior Classes. Fourth Edition. 12mo. 2s. 6d.
C 2
KINGLAKE'S (A. W.) History of the War in the Crimea. Based chiefly upon the Private Papers of Field Marshal Lord Raglan, and other authentic materials. Vols. I.—III. 8vo. (In Preparation.)
KNAPP'S (J. A.) English Roots and Ramifications; or, the Derivation and Meaning of Divers Words. Fcap. 8vo. 4s.
KUGLER'S Italian Schools of Painting. Edited, with Notes, by SIR CHARLES EASTLAKE. Third Edition. Woodcuts. 2 Vols. Post 8vo. 30s.
— German, Dutch, and Flemish Schools of Painting. Edited, with Notes, by DR. WAAGEN. Second Edition. Woodcuts. 2 Vols. Post 8vo. 24s.
LABARTE'S (M. JULES) Handbook of the Arts of the Middle Ages and Renaissance. With 200 Woodcuts. 8vo. 18s.
LABORDE'S (LEON DE) Journey through Arabia Petræa, to Mount Sinai, and the Excavated City of Petræa,—the Edom of the Prophecies. Second Edition. With Plates. 8vo. 18s.
LANE'S (E. W.) Manners and Customs of the Modern Egyptians. A New Edition, with Additions and Improvements by the Author. Edited by E. STANLEY POOLE. Woodcuts. 8vo. 18s.
LATIN GRAMMAR (KING EDWARD VITH'S). For the Use of Schools. Fifteenth Edition. 12mo. 3s. 6d.
— First Book (KING EDWARD VITH's); or, the Accidence, Syntax, and Prosody, with English Translation for Junior Classes. Fourth Edition. 12mo. 2s. 6d.
LAYARD'S (A. H.) Nineveh and its Remains. Being a Natrative of Researches and Discoveries amidst the Ruins of Assyria. With an Account of the Chaldean Christians of Kurdistan; the Yezedis, or Devil-worshippers; and an Enquiry into the Manners and Arts of the Ancient Assyrians. Sixth Edition. Plates and Woodcuts. 2 Vols. 8vo. 36s.
— Nineveh and Babylon; being the Result of a Second Expedition to Assyria. Fourteenth Thousand. Plates. 8vo. 21s. Or Fine Paper, 2 Vols. 8vo. 30s.
— Popular Account of Nineveh. 15th Edition. With Woodcuts. Post 8vo. 5s.
LESLIE'S (C. R.) Handbook for Young Painters. With Illustrations. Post 8vo. 10s. 6d.
— Autobiographical Recollections, with Selections from his Correspondence. Edited by TOM TAYLOR. Portrait. 2 Vols. Post 8vo. 18s.
— Life of Sir Joshua Reynolds. With an Account of his Works, and a Sketch of his Cotemporaries. Fcap. 4to. (In the Press.)
LEAKE'S (COL.) Topography of Athens, with Remarks on its Antiquities. Second Edition. Plates. 2 Vols. 8vo. 30s.
— Travels in Northern Greece. Maps. 4 Vols. 8vo. 60s.
— Disputed Questions of Ancient Geography. Map. 8vo. 6s. 6d.
— Numismata Hellenica, and Supplement. Completing a descriptive Catalogue of Twelve Thousand Greek Coins, with Notes Geographical and Historical. With Map and Appendix. 4to. 63s.
— Peloponnesiaca. 8vo. 15s.
— On the Degradation of Science in England. 8vo. 3s. 6d.
[page] 21 PUBLISHED BY MR. MURRAY.
LETTERS FROM THE SHORES OF THE BALTIC. By a LADY. Post 8vo. 2s.
— Madras; or, Life and Manners in India. By a LADY. ost 8vo. 2s.
— Sierra Leone, written to Friends at Home. By a LADY. Edited by Mrs. NORTON. Post 8vo. 3s. 6d.
— Head Quarters; or, The Realities of the War in the Crimea. By a STAFF OFFICER. Plans. Post 8vo. 6s.
LEXINGTON (THE) PAPERS; or, Some Account of the Courts of London and Vienna at the end of the 17th Century. Edited by HON. H. MANNERS SUTTON. 8vo. 14s.
LEWIS' (SIR G. C.) Essay on the Government of Dependencies. 8vo. 12s.
— Glossary of Provincial Words used in Herefordshire and some of the adjoining Counties. 12mo. 4s. 6d.
— (LADY THERESA) Friends and Contemporaries of the Lord Chancellor Clarendon, illustrative of Portraits in his Gallery. With a Descriptive Account of the Pictures, and Origin of the Collection. Portraits. 3 Vols. 8vo. 42s.
— (M. G.) Journal of a Residence among the Negroes in the West Indies. Post 8vo. 2s.
LIDDELL'S (DEAN) History of Rome. From the Earliest Times to the Establishment of the Empire. With the History of Literature and Art. 2 Vols. 8vo. 28s.
— Student's History of Rome. Abridged from the Larger Work. Eighteenth Thousand. With 100 Woodcuts. Post 8vo. 7s. 6d.
LINDSAY'S (LORD) Lives of the Lindsays; or, a Memoir of the Houses of Crawfurd and Balcarres. With Extracts from Official Papers and Personal Narratives. Second Edition. 3 Vols. 8vo. 24s.
— Report of the Claim of James, Earl of Crawfurd and Balcarres, to the Original Dukedom of Montrose, created in 1488. Folio. 15s.
— Scepticism; a Retrogressive Movement in Theology and Philosophy. 8vo. 9s.
LITTLE ARTHUR'S HISTORY OF ENGLAND. By LADY CALLCOTT. 120th Thousand. With 20 Woodcuts. Fcap. 8vo. 2s. 6d.
LIVINGSTONE'S (REV. DR.) Missionary Travels and Researches in South Africa; including a Sketch of Sixteen Years' Residence in the Interior of Africa, and a Journey from the Cape of Good Hope to Loanda on the West Coast; thence across the Continent, down the River Zambesi, to the Eastern Ocean. Thirtieth Thousand. Map, Plates, and Index. 8vo. 21s.
— Popular Account of Travels in South Africa. Condensed from the above. Map and Illustrations. Post 8vo. 6s.
LIVONIAN TALES. By the Author of "Letters from the Baltic." Post 8vo. 2s.
LOCKHART'S (J. G.) Ancient Spanish Ballads. Historical and Romantic. Translated, with Notes. Illustrated Edition. 4to. 21s. Or, Popular Edition, Post 8vo. 2s. 6d.
— Life of Robert Burns. Fifth Edition. Fcap. 8vo. 3s.
LONDON (BISHOP OF). The Dangers and Safeguards of Modern Theology. Containing Suggestions to the Theological Student under present difficulties. 8vo. 9s.
LOUDON'S (MRS.) Instructions in Gardening for Ladies. With Directions and Calendar of Operations for Every Month. Eighth Edition. Woodcuts. Fcap. 8vo. 5s.
— Modern Botany; a Popular Introduction to the Natural System of Plants. Second Edition. Woodcuts. Fcap. 8vo. 6s.
LOWE'S (SIR HUDSON) Letters and Journals, during the Captivity of Napoleon at St. Helena. By WILLIAM FORSYTH. Portrait. 3 Vols. 8vo. 45s.
LUCKNOW: A Lady's Diary of the Siege. Fourth Thousand. Fcap. 8vo. 4s. 6d.
LYELL'S (SIR CHARLES) Principles of Geology; or, the Modern Changes of the Earth and its Inhabitants considered as illustrative of Geology. Ninth Edition. Woodcuts. 8vo. 18s.
— Visits to the United States, 1841-46. Second Edition. Plates. 4 Vols. Post 8vo. 24s.
MAHON'S (LORD) History of England, from the Peace of Utrecht to the Peace of Versailles, 1713—83. Library Edition, 7 Vols. 8vo. 93s. Popular Edition, 7 Vols. Post 8vo. 35s.
— Life of William Pitt, with Extracts from his MS. Papers. Portrait. Vols. I. and II. Post 8vo. 21s.
— "Forty-Five;" a Narrative of the Rebellion in Scotland. Post 8vo. 3s.
— History of British India from its Origin till the Peace of 1783. Post 8vo. 3s. 6d.
— History of the War of the Succession in Spain. Second Edition. Map. 8vo. 15s.
— Spain under Charles the Second; or, Extracts from the Correspondence of the Hon. ALEXANDER STANHOPE, British Minister at Madrid from 1690 to 1700. Second Edition. Post 8vo. 6s. 6d.
— Life of Louis, Prince of Condé, surnamed the Great. Post 8vo. 3s. 6d.
— Life of Belisarius. Second Edition. Post 8vo. 10s. 6d.
— Historical and Critical Essays. Post 8vo. 3s. 6d.
— Story of Joan of Arc. Fcap. 8vo. 1s.
— Addresses Delivered at Manchester, Leeds, and Birmingham. Fcap. 8vo. 1s.
McCLINTOCK'S (CAPT. SIR F. L.) Narrative of the Discovery of the Fate of Sir John Franklin and his Companions in the Arctic Seas. Twelfth Thousand. Illustrations. 8vo. 16s.
McCOSH (REV. DR.) on the Intuitive Convictions of the Mind inductively investigated. 8vo. 12s.
McCULLOCH'S (J. R.) Collected Edition of RICARDO'S Political Works. With Notes and Memoir. Second Edition. 8vo. 16s.
MAINE (H. SUMNER) on Ancient Law: its Connection with the Early History of Society, and its Relation to Modern Ideas. 8vo. 12s.
[page] 23 PUBLISHED BY MR. MURRAY.
MALCOLM'S (SIR JOHN) Sketches of Persia. Third Edition. Post 8vo. 3s. 6d.
MANSEL (REV. H. L.) Limits of Religious Thought Examined. Being the Bampton Lectures for 1858. Fourth Edition. Post 8vo. 7s. 6d.
MANTELL'S (GIDEON A.) Thoughts on Animalcules; or, the Invisible World, as revealed by the Microscope. Second Edition. Plates. 16mo. 6s.
MANUAL OF SCIENTIFIC ENQUIRY, Prepared for the Use of Officers and Travellers. By various Writers. Third Edition revised by the Rev. R. MAIN. Maps. Post 8vo. 9s. (Published by order of the Lords of the Admiralty.)
MARKHAM'S (MRS.) History of England. From the First Invasion by the Romans, down to the fourteenth year of Queen Victoria's Reign. 118th Edition. Woodcuts. 12mo. 6s.
— History of France. From the Conquest by the Gauls, to the Death of Louis Philippe. Sixtieth Edition. Woodcuts. 12mo. 6s.
— History of Germany. From the Invasion by Marius, to the present time. Fifteenth Edition. Woodcuts. 12mo. 6s.
— A Smaller History of Greece. From the Earliest Times to the Roman Conquest. By Dr. WM. SMITH. Woodcuts. 16mo. 3s. 6d.
— A Smaller History of Rome, from the Earliest Times to the Establishment of the Empire. By DR. WM. SMITH. Woodcuts. 16mo. 3s. 6d.
MARKLAND'S (J. H.) Reverence due to Holy Places. Third Edition. Fcap. 8vo. 2s.
MARRYAT'S (JOSEPH) History of Modern and Mediæval Pottery and Porcelain. With a Description of the Manufacture. Second Edition. Plates and Woodcuts. 8vo. 31s. 6d.
— (HORACE) Residence in Jutland, the Danish Isles, and Copenhagen. Illustrations. 2 Vols. Post 8vo. 24s.
— One Year in Sweden, including a Visit to the Isle of Gothland. Illustrations. 2 Vols. Post 8vo. 24s.
MATTHIÆ'S (AUGUSTUS) Greek Grammar for Schools. Abridged from the Larger Grammar. By Blomfield. Ninth Edition. Revised by EDWARDS. 12mo. 3s.
MAUREL'S (JULES) Essay on the Character, Actions, and Writings of the Duke of Wellington. Second Edition. Fcap. 8vo. 1s. 6d.
MAWE'S (H. L.) Journal of a Passage from the Pacific to the Atlantic. 8vo. 12s.
MAXIMS AND HINTS on Angling and Chess. To which is added the Miseries of Fishing. By RICHARD PENN. New Edition. Woodcuts. 12mo. 1s.
MAYO'S (DR.) Pathology of the Human Mind. Fcap. 8vo. 5s. 6d.
MELVILLE'S (HERMANN) Typee and Omoo; or, Adventures amongst the Marquesas and South Sea Islands. 2 Vols. Post 8vo. 7s.
MENDELSSOHN'S Life. By JULES BENEDICT. 8vo. 2s. 6d.
MEREDITH'S (MRS. CHARLES) Notes and Sketches of New South Wales, during a Residence from 1839 to 1844. Post 8vo. 2s.
— Tasmania, during a Residence of Nine Years. With Illustrations. 2 Vols. Post 8vo. 18s.
MERRIFIELD (MRS.) on the Arts of Painting in Oil, Miniature, Mosaic, and Glass; Gilding, Dyeing, and the Preparation of Colours and Artificial Gems, described in several old Manuscripts. 2 Vols. 8vo. 30s.
MESSIAH (THE). Map. 8vo. 18s.
MILLS' (ARTHUR) India in 1858; A Summary of the Existing Administration—Political, Fiscal, and Judicial; with Laws and Public Documents, from the earliest to the present time. Second Edition. With Coloured Revenue Map. 8vo. 10s. 6d.
MITCHELL'S (THOMAS) Plays of Aristophanes. With English Notes. 8vo.—1. CLOUDS, 10s.—2. WASPS, 10s.—3. FROGS, 15s.
MILMAN'S (DEAN) History of Latin Christianity; including that of the Popes to the Pontificate of Nicholas V. Second Edition. 6 Vols. 8vo. 72s.
— Character and Conduct of the Apostles considered as an Evidence of Christianity. 8vo. 10s. 6d.
— Life and Works of Horace. With 300 Woodcuts. New Edition. 2 Vols. Crown 8vo. 30s.
— Poetical Works. Plates. 3 Vols. Fcap. 8vo. 18s.
— Fall of Jerusalem. Fcap. 8vo. 1s.
— (CAPT. E. A.) Wayside Cross; or, the Raid of Gomez. A Tale of the Carlist War. Post 8vo. 2s.
MODERN DOMESTIC COOKERY. Founded on Principles of Economy and Practical Knowledge, and adapted for Private Families. New Edition. Woodcuts. Fcap. 8vo. 5s.
MOLTKE'S (BARON) Russian Campaigns on the Danube and the Passage of the Balkan, 1828-9. Plans. 8vo. 14s.
MONASTERY AND THE MOUNTAIN CHURCH. By Author of "Sunlight through the Mist." Woodcuts. 16mo. 4s..
MOORE'S (THOMAS) Life and Letters of Lord Byron. Cabinet Edition. Plates. 6 Vols. Fcap. 8vo. 18s.
— Life and Letters of Lord Byron. Portraits. Royal 8vo. 9s.
MOTLEY'S (J. L.) History of the United Netherlands: from the Death of William the Silent to the Synod of Dort. Embracing the English-Dutch struggle against Spain; and a detailed Account of the Spanish Armada. Fourth Thousand. Portraits. 2 Vols. 8vo. 30s.
MOZLEY'S (REV. J. B.) Treatise on the Augustinian Doctrine of Predestination. 8vo. 14s.
— Primitive Doctrine of Baptismal Regeneration. 8vo. 7s. 6d.
MUCK MANUAL (The) for the Use of Farmers. A Practical Treatise on the Chemical Properties, Management, and Application of Manures. By FREDERICK FALENER. Second Edition. Fcap.8vo. 5s.
MUNDY'S (GEN.) Pen and Pencil Sketches during a Tour in India. Third Edition. Plates. Post 8vo. 7s. 6d.
MUNRO'S (GENERAL SIR THOMAS) Life and Letters. By the REV. G. R. GLEIG. Post 8vo. 3s. 6d.
MURCHISON'S (SIR RODERICK) Russia in Europe and the Ural Mountains; Geologically Illustrated. With Coloured Maps, Plates, Sections, &c. 2 Vols. Royal 4to
— Siluria; or, a History of the Oldest Rocks containing Organic Remains. Third Edition. Map and Plates. 8vo. 42s.
[page] 25 PUBLISHED BY MR. MURRAY.
MURRAY'S RAILWAY READING. For all classes of Readers.
[The following are published:]
WELLINGTON. By LORD ELLESMERE. 6d.
NIMBOD ON THE CHASE, 1s.
ESSAYS FROM "THE TIMES." 2 Vols. 8s.
MUSIC AND DRESS. 1s.
LATARD'S ACCOUNT OF NINEVEH. 5s.
MILMAN'S FALL OF JERUSALEM. 1s.
MAHON'S "FORTY-FIVE" 3s.
LIFE OF THEODORE HOOK. 1s.
DEEDS OF NAVAL DARING. 2 Vols. 5s.
THE HONEY BEE. 1s.
JAMES' ÆSOP'S FABLES. 2s. 6d.
NIMBOD ON THE TURF. 1s. 6d.
OLIPHANT'S NEPAUL. 2s. 6d.
ART OF DINING. 1s. 6d.
HALLAM'S LITERARY ESSAYS. 2s.
MAHON'S JOAN OF ARC. 1s.
HEAD'S EMIGRANT. 2s. 6d.
NIMBOD ON THE ROAD. 1s.
WILKINSON'S ANCIENT EGYPTIANS. 12s.
CROKER ON THE GUILLOTINE. 1s.
HOLLWAY'S NORWAY. 2s.
MAURRL'S WELLINGTON. 1s. 6d.
CAMPBELL'S LIFE OF BACON. 2s. 6d.
THE FLOWER GARDEN. 1s.
LOCKHART'S SPANISH BALLADS. 2s. 6d.
LUCAS ON HISTORY. 6d.
BEAUTIES OF BYRON. 3s.
TAYLOR'S NOTKS. FROM LIFE. 2s.
REJECTED ADDRESSES. 1s.
PENN'S HINTS ON ANGLING. 1s.
MURRAY'S (CAPT. A.) Naval Life and Services of Admiral Sir Philip Durham. 8vo. 5s. 6d.
MUSIC AND DRESS. Two Essays, by a Lady. Reprinted from the "Quarterly Review." Fcap. 8vo. 1s.
NAPIER'S (SIR WM.) English Battles and Sieges of the Peninsular War. Third Edition. Portrait. Post 8vo. 10s. 6d.
— Life of General Sir Charles Napier; chiefly derived from his Journals, Letters, and Familiar Correspondence. Second Edition. Portraits. 4 Vols. Post 8vo. 48s.
NAUTICAL ALMANACK (The). Royal 8vo. 2s. 6d. (Published by Authority.)
NAVY LIST (The Quarterly). (Published by Authority.) Post 8vo. 2s. 6d.
NELSON (ROBERT), Memoir of his Life and Times. By Rev. C. T. SECRETAN, M.A. Portrait. 8vo. 10s. 6d.
NEWBOLD'S (LIEUT.) Straits of Malacca, Penang, and Singapore. 2 Vols. 8vo. 26s.
NEWDEGATE'S (C. N.) Customs' Tariffs of all Nations; collected and arranged up to the year 1855. 4to. 30s.
NICHOLLS' (SIR GEORGE) History of the English Poor-Laws. 2 Vols. 8vo. 28s.
— History of the Irish Poor-Law. 8vo. 14s.
— History of the Scotch Poor-Law. 8vo. 12s.
— (Rev. H. G.) Historical and Descriptive Account of the Forest of Dean: from Sources Public, Private, Legendary, and Local. Woodcuts, &c. Post 8vo. 10s. 6d.
NICOLAS' (SIR HARRIS) Historic Peerage of England. Exhibiting the Origin, Descent, and Present State of every Title of Peerage which has existed in this Country since the Conquest. Being a New Edition of the "Synopsis of the Peerage." Revised and Continued to the Present Time. By WILLIAM COURTHOPE, Somerset Herald. 8vo. 30s.
NIMROD On the Chace—The Turf—and The Road. Reprinted from the "Quarterly Review." Woodcuts. Fcap. 8vo. 3s. 6d.
O'CONNOR'S (R.) Field Sports of France; or, Hunting, Shooting, and Fishing on the Continent. Woodcuts. 12mo. 7s. 6d.
OLIPHANT'S (LAURENCE) Journey to Katmandu, with Visit to the Camp of the Nepaulese Ambassador. Fcap. 8vo. 2s. 6d.
OXENHAM'S (REV. W.) English Notes for Latin Elegiacs; designed for early Proficients in the Art of Latin Versification, with Prefatory Rules of Composition in Elegiac Metre. Fourth Edition. 12mo. 3s. 6d.
PAGET'S (JOHN) Hungary and Transylvania. With Remarks on their Condition, Social, Political, and Economical. Third Edition. Woodcuts. 2 Vols. 8vo. 18s.
PARIS' (Dr.) Philosophy in Sport made Science in Earnest; or, the First Principles of Natural Philosophy inculcated by aid of the Toys and Sports of Youth. Eighth Edition. Woodcuts. Post 8vo. 7s. 6d.
PARKYNS' (MANSFIELD) Personal Narrative of Three Years' Residence and Adventures in Abyssinia. Woodcuts. 2 Vols. 8vo. 30s.
PEEL'S (SIR ROBERT) Memoirs. Left in MSS. Edited by EARL STANHOPE and the Right Hon. EDWARD CARDWELL. 2 Vols. Post 8vo. 7s. 6d. each.
PEILE'S (REV. DR.) Agamemnon and Choephorœ of Æschylus. A New Edition of the Text, with Notes. Second Edition. 2 Vols. 8vo. 9s. each.
PENN'S (RICHARD) Maxims and Hints for an Angler and Chessplayer. New Edition. Woodcuts. Fcap. 8vo. 1s.
PENROSE'S (REV. JOHN) Faith and Practice; an Exposition of the Principles and Duties of Natural and Revealed Religion. Post 8vo. 8s. 6d.
— (F. C.) Principles of Athenian Architecture, and the Optical Refinements exhibited in the Construction of the Ancient Buildings at Athens, from a Survey. With 40 Plates. Folio. 5l. 5s.
PERCY'S (JOHN, M.D.) Metallurgy; or, the Art of Extracting Metals from their Ores and adapting them to various purposes of Manufacture. First Division—Slags, Fire-Clays, Fuel-Copper, Zinc, and Brass. Illustrations. 8vo. 21s.
PERRY'S (SIR ERSKINE) Bird's-Eye View of India. With Extracts from a Journal kept in the Provinces, Nepaul, &c. Fcap. 8vo. 5s.
PHILLIPS' (JOHN) Memoirs of William Smith, LL.D. (the Geologist). Portrait. 8vo. 7s. 6d.
— Geology of Yorkshire, The Yorkshire Coast, and the Mountain-Limestone District. Plates. 4to. Part I., 20s. —Part II., 30s.
— Rivers, Mountains, and Sea Coast of Yorkshire. With Essays on the Climate, Scenery, and Ancient Inhabitants of the Country. Second Edition, with 36 Plates. 8vo. 15s.
PHILPOTT'S (BISHOP) Letters to the late Charles Butler, on the Theological parts of his "Book of the Roman Catholic Church;" with Remarks on certain Works of Dr. Milner and Dr. Lingard, and on some parts of the Evidence of Dr. Doyle. Second Edition. 8vo. 16s.
PHIPPS' (HON. EDMUND) Memoir, Correspondence, Literary and Unpublished Diaries of Robert Plumer Ward. Portrait. 2 Vols. 8vo. 28.s.
POPE'S (ALEXANDER) Life and Works. A New Edition. Containing nearly 500 unpublished Letters. Edited with a New LIFE, Introductions and Notes. By Rev. WHITWELL ELWIN. Portraits. Vol. I. 8vo. (In the Press.)
PORTER'S (REV. J. L.) Five Years in Damascus. With Travels to Palmyra, Lebanon, and other Scripture Sites. Map and Woodcuts. 2 Vols. Post 8vo. 21s.
— Handbook for Syria and Palestine: including an Account of the Geography, Histoy, Antiquities, and Inhabitants of these Countries, the Peninsula of Sinai, Edom, and the Syrian Desert. Maps. 2 Vols. Post 8vo. 24s.
— (MRS.) Rational Arithmetic for Schools and for Private Instruction. 12mo. 3s. 6d.
PRAYER-BOOK (The Illustrated), with 1000 Illustrations of Borders, Initials, Vignettes, &c. Medium 8vo. 21s.
[page] 27 PUBLISHED BY MR. MURRAY.
PRECEPTS FOR THE CONDUCT OF LIFE. Extracted from the Scriptures. Second Edition. Fcap. 8vo. 1s.
PRINSEP'S (JAS.) Essays on Indian Antiquities, Historic, Numismatic, and Palæographic, with Tables. Edited by EDWARD THOMAS. Illustrations. 2 Vols. 8vo. 52s. 6d.
PROGRESS OF RUSSIA IN THE EAST. An Historical Summary, continued to the Present Time. Third Edition. Map. 8vo. 6s. 6d.
PUSS IN BOOTS. With 12 Illustrations; for Old and Young. By OTTO SPECKTER. 16mo. 1s. 6d.; or Coloured, 2s.
QUARTERLY REVIEW (THE). 8vo. 6s.
RAWLINSON'S (REV. GEORGE) Herodotus. A New English Version. Edited with Notes and Essays. Assisted by SIR HENRY RAWLINSON and SIR J. G. WILKINSON. Second Edition. Maps and Woodcut Vols. 8vo. 48s.
— Historical Evidences of the truth of the Scripture Records stated anew, with special reference to the Doubts and Discoveries of Modern Times; the Bampton Lectures for 1859. Second Edition. 8vo. 14s.
— Five Great Monarchies of the Ancient World. Or the History, Geography, and Antiquities of Chaldæa, Assyria, Babylonia, Media, and Persia. Illustrations. Vol. I. 8vo. 18s.
REJECTED ADDRESSES (THE). By JAMES AND HORACE SMITH. New Edition. Fcap. 8vo. 1s., or Fine Paper, with Portrait, fcap.8vo, 5s.
RICARDO'S (DAVID) Political Works. With a Notice of his Life and Writings. By J. R. M'CULLOCH. New Edition. 8vo. 16s.
RIPA'S (FATHER) Memoirs during Thirteen Years' Residence at the Court of Peking. From the Italian. Post 8vo. 2s.
ROBERTSON'S (CANON) History of the Christian Church, From the Apostolic Age to the Concordat of Worms, A.D. 1123. Second Edition. 2 Vols. 8vo. 34s.
— Life of Archbishop Becket. Illustrations. Post 8vo. 9s.
ROBINSON'S (REV. DR.) Biblical Researches in the Holy Land. Being a Journal of Travels in 1838, and of Later Researches in 1852. Maps. 3 Vols. 8vo. 36s.
ROMILLY'S (SIR SAMUEL) Memoirs and Political Diary. By his SONS. Third Edition. Portrait. 2 Vols. Fcap. 8vo. 12s.
ROSS'S (SIR JAMES) Voyage of Discovery and Research in the Southern and Antarctic Regions, 1839-43. Plates. 2 Vols. 8vo. 36s.
ROWLAND'S (DAVID) Manual of the English Constitution; a Review of its Rise, Growth, and Present State. Post 8vo. 10s. 6d.
RUNDELL'S (MRS.) Domestic Cookery, founded on Principles of Economy and Practice, and adapted for Private Families. New and Revised Edition. Woodcuts. Fcap. 8vo. 5s.
RUSSELL'S (J. RUTHERFURD, M.D.) Art of Medicine—Its History and its Heross. Portraits. 8vo. 14s.
RUSSIA; A Memoir of the Remarkable Events which attended the Accession of the Emperor Nicholas. By BARON M. KORFF, Secretary of State. 8vo. 10s. 6d. (Published by Imperial Command.)
BUXTON'S (GEORGE F.) Travels in Mexico; with Adventures among the Wild Tribes and Animals of the Prairies and Rocky Mountains. Post 8vo. 3s. 6d.
SALE'S (LADY) Journal of the Disasters in Affghanistan. Post 8vo. 12s.
— (SIR ROBERT) Brigade in Affghanistan. With an Account of the Defence of Jellalabad. By REV. G. R. GLEIG. Post 8vo. 2s.
SANDWITH'S (HUMPHRY) Siege of Kars and Resistance by the Turkish Garrison under General Williams. Post 8vo. 3s. 6d.
SCOTT'S (G. GILBERT) Secular and Domestic Architecture, Present and Future. Second Edition. 8vo. 9s.
— (Master of Baliol) Sermons Preached before the University of Oxford. Post 8vo. 8s. 6d.
SCROPE'S (WILLIAM) Days of Deer-Stalking; with some Account of the Red Deer. Third Edition. Woodcuts. Crown 8vo. 20s.
— Days and Nights of Salmon Fishing in the Tweed; with a short Account of the Salmon. Second Edition. Woodcuts. Royal 8vo. 31s. 6d.
—(G. P.) Memoir of Lord Sydenham, and his Administration in Canada. Second Edition. Portrait. 8vo. 9s. 6d.
— Geology and Extinct Volcanoes of Central France. Second Edition. Illustrations. Medium 8vo. 30s.
SELF-HELP. With Illustrations of Character and Conduct. By SAMUEL. SMILES. Fortieth Thousand. Post 8vo. 6s.
SENIOR'S (N. W.) Suggestions on Popular Education. 8vo. 9s.
SHAFTESBURY (LORD CHANCELLOR); Memoirs of his Early Life. With his Letters, &c. By W. D. CHRISTIE. Portrait. 8vo. 10s. 6d.
SHAW'S (J. F.) Outlines of English Literature for Students.Second Edition. Revised. Post 8vo. (In the Press.)
SIERRA LEONE; Described in Letters to Friends at Home. By A LADY. Post 8vo. 3s. 6d.
SMILES' (SAMUEL) Lives of Engineers; with an Account of their Principal Works, and a History of Inland Communication in Britain. Portraits and numerous Woodcuts. Vols. I. and II. 8vo. 42s.
— Story of the Life of George Stephenson. Woodcuts. Eighteenth Thousand. Post 8vo. 6s.
— Self-Help. With Illustrations of Character and Conduct. Fortieth Thousand. Post 8vo. 6s.
— Workmen's Earnings, Savings, and Strikes. Fcap. 8vo. 1s. 6d.
SOMERVILLE'S (MARY) Physical Geography. Fifth Edition. Portrait. Post 8vo. (In the Press.)
— Connexion of the Physical Sciences. Ninth Edition. Woodcuts. Post 8vo. 9s.
SOUTH'S (JOHN F.) Household Surgery; or, Hints on Emergencies. Seventeenth Thousand. Woodcuts. Fcp. 8vo. 4s. 6d.
SOUTHEY'S (ROBERT) Book of the Church. Seventh Edition. Post 8vo. 7s. 6d.
— Lives of Bunyan & Cromwell. Post 8vo. 2s.
SPECKTER'S (OTTO) Puss in Boots. With 12 Woodcuts. Square 12mo. 1s. 6d. plain, or 2s. coloured.
— Charmed Roe; or, the Story of the Little Brother and Sister. Illustrated. 16mo.
[page] 29 PUBLISHED BY MR. MURRAY.
SMITH'S (DR. WM.) Dictionary of the Bible; its Antiquities, Biography, Geography, and Natural History. Second Edition. Woodcuts. Vol.1. 8vo. 42s.
— Greek and Roman Antiquities. 2nd Edition. Woodcuts. 8vo 42s.
— Biography and Mythology. Woodcuts. 3 Vols. 8vo. 5l. 15s. 6d.
— Geography. Woodcuts. 2 Vols. 8vo. 80s.
— Latin-English Dictionary. Based upon the Works of Forcellini and Freund. Ninth Thousand. 8vo. 21s.
—English-Latin Dictionary. 8vo. & 12mo. (In preparation.)
—Mediæval Latin-English Dictionary. Based on the Work of DUCANGE. 8vo. (In preparation.)
— Classical Dictionary. 5th Edition. 750 Woodcuts. 8vo. 18s.
— Smaller Classical Dictionary. Twentieth Thousand. 200 200 Woodcuts. Crown 8vo. 7s. 6d.
—Dictionary of Antiquities. Sixteenth Thousand. 200 Woodcuts. Crown 8vo. 7s. 6d.
— Latin-English Dictionary. Twentieth Thousand. Square 12mo. 7s. 6d.
— Principia Latina—Part I. A Grammar, Delectus, and Exercise Book, with Vocabularies. 2nd Edition. 12mo. 3s. 6d.
— Principia Latina—Part II. A Reading-book, Mythology, Geography, Roman Antiquities, and History. With Notes and Dictionary. 12mo. 3s. 6d.
— A First Latin Dictionary; applicable for those reading Phædrus, Cornelius Nepos, and Cæsar. 12mo. 3s. 6d.
— Principia Græca; a First Greek Course. A Grammar, Delectus, and Exercise-book with Vocabularies. By H. E. HUTTON, M .A. 2nd Edition. 12mo. 3s.
— (JAMES & HORACE) Rejected Addresses. Fcap. 8vo. 1s; or Fine Paper, with Portrait, fcap. 8vo. 5s.
— (THOMAS ASSHETON) Life and Pursuits. By SIR EARDLEY WILMOT. 2nd Edition. Illustrations. 8vo. 15s.
STANLEY'S (CANON) Lectures on the History of the Eastern Church. Second Edition. Plans. 8vo. 16s.
— Lectures on the History of the Jewish Church. From ABRAHAM TO SAMUEL. 8vo. (In the Press.)
— ADDRESSES AND CHARGES OF BISHOP STANLEY. With Memoir. Second Edition. 8vo. 10s. 6d.
—Unity of Evangelical and Apostolical Teaching. Second Edition. Post 8vo. 7s. 6d.
— St. Paul's Epistles to the Corinthians, with Notes and Dissertations. Second Edition. 8vo. 18s.
— Historical Memorials of Canterbury. Third Edition. Woodcuts. Post 8vo. 7s. 6d.
— Sinai and Palestine, in Connexion with their History. Sixth Edition. Map. 8vo. 16s.
ST. JOHN'S (CHARLES) Wild Sports and Natural History of the Highlands. Post 8vo. 3s. 6d.
—(BAYLE) Adventures in the Libyan Desert and the Oasis of Jupiter Ammon. Woodcuts. Post 8vo. 2s.
STEPHENSON (GEORGE) The Railway Engineer. The Story of his Life. By SAMUEL SMILES. Eighteenth Thousand. Woodcuts. Post 8vo. 6s.
STOTHARD'S (THOS.) Life. With Personal Reminiscences. By Mrs. BRAY. With Portrait and 60 Woodcuts. 4to.
STREET'S (G. E.) Brick and Marble Architecture of Italy in the Middle Ages. Plates. 8vo. 21s.
STRIFE FOR THE MASTERY. Two Allegories. With Illustrations. Crown 8vo. 6s.
STUDENT'S HUME. A History of England from the Invasion of Julius Cæsar to the Revolution of 1688. Based on the Work by DAVID HUME. Continued to 1858. Twentieth Thousand. Woodcuts. ost 8vo. 7s. 6d.
—HISTORY OF FRANCE; From the Earliest Times to the Establishment of the Second Empire, 1852. Woodcuts. Post 8vo. 7s. 6d.
— HISTORY OF GREECE; from the Earliest Times to the Roman Conquest. With the History of Literature and Art. By WM. SMITH, LL.D. 20th Thousand. Woodcuts. Crown 8vo. 7s. 6d. (Questions. 2s.)
*** A SMALLER HISTORY OF GREECE, for Junior Classes. 12mo. 3s. 6d.
— HISTORY OF ROME; from the Earliest Times to the Establishment of the Empire. With the History of Literature and Art. By H. G. LIDDELL, D.D. 15th Thousand. Woodcuts. Crown vo. 7s. 6d.
*** SMALLER HISTORY OF ROME, for Junior Classes. By Dr. WM. SMITH. 12mo. 3s. 6d.
— GIBBON; an Epitome of the History of the Decline and Fall of the Roman Empire. Incorporating the Researches of Recent Commentators. 6th Thousand. Woodcuts. Post 8vo. 7s. 6d.
— MANUAL OF ANCIENT GEOGRAPHY. Based on the larger Dictionary of Greek and Roman Geography. Woodcuts. Post 8vo. 9s.
— THE ENGLISH LANGUAGE; A Series of Lectures. By GEORGE P. MARSH. Post 8vo. (Nearly Ready.)
— GREEK GRAMMAR; from the German of PROFESSOR CURTIUS. Edited by DR. WM. SMITH. Post 8vo. (In the Press.)
SWIFT'S (JONATHAN) Life, Letters and Journals. By JOHN FORSTER. 8vo. (In Preparation.)
— Works. Edited, with Notes. By JOHN FORSTER. 8vo. (In Preparation.)
SYME'S (JAS.) Principles of Surgery. Fourth Edition. 8vo. 14s.
TAIT'S (BISHOP) Dangers and Safeguards of Modern Theology. 8vo. 9s.
TAYLOR'S (HENRY) Notes from Life. Fcap 8vo. 2s.
THOMSON'S (BISHOP) Sermons Preached in Lincoln's Inn Chapel. 8vo. 10s. 6d.
— (DR.) Story of New Zealand; Past and Present—Savage and Civilised. Second Edition. Illustrations. 2 Vols. Post 8vo. 24s.
THREE-LEAVED MANUAL OF FAMILY PRAYER; arranged so as to save the trouble of turning the Pages backwards and forwards. Royal 8vo. 2s.
[page] 31 PUBLISHED BY MR. MURRAY.
TICKNOR'S (GEORGE) History of Spanish Literature. With Criticisms on particular Works, and Biographical Notices of Prominent Writers. Second Edition. 3 Vols. 8vo. 24s.
TOCQUEVILLE'S (M.DE) State of France before the Revolution, 1789, and on the Causes of that Event. Translated by HENRY REEVE, ESQ. 8vo. 14s.
TREMENHEERE'S (H. S.) Political Experience of the Ancients, in its bearing on Modern Times. Feap. 8vo. 2s. 6d.
— Notes on Public Subjects, made during a Tour in the United States and Canada. Post 8vo. 10s. 6d.
— Constitution of the United States compared with our own. Post 8vo. 9s. 6d.
TRISTRAM'S (H. B.) Great Sahara; or Wanderings South of the Atlas Mountains. Illustrations. Post 8vo. 15s.
TWISS' (HORACE) Public and Private Life of Lord Chancellor Eldon, with Selections from his Correspondence. Portrait. Third Edition. 2 Vols. Post 8vo. 21s.
TYNDALL'S (JOHN) Glaciers of the Alps. Being a Narrative of various Excursions among them, and an Account of Three Years' Observations and Experiments on their Motion, Structure, and General Phenomena. Woodcuts. Post 8vo. 14s.
TYTLERS'S (PATRICK FRASER) Memoirs. By REV. J. W. BURGON, M.A. Second Edition. 8vo. 9s.
UBICINI'S (M. A.) Letters on Turkey and its Inhabitants—the Moslems, Greeks, Armenians, &c. Translated by LADY EASTHOPE. 2 Vols. Post 8vo. 21s.
VAUGHAN'S (REV. DR.) Sermons preached in Harrow School. 8vo. 10s. 6d.
VENABLES' (REV. R. L.) Domestic Scenes in Russia during a Year' Residence, chiefly in the Interior. Second Edition. Post 8vo. 5s.
VOYAGE to the Mauritius and back, touching at the Cape of Good Hope and St. Helena. By Author of "PADDIANA." Post 8vo. 9s. 6d.
WAAGEN'S (DR.) Treasures of Art in Great Britain. Begin an Account of the Chief Collections of Paintings, Sculpture, Manuscripts, Miniatures, &c. &c., in this Country. Obtained from Personal Inspection during Visits to England. 3 Vols. 8vo. 36s.
— Galleries and Cabinets of Art in England. Being an Account of more than Forty Collections, visited in 1854-56. With Index. 8vo. 18s.
WADDINGTON'S (DEAN) Condition and Prospects of the Greek Church. New Edition. Fcap. 8vo. 3s. 6d.
WAKEFIELDS'S (E. J.) Adventures in New Zealand. With some Account of the Beginning of the British Colonisation of the Island. Map. 2 Vols. 8vo. 28s.
WALKS AND TALKS. A Story-book for Young Children. By AUNT IDA. With Woodcuts. 16mo. 5s.
WALSH'S (SIR JOHN) Practical Results of the Reform Bill of 1832. 8vo. 5s. 6d.
WARD'S (ROBERT PLUMER) Memoir, Correspondence, Literary and Unpublished Diaries and Remains. By the HON. EDMUD PHIPPS. Portrait. 2 Vols. 8vo. 28s.
[page] 32 LIST OF WORKS PUBLISHED BY MR. MURRAY.
WATT'S (JAMES) Life. Incorporating the most interesting passages from his Private and Public Correspondence. By JAMES P. MUIRHEAD, M.A. Second Edition. Portrait. 8vo. 16s.
— Origin and Progress of his Mechanical Inventions. Illustrated by his Correspondence. By J. P. MUIRHEAD. Plates. 3 Vols. 8vo. 45s.
WILKIE'S (SIR DAVID) Life, Journals, Tours, and Critical Remarks on Works of Art, with a Selection from his Correspondence. By ALLAN CUNNINGHAM. Portrait. 3 Vols. 8vo. 42s.
WOOD'S (LIEUT.) Voyage up the Indus to the Source of the River Oxus, by Kabul and Badakhshan. Map. 8vo. 14s.
WELLINGTON'S (THE DUKE OF) Despatches during his various Campaigns. Compiled from Official and other Authentic Documents. By COL. GURWOOD, C.B. New Enlarged Edition. 8 Vols. 8vo. 21s. each.
— Supplementary Despatches, and other Papers. Edited by his SON. Vols. I. to VIII. 8vo. 20s. each.
— Selections from his Despatches and General Orders. By COLONEL GURWOOD. 8vo. 18s.
— Speeches in Parliament. 2 Vols. 8vo. 42s.
WILKINSON'S (SIR J. G.) Popular Account of the Private Life, Manners, and Customs of the Ancient Egyptians. New Edition. Revised and Condensed. With 500 Woodcuts. 2 Vols. Post 8vo. 12s.
— Dalmatia and Montenegro; with a Journey to Mostar in Hertzegovina, and Remarks on the Slavonic Nations. Plates and Woodcuts. 2 Vols. 8vo. 42s.
— Handbook for Egypt.—Thebes, the Nile, Alexandria, Cairo, the Pyramids, Mount Sinai, &c. Map. Post 8vo. 15s.
—On Colour, and on the Necessity for a General Diffusion of Taste among all Classes; with Remarks on laying out Dressed or Geometrical Gardens. With Coloured Illustrations and Woodcuts. 8vo. 18s.
— (G. B.) Working Man's Handbook to South Australia; with Advice to the Farmer, and Detailed Information for the several Classes of Labourers and Artisans. Map. 18mo. 1s. 6d.
WILSON'S (DANIEL, D.D., BISHOP OF CALCUTTA) Life, with Extracts from his Letters and Journals. By Rev. JOSIAH BATEMAN. New and Condensed Edition. Illustrations. Post 8vo. 9s.
— (GENL. SIR ROBERT) Secret History of the French Invasion of Russia, and Retreat of the French Army, 1812. Second Edition. 8vo. 15s.
— Private Diary of Travels, Personal Services, and Public Events, during Missions and Employments in Spain, Sicily, Turkey, Russia, Poland, Germany, &c. 1812-14. 2 Vols. 8vo. 26s.
WORDSWORTH'S (REV. DR.) Journal of a Tour in Athens and Attica. Third Edition. Plates. Post 8vo. 8s. 6d.
— Pictorial, Descriptive, and Historical Account of Greece, with a History of Greek Art, by G. SCHARF, F.S.A. New Edition. With 600 Woodcuts. Royal 8vo. 28s.
WORNUM (RALPH). A Biographical Dictionary of Italian Painters: with a Table of the Contemporary Schools of Italy. By a LADY. Post 8vo. 6s. 6d.
WROTTESLEY'S (LORD) Thoughts on Government and Legislation. Post 8vo. 7s. 6d.
YOUNG'S (DR. THOS.) Life and Miscellaneous Works, edited by DEAN PEACOCK and JOHN LEITCH. Portrait and Plates. 4 Vols. 8vo. 15s. each.
BRADBURY AND EVANS, PRINTERS, WHITEPRIARS.