No. IV. of Part I.] [Completion of "FOSSIL MAMMALIA."] [Price 6s.
THE
ZOOLOGY
OF
THE VOYAGE OF H.M.S. BEAGLE,
UNDER THE COMMAND OF CAPTAIN FITZROY, R.N.,
DUHING THE YEARS
1832 to 1836.
PUBLISHED WITH THE APPROVAL OF THE LORDS COMMISSIONERS OF HER MAJESTY'S TREASURY.
Edited and superintended by
CHARLES DARWIN, ESQ. M.A., F.R.S., Sec. G.S.
NATURALIST TO THE EXPEDITION.
FOSSIL MAMMALIA,
BY RICHARD OWEN, ESQ., F.R.S.
PROFESSOR OF ANATOMY AND PHYSIOLOGY TO THE ROYAL COLLEGE OF SURGEONS, LONDON; CORRESPONDING MEMBER OP THE INSTITUTE OP FRANCE, ETC. ETC.
LONDON:
PUBLISHED BY SMITH, ELDER AND CO. 65, CORNHILL.
MDCCCXL.
XIII.
April.
STEWART AND MURRAY, OLD BAILEY.
served, is smooth and vertical. Each superior maxillary bone contains the sockets of five teeth, occupying an antero-posterior extent of three inches seven lines, (Pl. XXII and XXIII. fig. 3). The posterior alveolus is situated just behind the transverse line, extending across the anterior boundary of the orbits; the remaining sockets of the molar series extend forwards three inches in front of the orbits. In the Megatherium, the roots of the five superior molars are all situated behind the anterior boundary of the orbit: in the Orycteropus, on the contrary, the grinders are all placed in advance of the orbit; so that the Scelidothere resembles that species more than the Megathere in the relative location of the teeth. The palatal interspace between the roots of the last molar tooth of each series is eleven lines; the palate gradually though slightly widens, as it advances forwards: the posterior margin of the palate is terminated by an acute-angled notch. In the breadth of the bony palate the Scelidothere is intermediate between the Megathere and Orycterope.
The anterior of the upper molars is represented at fig. 3, 4, and 5, Pl. XXI., and at 1, fig. 3, Pl. XXIII.; it corresponds closely in form and size with the opposite molar below; the base of the triangle given by its transverse section is turned inwards and obliquely forwards.
The second molar of the upper jaw, also presents in transverse section a triangular form, with the angles rounded off; but the inner side of the tooth is traversed by a longitudinal groove. The largest diameter of the transverse section, which is placed obliquely as regards the axis of the skull, measures ten lines and a half; the opposite diameter of the tooth is six lines.
The third and fourth molars present the same form and size, and relative position as the second.
The fifth molar is the smallest of the series; its transverse section gives an inequilateral triangle, with the corners rounded off; the broadest side is turned outwards, and is slightly concave; the antero-posterior diameter of this tooth is seven lines; the transverse four lines. The length of the teeth in the upper jaw is about two inches and a half.
It is almost superfluous to observe that the teeth of the Scelidothere, as in other Bruta, are without fangs, and have their inserted base excavated by large conical cavities, for the lodgment of a persistent pulp. The tooth is composed of a small central body of coarse ivory or 'dentine,' traversed by medullary canals, which at the periphery of the coarse dentine anastomose by loops, from the convexity of which the calcigerous tubes are given off which form the fine dentine: the layer of this substance, which immediately surrounds the coarse dentine, is about one line and a half in thickness, and the whole is invested with a very thin coating of cement. The teeth of the Scelidothere thus
M
[page] 82 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
present a more resisting structure than do those of the Mylodon; having a larger proportion of the dense ivory composed of the minute calcigerous tubes, and a much smaller proportion of the softer external cæmentum; in this respect the Scelidothere recedes farther from Megathere, and approaches nearer the Armadillos than does the Mylodon.
The lower jaw resembles, in the general form of the posterior moiety which is here preserved, that of the Sloth and Mylodon more than that of any other Edentate species. Its deep posterior angle is produced backwards, and a broad coronoid process rises and nearly fills the zygomatic space; the condyle is flat, as the glenoid surface has already indicated; its transverse diameter is an inch and eight lines; its antero-posterior diameter seven lines: it is principally extended inwards beyond the vertical line of the ascending ramus. The lower contour of the jaw describes an undulating line; which, commencing from the posterior angle, is at first gently convex, then slightly concave, then again convex, below the alveoli of the teeth, where it is rounded and expanded, as in the Orycterope. The fractured condition of the right ramus of this part fortunately exposed the roots of the four grinding teeth, which constitute the dental series on each side of the lower jaw. The length of the jaw occupied by these four alveoli is three inches ten lines, which exceeds a little that of the opposed five grinders above; the ramus of the jaw gradually diminishes in all its dimensions anterior to the molar teeth; the dental canal passes in a gentle curve below, and on the inner side of the alveoli, whence it gradually inclines to the outer wall of the jaw.
The whole ascending ramus of the jaw consists of a very thin plate of bone; it is slightly concave on the inner side, and the inferior margin of the produced angle inclines inwards, as in the Mylodon and Sloth; it is impressed on the outer side with two shallow depressions, and two parallel ridges, both following the gentle curvature of the part. There is a foramen on the outer side of the ramus at the anterior part of the base of the coronoid process corresponding with that in the lower jaw of the Mylodon, but the longitudinal channel which runs along the outer side of the alveolar processes is wanting, and the expansion at the base of those processes is more sudden and relatively greater; the general correspondence, however, between these lower jaws is such as would lead to the idea that they belonged to animals of the same genus, were it not that the teeth present modifications of form in the Scelidothere, as distinct from those of the Mylodon, as are any of the minor dental differences on which genera or sub-genera of existing Mammalia are founded in the present state of Zoological Classification.
To make this distinction more readily intelligible, I have given a view of the transverse section of the teeth in the right ramus of the lower jaw (fig. 4, Pl. XXIII.), corresponding with that of the Mylodon Darwinii, (Pl. XVII., fig. 5). In the present
sub-genus the antero-posterior extent of the four alveoli of the lower jaw nearly equals four inches, and is relatively greater than in the Mylodon, although the teeth are placed closer together; this is owing to their greater relative size. The first molar tooth presents the simplest form; its transverse section is a compressed inequilateral triangle with the angles rounded off; the longest diameter of this section which is parallel with the inner alveolar border is eleven lines, the transverse diameter almost six lines; the base or broadest side of the triangle is turned inwards, and is slightly concave; the two smaller sides are also slightly concave.
The second molar is placed more obliquely in the jaw; the long axis of its transverse section intersects at an acute angle that of the jaw itself; the transverse section presents a compressed or oblong form, with the larger end next the outer side, and the smaller end next the inner side of the jaw; this end is simply rounded, but the outer end presents a sinuosity, corresponding to a broad groove which traverses the whole length of the outer side of the tooth; the anterior, which corresponds to the internal side or base of the transverse section of the preceding molar, is slightly concave.
The third molar has nearly the same form and relative position as the preceding; the long diameter of the transverse section is, in both, ten lines and a half; the principal transverse diameter is, in the second molar five lines, in the third nearly six; the difference of form observable in these as compared with the two middle grinders of the Mylodon is well marked; in the latter these teeth are impressed with a longitudinal groove on their inner sides; in the Scelidothere they have a similar impression along their outer but not along the inner side.
In the last molar the resemblance is much closer, and the modification of form by which it differs from the preceding ones is of the same kind; the transverse section gives an irregular oblong figure with its axis nearly parallel with that of the jaw, and constricted at the middle by sinuosities produced by two wide channels which traverse longitudinally, one the outer, the other the inner side of the tooth; the latter groove is much wider and shallower in the Scelidothere than in the Mylodon. The two lobes produced by these grooves are more equal in Scelidothere; the anterior one is concave on its anterior surface instead of convex as in the Mylodon; the posterior one is more compressed; the longitudinal or antero-posterior diameter of the transverse section of this tooth is one inch five lines; the greatest transverse diameter is nine lines; the diameter of the isthmus joining the lobes is three lines and a half; the entire length of this tooth is three inches three lines.*
* It requires little stretch of imagination to conceive that this more complex posterior tooth (Pl. XXIII, fig. 4, 4) in the lower jaw is the representative of the two smaller posterior teeth (ib. fig. 3, 4, and 5) of the upper jaw conjoined.
M 2
[page] 84 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
VERTEBRAL COLUMN.
Of this part of the skeleton of the Scelidothere, Mr. Darwin's specimen includes, as is represented in Plate XX., the cervical, part of the dorsal, and the sacral series of vertebræ in a more or less perfect condition.
The cervical vertebræ present the ordinary mammalian number, seven, and are free, or so articulated as to have permitted reciprocal movement upon each other. Their transverse processes are perforated as usual for the vertebral arteries. These processes in the atlas are remarkable for their great breadth, length, and thickness; and indicate the muscular forces which must have worked the head upon the spine to have been very powerful. The axis is provided with a robust 'processus dentatus,' having a base equal in breadth to the body of the axis itself; and a smooth articular convexity on the side of the apex on which the ring of the atlas rotated. The line of union between the axis and its characteristic process, which here resembles the body of an abortive vertebra, is very distinct. The transverse processes of the vertebra dentata are comparatively feeble, but this condition is amply compensated for by the great development of the spinous process. (Pl. XXIV. fig. 1.) This process is bent backwards at nearly a right angle, overlaps with its reflected extremity the spine of the third cervical vertebra, and rests by its base, on the under part of which are the posterior articular surfaces, upon the broad and strong anterior oblique processes of the third vertebra.
The third, fourth, fifth, and sixth cervical vertebræ have moderately developed and pointed spinous processes: their transverse processes are broad, and extend obliquely backwards, and slightly overlap each other. On the under part of the transverse process of the sixth cervical vertebra there is the fractured base of what I conjecture to have been an expanded aliform plate, analogous to that observable in the corresponding vertebra of the Orycterope. The seventh cervical vertebra has part of the articular depression for the head of the first rib upon each side of its body: the transverse process is feebly developed, but the spine is double the height and size of those of the preceding vertebræ.
The spinous process of the first dorsal vertebra in like manner rises to twice the height of the preceding spine of the seventh cervical, and preserves an equal antero-posterior diameter from its base to its summit, which is thick and slightly bent backwards: four or five succeeding dorsal vertebræ give evidence of having been surmounted by spines of equal height and strength. The transverse processes of these dorsal vertebræ present bold concavities on their inferior part for the reception of the tubercles of the ribs, and they gradually ascend upon the base of the spines as the vertebræ are placed further back, so as to increase the expansiveness
of the chest. The state of the fossil did not afford further information as to the condition of this part of the vertebral column, but the parts which have been preserved are precisely those from which the most interesting inferences as to the affinities and habits of the extinct quadruped can be deduced.
Whether the Megatherium be most nearly allied to the tribes of the Sloth or Armadillo has been a question under recent discussion, and, as a corollary of this problem, whether its habits were those of a scansorial or of a fossorial quadruped. For, strange as it may appear at first sight, there have not been wanting arguments, and those urged by an anatomist to whom we owe much novel and interesting information respecting the extinct Edentata, in support of the belief that the Megatherium, gigantic and ponderous as must have been its frame, actually climbed trees like a Sloth, and had claws and feet organised for prehensile actions, and not in accordance with that type by which they are usually adapted for digging up the soil.*
Now, in whatever degree the Megatherium may be involved in this question, the smaller Megatherioid species at present under consideration must be at least equally implicated in it. In the adaptation of the frame of a mammiferous quadruped for especial and peculiar actions and modes of life, such as for climbing and living in trees, or for burrowing and seeking concealment in the earth, not only the immediate instruments, as the feet, are modified, but the whole of the osseous and muscular fabric is more or less impressed with corresponding adaptations, whilst at the same time these special adjustments are invariably subordinated to the type of organization which characterizes the group.
The type of the order Bruta or Edentata is well-marked; one or more claws of unusual length and strength, characterize the fore-feet and sometimes the hind-feet in every genus, and the term 'Macronykia' would more aptly designate them than the term which Cuvier substituted for the good old Linnæan appellation. The uniform absence of true roots to the teeth, where these are present, is another general character; the skeleton exhibits many well-marked peculiarities common to the whole order; while at the same time it is modified in various modes and degrees in accordance with the peculiar habits and exigencies of the species.
One of the regions of the skeleton which manifests adaptive modifications of this kind in the most remarkable degree is the cervical division of the vertebral column. In one edentate species it is lengthened out by two additional vertebræ more than in any other mammal; in another it is reduced by anchylosis to as great an extent below the regular number of moveable pieces: and these, the two most opposite conditions of the cervical vertebræ which are to be met with in the mammiferous class are related to equally diverse and opposite habits of life.
* Lund, Videnskabernes Selskabs, Natur: og Mathem. Afhandlinger, Kiöbenhavn, vol. viii.
[page] 86 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
With respect to the Ai, or three-toed Sloth, "an animal, great part of whose life, when not engaged in eating, is spent in sleeping on trees,—an easy attitude for repose is most essential to its comfortable existence; and accordingly we find, that the auxiliary vertebræ at the base of the neck contribute to produce that flexibility of this organ which allows the head of the animal to incline forwards and rest upon its bosom." Dr. Buckland, from whose Paper on the "Adaptation of the Structure of the Sloths to their peculiar Mode of Life,"* the preceding judicious physiological remark is quoted, adduces the authority of Mr. Burchell in proof that the Sloth can in a remarkable manner and with great facility twist its head quite round, and look in the face of a person standing directly behind it, while at the same time the body and limbs remain unmoved. A single glance at the length and slenderness of the cervical region of the spine, and of the feeble condition of the transverse and spinous processes in the vertebræ composing that part of the skeleton of the Sloth, is enough to show its adaptation to increase the rotatory motion and flexibility of the neck.
In describing the skeleton of a species of Armadillo (Dasypus 6-cinctus, Linn.)† I was led in like manner to point out the subserviency of the peculiarities of the cervical vertebræ to the habits and mode of life of that animal; observing that the "anchylosis of the cervical vertebræ obtains in the Cetacea, as well as in the genus Dasypus, and that as in the aquatic order this firm connexion of the cervical vertebræ assists materially in enabling the head to overcome the resistance of the dense fluid through which they perpetually move, so in the Armadillos a like advantage may be derived from this structure during the act of displacing the denser material in which they excavate their retreats."‡
Having in view these well-marked examples of the subserviency of the structure of the bones of the neck to the habits of existing species of the order Bruta, I proceeded to investigate the structure of the corresponding part of the skeleton in the Scelidotherium, hoping thereby to gain a new and useful element in the determination of the problem at present under discussion, as to the affinities and habits of the extinct Megatherioid quadrupeds.
The fossil, in its original state, yielded a view of so much of the anterior part of the bodies of the cervical vertebræ as proved that they were neither so numerous as in the Sloth, nor anchylosed together as in the Armadillos: after a long and careful chiselling at the hard matrix in which they were imbedded, the trans-
* Linn. Trans, vol. xvii. (1833) p. 17.
† Zool. Proceedings, 1832, p. 134.
‡ The anterior prolongation of the sternum in front of the neck and the corresponding anterior position of the clavicles and scapulæ occasions a transference of such a proportion of the moving powers of the head from the cervical vertebræ to these bones in the mole, as renders any modifications of these vertebræ, like those in the Armadillo, uncalled for.
verse and spinous processes were exposed to view, as they are represented in Plates XX. and XXIV. The description of these processes has already been given.
On comparing the cervical vertebræ of the Scelidotherium with those of the existing Bruta, the closest resemblance to them was found in the skeleton of the Orycterope. Now this quadruped, though not so rapid a burrower, or so strictly a subterranean species as the Armadillos, participates, nevertheless, to a certain extent, in their fossorial habits, and is closely allied to them in general structure: it differs from them, indeed, mainly in a modification of the dental system, in the absence of dermal armour, and of anchylosis of the cervical vertebræ. But the advantages which, as a burrower, it would have derived from the latter structure, are compensated for by the shortness of the cervical vertebræ, and by the great development and imbricated or interlocking co-adaptation of the transverse and anterior spinous processes of the cervical vertebræ. The analogous quadruped in the South American Continent—the great ant-eater (myrmecophaga jubata) which uses its powerful compressed fossorial claws for breaking through the hard walls of the habitations of its insect prey, but which does not excavate a subterraneous retreat for itself, presents the cervical vertebræ of a more elongated form, and without that development of the spinous and transverse processes which tend to fix the neck and increase the size of the muscles which move the head: and, if we could conceive that its fore-feet were employed to scratch up vegetable roots, instead of disinterring termites, there would be no reason to expect any modification of the cervical vertebræ as a direct consequence of such a difference in the application of its fossorial extremities: when, therefore, we find that the cervical vertebræ do actually differ in two myrmecophagous species, to the extent observable in the Cape and South American ant-eaters, we arrive legitimately at the conclusion that such difference relates to fossorial habits of the one species, in which habits the other does not participate.
Now, therefore, if this conclusion be just in regard to the Orycterope, it must bear with more force upon the question of the habits of the Scelidotherium as the mechanism for strengthening the connection of cervical vertebræ, and for augmenting the surface of attachment of the muscles which worked the head and neck, is more strongly wrought out in that extinct species.
The great size and strength of the spinous process of the dentata, and the mode in which it is interlocked with the spinous and oblique processes of the third cervical, together with the imbricated disposition of the transverse processes of this and the succeeding vertebræ, and the remarkable height of the dorsal spines, all combine to indicate in a very striking manner, if not to demonstrate, that the conical head of the present species, which is comparatively small and slender, and
[page] 88 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
for its own mere support requiring therefore no such mechanism, was used in aid of the fossorial actions of the extremities.
As the cervical vertebræ of the Megatherium have their processes comparatively weaker than in the Scelidotherium, and the anterior dorsal spines are relatively shorter, it may be concluded, that whatever were the extent or nature of the fossorial labours of the enormous claws with which it was provided, the head did not co-operate with the digging implements in their especial task in the same degree as in the Scelidothere and Orycterope. At the same time there is no modification of the cervical region of the spine of the Megathere corresponding with those which we have seen to be subservient to the arboreal habits of the sloth, a remark which will not be deemed superfluous by those who have perused the acute observations and arguments adduced by M. Lund in favour of the scansorial character of the extremities of the Megatherium and Megalonyx.
The fragments of the dorsal vertebræ and ribs of the Scelidotherium, which are figured in Plate XX, offer no modifications which need detain our attention; they closely conform, excepting in the greater relative height of the anterior dorsal spines, already noticed, with the Megatherioid type. The sacrum manifests in its vast expanse, the great development of the posterior transverse processes to join the ischium, the capacious medullary cavity, and wide nervous foramina, a like conformity with the Megatherium, and a corresponding harmony with the disproportionate bulk of the hind legs.
BONES OF THE EXTREMITIES.
The Scapula in its double spine, the osseous arch formed by the confluence of the acromion with the coracoid process, and the substitution of a distinct foramen for the suprascapular notch, agrees with that of the Megatherium: but the span of the acromial arch is relatively wider, and the surface for the articulation of the clavicle is better marked. This articular surface, which is distinctly shewn upon the acromion of both the scapulæ in Pl. XX. is the more interesting, as being the only evidence of the clavicle of the Scelidothere which we at present possess; but it is enough to prove that this quadruped enjoyed all the advantages in the actions of the fore-extremity, which arise out of the additional fixation of the shoulder-joint afforded by the clavicle—a bone which the extinct Megatherioids are the largest of the mammiferous class to possess in a completely developed state. The form, position, and aspect of the glenoid cavity for the humerus closely correspond with the condition of the same part in the Megatherium. The limits of the acromial and coronoid portions of the arch were still defineable in the
present skeleton, which indicates the nonage of the individual in the unanchylosed condition of most of the epiphyses.
In regard to the presence of a clavicle in the Megalonyx M. Lund has deduced certain conclusions, which, if well founded, would be equally applicable to the present allied species, and to the great Megatherium. I am induced, therefore, to offer a few physiological observations on that bone, which appear to me to lead to a more correct interpretation of its uses and relations in the great mammiferous animals now under consideration.
When the anterior extremities in mammalia are used simply for the purpose of progressive motion on dry land, as in the Pachyderms and Ruminants, or in water, as in the Cetaceans, there is no clavicle; this bone is introduced between the sternum and acromion, in order to give firmness and fixity to the shoulder-joint when the fore-leg is to discharge some other office than that of locomotion. In these cases, however, the clavicle exists in various degrees of development, and even its rudiment may be dispensed with in some of the actions which require a considerable extent of lateral or outward motion, and of freedom of rotation of the fore-limb. When, therefore, we find the clavicle fully developed in the skeleton of an extinct mammiferous animal, and so placed as to give the humeral articulation all the benefit of this additional mechanism, we may confidently expect that it will afford an insight into the habits and mode of life of such extinct species. M. Lund* has argued from the clavicle of the Megalonyx, that it climbed like a Sloth. "Animals," says Sir C. Bell,† "which fly or dig, or climb, as Bats, Moles, Porcupines, Squirrels, Ant-eaters, Armadilloes, and Sloths, have this bone; for in them, a lateral or outward motion is required." But in regard to the present problem, we have to enquire whether the clavicle manifests any modifications of form, of strength, or development in relation to the special differences of these several actions, with which its presence is asserted to be associated?
In mammals which fly, the clavicle is always complete: the rabbit, the fox, and the badger are instances of burrowing animals in which the clavicle is absent or rudimental. The presence of a perfect clavicle is not more constant in climbing quadrupeds. The Ai, for example, has an incomplete clavicle, which is attached to the acromion process, and terminates in a point about one-fourth of the distance between the acromion and the top of the sternum, to which the clavicular style is attached by a long slender ligament: the advantage, therefore, which a perfect clavicle affords in the fixation of the shoulder-joint, is lost to this climber par excellence. Again, the Bears, which are the bulkiest quadrupeds that are gifted with the faculty of climbing, and this in so perfect a degree that the Sun-bears of the Eastern Tropics may be termed arboreal animals,—these scan-
* Loc. cit.
† Bridgewater Treatise, p. 46.
N
[page] 90 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
sorial quadrupeds are destitute of even the smallest rudiment of a clavicle, as I have ascertained by repeated careful dissection.
Since, therefore, a clavicle in any degree of development is not essential to a climbing quadruped, we must seek for some other relation and use of that remarkably strong, and perfect bone, as it exists in the Megathere, Megalonyx, and Scelidothere. The absence of 'dentes primores' or of anterior or incisive teeth in these quadrupeds at once sets aside any idea of its connection with an action of the fore extremities, very common in the mammals which possess clavicles, viz., that of carrying the food to the mouth, and holding it there to be gnawed by the teeth. Flying is of course out of the question, although our surprise would hardly be less at seeing a beast as bulky as an elephant climbing a tree, than it would be to witness it moving through the air. If now we restrict our comparison to the relations of the clavicle in that order of Mammalia to which the extinct species in question belonged, we shall see that it is most constant, strongest, and most complete in those species which make most use of their strong and long claws in displacing the earth, as the Armadilloes and Orycteropus: and, as the clavicle is incomplete in one climbing Edental, we are naturally led to conclude that its perfect development in an extinct species must have been associated with uses and relations analogous to those with which it coexists in other genera of the same order. Thus it will be seen, that, in rejecting the conclusion drawn by M. Lund from the presence of a clavicle, I concur in the opinion expressed by Dr. Buckland* that the Megatherium—and with it the Megalonyx and Scelidotherium—had the shoulder-joint strengthened by the clavicle, in reference to the office of the fore-arm, as an instrument to be employed in digging roots out of the ground. Not, however, that these gigantic quadrupeds fed on roots, but rather, as the structure of the teeth would show, on the foliage of the trees uprooted by the agency of this powerful mechanism of the fore-legs, and of the otherwise unintelligible colossal strength of the haunches, hind-legs, and tail.
The humerus presents a large convex oval head, on each side of which is a tuberosity for the implantation of the supra- and sub-scapular muscles: these tuberosities do not rise above the articular convexity, so as to restrict the movements of the shoulder-joint, as in the Horse and Ruminants, but exhibit a structure and disposition conformable to those which characterize the proximal extremity of the humerus in other mammalia which enjoy rotatory movements of the upper or fore-limb. The tuberosities are, however, relatively more developed, and give greater breadth to the proximal end of the humerus in the Scelidothere than in the Megathere. The distal end of the humerus, although mutilated, clearly indicates that it had the same characteristic breadth of the external and internal
* Bridgewater Treatise, p. 152.
condyles, as in the Megatherium. In fig. 1. Pl. XXV. which gives a front view of the left humerus, the broad internal condyle, with its extremity broken off, is seen projecting to the left hand; both in this figure and in fig. 2. in which the internal side of the humerus is turned towards the observer, the wide groove, with its two osseous boundaries, is shewn, which plainly indicates that the left condyle was perforated for the direct passage of the artery or median nerve, or of both, to the fore-arm. The groove for the musculo-spiral nerve on the outer side of the humerus is over-arched at its upper part by a strong obtuse process; which is comparatively less developed in the Megatherium. The trochlear or inferior articular surface of the humerus presents, as in the Megatherium, two well-marked convexities, with an intervening concavity: this indication of the rotatory power of the fore-leg is confirmed by the form of the head of the radius.
In Pl. XXV. fig. 4. a view is given of this articular surface: it presents the form of a subcircular gentle concavity, which plays upon the outer convexity of the humeral articular surface: immediately below the upper concavity the radius presents a lateral smooth convex surface, which rotates upon a small concavity on the ulna, analogous to the 'lesser semilunar,' in human anatomy, in which the mechanism for rotation, so far as the upper joint of the radius is concerned, is not more elaborately wrought out than in the present extinct edentate quadruped. The radius expands as it proceeds to the elbow-joint, where it attains a breadth indicative of the great power and size of the unguiculate paw, of which it may be called the stem, and to the movements of which it served as the pivot.
All the bones of the fore-limb just described—the scapula, the humerus, and the radius,—indicate by the bold features and projections of the muscular ridges and tubercles the prodigious force which was concentrated upon the actions of the fore-paw, and the ulna, in its broad and high olecranon (of which a side-view is given in fig. 2. Pl. XXV.) gives corresponding evidence. The great semilunar concavity is traversed by a sub-median smooth ridge, which plays upon the interspace of the two humeral convexities. The body of the bone is subcompressed, straight, and diminishes in size as it approaches the carpal joint: the immediate articulating surfaces are wanting in both the radius and ulna, the epiphysial distal extremities having become detached from their respective diaphyses.
Of the terminal segment of the locomotive extremities, the only evidence among the remains of the skeleton of the Scelidothere is the ungueal phalanx figured at Pl. XXVII. 3, 4, and 5; but as it is uncertain whether it belong to the fore or hind-foot, it will be described after the other bones of the extremities have been noticed.
Of these bones the femur is the most remarkable, both for its great proportional size, and its extreme breadth, as compared with its length or thick-
N 2
[page] 92 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
ness: but in all these circumstances the affinity of the Scelidothere with the Megathere is prominently brought into view. There is no other known quadruped with which the Scelidothere so closely corresponds in this respect. In proceeding, however, to compare together the thigh-bones of these two extinct quadrupeds, several differences present themselves, which are worthy of notice: of these the first is the presence in the Scelidothere of a depression for a 'ligamentum teres' on the back part of the head of the femur, near its junction with the neck of the bone: this is shewn in the posterior view of the femur given in Pl. XX. The head itself forms a pretty regular hemisphere: the great trochanter does not rise so high as in the Megatherium, but, relatively, it emulates it in breadth: the small trochanter is proportionally more developed: the external contour of the shaft of the femur is straighter in the Scelidothere than in the Megathere, and the shaft itself is less bowed forwards at that part. The articular condyles occupy a relatively smaller space upon the distal extremity of the femur in the Scelidothere, and they differ more strikingly from those of the Megathere, in being continued one into the other: the rotular surface, for example, which is shewn in fig. 5. Pl. XXV. is formed by both condyles, while in the Megatherium it is a continuation exclusively of the external articular surface.
The patella, which works upon the above-mentioned surface, is a thick strong ovate bone, with the smaller end downwards: rough and convex externally, smooth on the internal surface, which is concave in the vertical and convex in the transverse directions.
Of the bones of the leg only the proximal end of the tibia is preserved; but this is valuable, as shewing another well-marked difference between the Scelidothere and Megathere; for whereas in the latter the fibula is anchylosed with the tibia, this bone, in the Scelidothere, presents a smooth flat oval articular surface, which is shewn in fig. 2. Pl. XXVII. below the outer part of the head of the bone; from the size and appearance of which, I infer, that the fibula would not have become confluent with the tibia, even in the mature and full-grown animal.
The relative length of the fore and hind extremities cannot be precisely determined from the present imperfect skeleton of the Scelidothere; but there is good evidence for believing, that the fore extremity was the shortest. The humerus is shorter than the femur by one-ninth part of the latter bone; and the radius, which wants only the distal epiphysis, must have been shorter than the humerus. Now the relative development of the fore and hind legs is one of the points to be taken into consideration in an attempt to determine the habits and nature of an extinct mammal.
In climbing animals the prehensile power is more essential to the hinder than to the fore parts or extremities. In the leech the principal sucker is in the tail;
and higher organized climbers, in like manner, depend mainly on their posterior claspers in descending trees, and hold on by means of them whilst selecting the place for the next application of those at the fore part of the body, whether their place be supplied by the beak, as in the Maccaws, or the fore-feet or hands in the Mammalia.
But, although we perceive the hinder limbs to be the last to lose the advantageous structure of the hand in the Quadrumanous species, and notwithstanding that the tail is for this purpose sometimes specially organized to serve as a prehensile instrument, yet we find that the power of grasping the branches of trees by either legs or tail is never maintained at the expense of undue bulk and weight of those organs. On the contrary, as the fore-limbs are the main instruments in the active exertions of climbing, so they are the strongest as well as the longest in all the best climbers, and the weight of the body which they have to drag along is diminished by dwarfish proportions of the hinder limbs, as in the Orangs and the Sloths.
Can those huge quadrupeds have been destined to climb that had the pelvis and hinder extremities more ponderous and bulky in proportion to the fore-parts of the body than in any other known existing or extinct vertebrate animals?
M. Lund argues for the scansorial character of the Megalonyx, because its anterior extremities are longer than the posterior ones; but if they somewhat exceed the hind-legs in length, how vastly inferior are they in respect of their breadth and thickness. The prehensile faculty of the hinder limbs of the best climbers, as the Sloths, Orangs, and Chameleons is by no means dependent on the superior mass of muscle and bone which enters into their conformation, but is associated with the very reverse conditions.
It is impossible to survey the discrepancy of size between the femur and the humerus of the Scelidothere, as exhibited in Pl. XX., without a conviction that it relates to other habits than those of climbing trees. The expanse of the sacrum, the evidence of the muscular masses employed in working the hind legs and tail, which is afforded by the capacity of the cavity lodging the part of the spinal marrow from which the nerves of those muscles were derived, both indicate the actions of the hind-legs and tail to have been more powerful and energetic than would be required for mere prehension: and the association of hinder extremities so remarkable for their bulk, with a long and powerful tail, forbids my yielding assent to the speculation set forth by M. Lund, as to the prehensile character of the tail of the Megalonyx.
Astragalus.—In the examination of this characteristic bone I have kept in view the question of the habits of the Megatherioid quadrupeds in general, and the especial affinities of the Scelidotherium, in illustration of which I shall notice at
[page] 94 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
the same time the peculiarities of the astragalus of the Sloth, Megatherium and Armadillo.*
The upper articular surface of the astragalus of the Scelidotherium (Pl. XXVI. fig. 4.), presents, in its transverse contour, two convex pulleys, a and b, and an intermediate concavity, forming one continuous articular surface. The external or fibular trochlea (a) is strictly speaking convex only at its posterior part, the upper surface gradually narrowing to a ridge, as it advances forwards, from which, the inner and outer parts slope away at an angle of 35°.
The tibial† convexity (b) is more regular and less elevated, it has only half the antero-posterior extent of the outer pulley; its marginal contour forms an obtuse angle at the inner side.
In the Megatherium the upper articular surface of the astragalus is also divided into two trochleæ, of which the one on the fibular side (fig. 3, a), is of much greater relative size and extent than the tibial one (b), and is raised nearly four inches above the level of the latter, although in the oblique position in which the bone is naturally placed in the skeleton, the highest part of each convexity is on the same level The fibular trochlea differs also from that in the Scelidothere in being regularly convex in the transverse as well as the antero-posterior direction. The tibial convexity resembles that in the Scelidothere, save in its smaller relative size; its internal margin likewise forms an angular projection below the internal malleolus.
The upper surface of the astragalus of the Mylodon, or Megalonyx(?) (Pl. XXVIII. fig. 5.),‡ differs from that in the Megatherium in having a narrower fibular trochlear ridge.
The astragalus of the Ai (Bradypus tridactylus) differs widely from that of either the Megathere, Mylodon (?) or Scelidothere in having a conical cavity on the upper surface, in place of the fibular convexity, in which concavity the distal end of the fibula rotates like a pivot. This mechanism is closely related to the scansorial uses of the inwardly inflected foot of the Sloth.
If the astragalus of an Armadillo§ were placed side by side with that of the
* Dasypus 6-cinctus, L., is the species of which I have the astragalus separate, so as to be able to follow out the comparison.
† In distinguishing these trochleæ as fibular and tibial, it is to be understood that the terms relate only to aspects corresponding to the position of those bones, and not that the fibula is articulated to the whole of the trochlea so called: it probably rested only upon the outer facet in the Scelidothere.
‡ This astragalus was found at Santa Fé, in Entre Rios, associated with the remains of the Mastodon and Toxodon; but from its size and form I entertain little doubt that it belonged to a Megatherioid quadruped as large as the Mylodon or Megalonyx. The brief allusion to the astragalus of the Megalonyx in M. Lund's Memoir does not afford the means of determining with certainty this point.
§ See the figures of this bone, given by Cuvier in Pl. x. and xi. Ossemens Fossiles, vol. v. part i.
Megathere, it would be very difficult to determine the analogous parts, especially of the upper surface, unless guided by the intermediate structure presented by the Scelidothere. The upper surface of this bone, in the Armadillo, is, however, divided into two transversely convex trochleæ, separated by a much wider transversely concave surface. The fibular trochlea resembles that of the Scelidothere in having its upper and outer facets sloping away at an acute angle, but without meeting at a ridge anteriorly; this surface is not more raised above the tibial trochlea than in the Scelidothere.
The inner trochlea differs from that of the Scelidothere in having a greater relative antero-posterior extent, and in forming, in place of an uniform convex surface, a trochlea similar in structure to that on the outer side. The extent of rough surface on the upper part of the astragalus intervening between the articular surface for the bones of the leg, and that for the scaphoides is extremely small in the Megathere and Mylodon (?); it is relatively greater in the Scelidothere; it is still more extensive in the Armadillo; but is the longest in the Sloth. The anterior extremity of the astragalus which is entirely occupied by the scaphoid articular surface is very peculiar in the Scelidothere (Pl. XXVI. fig. 2.): it presents one convex and two concave facets, which, however, form part of one continuous articular surface: the convex facet forms the internal part of the surface, and presents a rhomboidal form with the long axis vertical. The concave facets (c and d) are extended transversely and placed one above the other; they are slightly concave in the transverse, and nearly flat in the vertical directions.
In the Megatherium (fig. 1.) the scaphoid surface of the astragalus is divided only into one concave and one convex portion, both continuous with each other: the concave facet (c) corresponds with the upper concavity in the Scelidothere, but is a pretty uniform subcircular depression, fourteen lines in depth: the convex facet, d, is continued across the whole breadth of the under part of the scaphoid surface and corresponds with both the inner convex, and lower concave surfaces of the scaphoid articulation in the Scelidothere.
In the Mylodon (?) (Pl. XXVIII. fig. 3.), the articular facet, corresponding with that marked (c) in the astragali of the Megathere and Scelidothere, is simply flattened, instead of being concave; the rest of the scaphoid surface corresponds with that in the Megatherium.
In the Armadillo the scaphoid articular surface is undivided and wholly convex: in this part of the astragalus, therefore, we find the Scelidothere deviating from the Armadillo further than does the Megathere; while the Mylodon or Megalonyx (?) most resembles the Armadillo in the configuration of this part of the astragalus.
If we compare the outer surfaces of the astragalus in these quadrupeds,
[page] 96 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
we shall find, however, that the Scelidothere and Armadillo closely agree: the outer facet of the fibular trochleæ, above described, is continued in the Scelidothere (Pl. XXVIII. fig. 2.), upon the fibular side of the astragalus reaching nearly half-way down the posterior part, and down nearly the whole of its anterior.
In the Armadillo, it extends over the whole of the anterior part of the outer side of the astragalus. In both animals the lower boundary of this articular surface describes a strong sigmoid curve.
In the Megatherium (Pl. XXVIII. fig. 1), the corresponding surface for the fibular malleolus on the outer side of the astragalus is formed by a comparatively very small semicircular flattened facet, which by its roughness indicates that the end of the fibula was attached to it by ligamentous substance, and that the synovial bag was not continued upon that surface as in the Scelidothere and Armadillo.
In the Mylodon (?) (Pl. XXVIII. fig. 4), even this rough facet is wanting and the fibular trochlea is bounded by the angle which divides the upper from the outer surface of the astragalus.
Turning now our attention to the under surface of the astragalus, we observe that it presents in the Scelidothere (Pl. XXVI. fig. 6), an irregular quadrate form, having the outer side occupied by an elongated sub-ovate articular facet, e, for the calcaneum, bounded externally by a sharp edge, with its long axis and its greatest concavity in the antero-posterior direction, and slightly convex from side to side: a second calcaneal articular surface (f) is situated at the inner and anterior angle; it is oblong and nearly flat; is continuous with the inferior concave facet of the scaphoid articulation, but is divided from the convex facet by a groove: the two calcaneal articulations are separated by a deep and rough depression, traversing the under surface of the astragalus diagonally, and increasing in breadth towards the posterior and internal angle. The inner side of the astragalus presents a convex protuberance.
The correspondence between the astragalus of the Scelidothere and Megathere is best seen at the under surface of the bone: in both the two calcaneal articulations are separated by the diagonal depression, and the internal and anterior surface is continuous with the scaphoid articulation. In the Megathere, however, in consequence of the absence of the inferior concavity which characterizes the Scelidothere, the anterior calcaneal facet (f) appears as a more direct backward continuation of the scaphoidal surface; but they are divided by a more marked angle than is represented in the figure (fig. 5, Pl. XXVI.). The posterior and outer calcaneal surface in the Megathere (e) is broader in proportion to its length, continued further upwards upon the outward surface, is consequently more convex in the transverse direction, and is not bounded externally by so sharp and prominent a ridge as in the
Scelidothere. The protuberance from the inner surface of the astragalus is more compressed laterally in the Megathere than in the Scelidothere. The correspondence between the astragali of the Mylodon (?) (Pl. XXVIII. fig. 6) and Megathere in the conformation of the under surface is so close, that the few differences which exist will be sufficiently appreciated by an inspection of the figures.
In the Armadillo the astragalus, in consequence of the greater production of its anterior part, presents more of an angular than a quadrate figure; and the scaphoid articular surface, being proportionally carried forwards, is altogether separated from the anterior calcaneal surface. The posterior and inner calcaneal surface resembles that in the Scelidothere, but is less inclined upwards; and is continuous with the posterior part of the tibial articular surface.
Thus the astragalus in the structure of its two most important articulations, viz. that which receives the superincumbent weight from the leg, and that which transmits it to the heel, presents a closer correspondence in the Scelidothere with that of the Dasypus, than with that of the Megathere or Mylodon.
The ungueal phalanx of the Scelidothere before alluded to, is represented of the natural size in Pl. XXVII. The side-view, fig. 3. shows the position of the articular surface on the proximal end, sloping obliquely towards the under surface, and overtopped by an obtuse protuberance, calculated to impede any upward retraction of the claw: the present joint, in fact, illustrates in every particular the argument by which Cuvier established the true affinities of the allied extinct genus Megalonyx.*
The present phalanx is, however, less compressed, and less incurved than those of the Megalonyx, which have been hitherto described; but it more resembles in these proportions one of the smaller, and presumed hinder, ungueal phalanges of the Megatherium. The upper and lateral parts of the bone are rounded, and it gradually tapers to the apex, which is broken off. The osseous sheath for the claw is developed only at the under part of the bone: it presents the form of a thick flat plate of bone, with the margin very regularly and obliquely bevelled off, and having a vertical process of bone attached lengthwise to the middle of its under surface This process must have served for the insertion of a very powerful flexor tendon. The figures of this bone preclude the necessity of any further verbal description.
M. Lund lays most stress upon the argument founded on the inward inflection of the sole of the foot in the Megalonyx, and appeals with greatest confidence to this structure in support of his hypothesis of the scansorial habits of that extinct Edental.†
* Ossemens Fossiles, vol. v. part i. p. 163.
† For the translation of the following passage, and of others alluded to in the present work, from the original Danish Memoir of M. Lund, loc. cit., I am much indebted to the Rev. W. Bilton, M.A. &c. &c.:—
O
[page] 98 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
It is quite true that the Quadrumana derive advantage from this position of the foot in climbing trees, and that it is carried to excess in the Sloths, which can only apply the outer edge of the foot to the ground. But we may ask, was the inversion of the sole of the foot actually carried to such an extent in the Megalonyx? And, admitting its existence in an inferior degree, is it then conclusive as to the scansorial habits of that species?
M. Lund expressly states that it is produced by a different structure and arrangement of the tarsal bones, from that which exists in the Sloth, but he does not specify the nature of this difference.
If the astragalus, which I have referred with doubt to the Megalonyx, do not actually belong to that genus, it is evidently part of a very closely allied species. Now this astragalus, as we have seen, resembles most closely that of the Megatherium; and since we may infer that the calcaneum, scaphoides, and cuboides had a like correspondence, the inclination of the sole of the foot inwards must have been very slight, as I have determined from examination of the structure and co-adaptation of those bones in the incomplete skeleton of the Megatherium in the London College of Surgeons. Such an inclination of the foot may be conceived to have facilitated the bending of the long claws upon the sole, during the ordinary progressive movements of the animal, but it is quite insufficient to justify the conclusion, that it related to an application of the hind feet for the purposes of climbing.
It is not without interest again to call to mind the deviation of the structure
"Thus in every point of comparison we have instituted between the organization of burrowers and climbers; we have seen that the Megalonyx constantly differs from the former and resembles the latter; but the point to which I last alluded (the obliquity of foot), I consider to be quite decisive.
"There is one other point in its organization, which is not quite without weight in reference to our present inquiry,—I mean its unusually powerful tail. Now, it is certainly true that many animals which are not climbers have a powerful tail, as e. g. Armadillos, while the others that climb well, have none, as Sloths and Apes. But when we find a remarkably powerful tail attached to an animal that according to all probability was a climber, we are led to infer that this organ must have served for that purpose: in other words, that the Megalonyx was furnished with a prehensile tail.
"How far the Megatherium is to be considered in the same light as the Megalonyx cannot be decided without an accurate and scientific examination of its skeleton at Madrid. Pander and D'Alton do not mention any distortion of the hind-foot, neither does their figure exhibit any. It is nevertheless quite possible that such may exist, but that it is disguised by the faulty manner in which the skeleton is put up. It strikes me as little probable that two animals which agree so well in the principal particulars of their organization should differ so much in one of the most important. The Megatherium has been proved by later discoveries to possess the same powerful tail as the Megalonyx, and as it corresponds also with the latter entirely in the conformation of its extremities, the same difficulties present themselves against the supposition of its having been a burrower. But if the Megatherium was really a climber, it must have had still more occasion (on account of its greater size), for that peculiar arrangement of the hind-feet which we have described in the Megalonyx."
of the astragalus of the Scelidothere from the Megatherioid to the Dasypodoid type of structure. For if the Megatherioid type of structure had really been one suitable to the exigencies of climbing quadrupeds, it might have been expected to have exhibited the scansorial modifications more decidedly, as the species diminished in stature; but as regards the instructive bone of the hind-foot, the modifications of which we have just been considering, this is by no means the case.
DESCRIPTION OF A MUTILATED LOWER JAW OF THE
MEGALONYX JEFFERSONII.
In the preceding section an astralagus was described, which was regarded as belonging possibly to the same Edentate species as the jaw figured and described, p. 69, Pl. XVIII. and XIX., under the name of Mylodon Darwinii; but the same correspondence,—that of relative size,—renders it equally possible that this astragalus may belong to the species of Megalonyx to which the lower jaw now under consideration appertains. There could be no doubt, from its structure, that it was the astragalus of a gigantic species of the order Bruta, and of the Megatherioid family, and more nearly allied to the Megathere than is the Scelidothere, but sufficiently distinct from both.
The lower jaw, figured in Pl. XXIX., is the only fossil brought home by Mr. Darwin that could be confidently referred to the genus Megalonyx; but the form of the tooth in place on the right side of the jaw fully justifies this determination. The jaw itself is deeply and firmly imbedded in the matrix, so that only the upper or alveolar border is visible. The coronoid and condyloid processes are broken away, and the texture of the remaining part of the jaw was too friable, and adhered too firmly to the surrounding matrix to admit of more of its form being ascertained than is figured.
There were four molars on each side of this jaw; the large oblique perforation near the fractured symphysis is the anterior extremity of the wide dental canal. The forms of the alveoli are best preserved in the right ramus: the first is the smallest, and seems to have contained a tooth, of which the transverse section must have been simply elliptical: the second tooth is likewise laterally compressed, but the transverse section is ovate, the great end being turned forwards: the third socket presents a corresponding form, but a larger size: the fourth socket is too much mutilated to allow of a correct opinion being formed as to the shape of the tooth which it once contained. The natural size of the tooth
O 2
[page] 100 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
in situ, and of the adjoining socket, is given in Pl. XXIX., fig. 2. The difference of form which the jaw of the Megalonyx presents, as compared with that of the Mylodon, especially in the greater recedence of the two horizontal rami from each other, will be appreciated by comparing Pl. XVIII. with Pl. XXIX.
DESCRIPTION OF A FRAGMENT OF THE SKULL AND OF THE
TEETH OF THE
MEGATHERIUM CUVIERI.
NOTWITHSTANDING the full, accurate, and elaborate accounts of the skeleton of the Megatherium given by Brû,* Cuvier,† Pander and D'Alton,‡; and Mr. Clift,§ the fragments of this most gigantic of quadrupeds brought home by Mr. Darwin, possess much interest, and have added, what could hardly have been anticipated, important information as to the dental system, whereby an error in the generic character of the Megatherium has been corrected.
The fragments here alluded to are portions of the skull of three full-grown Megatheres: the most perfect part of which affords a view of the posterior, and of part of the basal surface, which regions of the cranium have not hitherto been elsewhere figured or described, (Pl. XXX.)
The plane of the occipital foramen forms with that of the base of the skull an angle of 140°, the plane of the posterior surface of the skull forms with the basal plane an angle of 68°. The occipital condyles are therefore terminal, or form the most posterior parts of the cranium. The extent of their convex curvature in the antero-posterior direction, which equals that of a semicircle, indicates that the Megatherium possessed considerable freedom and extent of motion of the head. The condyles are not extended in the lateral direction so far as in the Toxodon; their axis is more oblique than in the Glossotherium, and their internal surface is more parallel with the axis of the skull, the foramen magnum not presenting that infundibuliform expansion which is so characteristic of the Glossotherium. The occipital condyles resemble most in form and position those of the Scelidotherium; but in the angle of the occipital plane the Megatherium is intermediate between the Scelidothere and Glossothere. The ex-occipitals terminate laterally and inferiorly, each in a short, but strong obtuse process. The posterior plane of the skull is traversed by a strong arched intermuscular crest, which
* Descripcion del Esqueleto de un quadrupedo muy corpulento y raro, que se conserva en el Real Gabinete de Historia Natural de Madrid. Folio, Madrid, 1796.
† Ossemens Fossiles, tom. v. pt. i. p. 179.
‡ Das Riesen Faulthier, Bradypus giganteus, von Dr. Chr. Pander und Dr. E. D'Alton." Folio, Bonn, 1821.
§ Transactions of the Geological Society, 1835, p. 438.
forms the upper boundary of a pretty deep fossa, which is divided by a median vertical ridge, extending downwards to within an inch of the upper margin of the foramen magnum. A second strong obtuse transversely arched ridge curves over the first, and forms the upper boundary of the posterior or occipital region of the skull: the interspace between the two transverse ridges is very irregular, and indicates the firm implantation of powerful nuchal muscles or ligaments, (Pl. XXX. fig. 1.)
In the configuration and angle of the occipital plane the Megatherium indicates the same general correspondence with the Edentate type, which has been pointed out in the descriptions of the crania of the Glossothere and Scelidothere: and the resemblance to the Scelidothere is not less striking in the small proportional size of the cranium in this quadruped, which surpasses the rest of its class in so great a degree in the colossal proportions of its hinder parts.
Having detected in the base of the skull of the Scelidothere an articular semicircular pit for the head of the styloglossal bone, similar to, but relatively smaller than, that remarkable one in the skull of the Glossothere, it became a matter of interest to determine whether this structure, which does not exist in any of the existing Edentals, should likewise be present in the gigantic type of the Megatherioid family. The result of a careful removal of the matrix from the basal region of one of the cranial fragments of the Megatherium was the detection of this articular cavity, in each temporal bone in the same relative position as in the Glossothere and Scelidothere. The styloid articular cavity is relatively smaller, and shallower, than in the Glossothere, its proportions being much the same as those of the Scelidothere. The cranial or posterior extremity of the stylo-hyoid bone in the Scelidotherium is bent upwards at an obtuse angle (Pl. XXI.), and terminates in an articular ball which rotates in this cavity. The size of this bone, and its mode of articulation, indicates great power and muscularity of tongue in the Megatherioids, and calls to mind the importance of that organ in the Giraffe, which subsists on the same kind of food as that which I have supposed to have supported the Megatherioids, although the general organization of these animals and the mode in which the foliage was brought within reach of the tongue are as opposite as can well be imagined.
The anterior condyloid foramen presents scarcely one half the absolute size of that of the Glossothere, whence we may infer a correspondingly inferior development of the tongue in the Megathere. The fractured parietes of the cranial cavity of the Megatherium every where exhibit evidences of the great extent of the air-cells or sinuses continued from the nasal cavity: on the basilar aspect of the cranium they extend as far back as the jugular foramina: the whole of the basi-sphenoid being thus excavated, and permeable
[page] 102 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
to air, derived from the sphenoid sinuses, (Pl. XXX. fig. 2.) The vertical diameter of the cranial cavity is four inches, eight lines; its transverse diameter, which is greatest in the posterior third part of the cavity, corresponding with the posterior part of the cerebrum is six inches: from the indications afforded by the remains of the cranial cavity in Mr. Darwin's specimens, I conclude that the brain of the Megatherium was more depressed, and upon the whole, smaller by nearly one-half than that of the Elephant; but with the cerebellum relatively larger, and situated more posteriorly with relation to the cerebral hemispheres: whence it may be concluded that the Megatherium was a creature of less intelligence, and with the command of fewer resources, or a less varied instinct than the Elephant.
It has been usual to characterize the Megatherium, in conformity with the concurrent descriptions of Bru, Cuvier, and D'Alton, by the dental formula of molares 4/4 4/4, i. e. by the presence of four grinding teeth on each side of the upper, as of the lower jaw. It was the agreement of the excellent authorities above cited in this statement, which induced Mr. Clift and myself to regard a single detached tooth, which formed part of the valuable collection of remains of the Megatherium deposited in the Hunterian Museum by Sir Woodbine Parish, as being, from its comparatively small size, the tooth of either a younger individual or of a smaller species of Megatherium. Upon clearing away the matrix from the palatal and alveolar surface of one of the cranial fragments of the Megatherium in Mr. Darwin's collection, I was gratified by the detection of the crown of a fifth molar, corresponding in size and form with the detached tooth, above alluded to: its small size, and its position have doubtless occasioned its being over-looked in the cranium of the great skeleton at Madrid.
The anterior molar of the upper jaw presents a nearly semicircular transverse section, with the angles rounded off; the three succeeding teeth are four-sided, with the transverse somewhat exceeding the antero-posterior diameter: they are rather longer and larger than the first: the last molar is likewise four-sided, but presents a sudden diminution of diameter, and is relatively broader. The following are the respective dimensions of the upper maxillary teeth.
| First Molar. | Second Molar. | Third Molar. | Fourth Molar. | Fifth Molar. | ||||||
| In. | Lines. | In. | Lines. | In. | Lines. | In. | Lines. | In. | Lines. | |
| Length . . . . . | 8 | 6 | 9 | 4 | 9 | 4 | 8 | 7 | 5 | 2 |
| Transverse diameter . . . | 1 | 9 | 2 | 4 | 2 | 3 | 2 | 0 | 1 | 4 |
| Antero-posterior diameter . . | 1 | 5 | 2 | 0 | 2 | 0 | 1 | 11 | 0 | 10 |
Besides the differences in size, the upper molars vary as to their curvature: this difference is exhibited in the vertical section of these teeth figured in Pl. XXXI. The convexity of the curve of the first, second and third molars is directed
forwards; the fourth is straight, its anterior surface only describing a slight convexity in the vertical direction; the fifth tooth is curved, but in a contrary direction to the others; and the bases of the five molars thus present a general convergence towards a point a little way behind the middle of the series.
The next peculiarity to be noticed in these remarkable teeth is the great length of the pulp-cavity (d), the apex of which is parallel with the alveolar margin of the jaw: a transverse fissure is continued from this apex to the middle concavity of the working surface of the tooth, which is thus divided into two parts. Each of these parts consists of three distinct substances,—a central part analogous to the body or bone of the tooth or 'dentine,' a peripheral and nearly equally thick layer of cæmentum, and an intermediate thinner stratum of a denser substance, which is described in Mr. Clift's memoir on the Megatherium as 'enamel,' and to which substance in the compound teeth of the Elephant, it is analogous both in its relative situation, and relative density to the other constituents.
Microscopic examinations of thin and transparent slices of the tooth of the Megatherium prove, however, that the dense layer separating the internal substance from the cæmentum is not enamel, but presents the same structure as the hard 'dentine' or ivory of the generality of Mammalian teeth; and corresponds with the thin cylinder of hard 'dentine' in the tooth of the Sloth. No species of the Order Bruta has true enamel entering into the composition of its teeth; but the modifications of structure which the teeth present in the different genera of this order are considerable, and their complexity is not less than that of the enamelled teeth of the Herbivorous Pachyderms and Ruminantia, in consequence of the introduction of a dental substance into their composition corresponding in structure with that of the teeth of the Myliobates, Psammodus, and other cartilaginous fishes.
The microscopic investigation of the structure of the teeth of the Megatherium was undertaken chiefly with the view of comparing this structure with that of the teeth of the Sloth and Armadillo, and of thus obtaining an insight into the food, and an additional test of the real nature of the disputed affinities of the Megatherium. The central part of the tooth (c. Pl. XXXI.) consists of a coarse ivory, like the corresponding part of the tooth of the Sloth. It is traversed throughout by medullary canals 1/1500th of an inch in diameter, which are continued from the pulp-cavity, and proceed, at an angle of 50°, to the plane of the dense ivory, parallel to each other, with a slightly undulating course, having regular interspaces, equal to one and a half diameters of their own arcæ, and generally anastomosing in pairs by a loop of which the convexity is turned towards the origin of the tubes of the fine dentine, as if each pair so joined consisted of a continuous reflected canal, (c. fig. 1, Pl. XXXII.) The loops are gene-
[page] 104 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
rally formed close to the fine dentine. In a few situations I have observed one of the medullary canals continued across the fine dentine, and anastomosing with the corresponding canals of the cæmentum. The interspaces of the medullary canals of the coarse dentine are principally occupied by calcigerous tubes which have an irregular course, anastomose reticularly, and terminate in very fine cells. The more regular and parallel calcigerous tubes, which constitute the thin layer of hard dentine, are given off from the convexity of the terminal loops of the medullary canals. The course of these tubes (b. fig. 1, Pl. XXXII.) is rather more transversely to the axis of the tooth than the medullary canals from which they are continued. They run parallel to each other, but with minute undulations throughout their course, in which they are separated by interspaces equal to one and a half their own diameter. As they approach the cæmentum they divide and sub-divide, and grow more wavy and irregular: their terminal branches take on a bent direction, and form anastomoses, dilate into small cells, and many are seen to become continuous with the radiating fibres or tubes of the cells or corpuscles of the contiguous cæmentum. This substance enters largely into the constitution of the compound tooth of the Megatherium: it is characterized, like the cæmentum of the Elephant's grinder, by the presence of numerous radiated cells, or purkingian corpuscles, scattered throughout its substance, but may be distinguished by wide medullary canals which traverse it in a direction parallel with each other, and forming a slight angle with the transverse axis of the tooth. These canals are wider than those of the central coarse dentine, their diameter being 1/1200th of an inch; they are separated by interspaces equal to from four to six of their own diameters, divide a few times dichotomously in their course, and finally anastomose in loops, the convexity of which is directed towards, and in most cases is in close contiguity with, the layer of dense dentine.
Fine calcigerous tubes are every where given off at right angles from the medullary canals of the cæmentum, which form a rich reticulation in their interspaces, and a direct continuation between the loops of the medullary canals and the calcigerous tubes of the dense dentine. The cæmentum differs from the coarse dentine in the larger size and wider interspaces of its medullary canals, and by the presence of the bone-corpuscles in their interspaces; but they are brought into organic communication with each other, not only by means of the tubes of the dense dentine, but by occasional continuity of the medullary canals across that substance. The tooth of the Megatherium thus offers an unequivocal example of a course of nutriment from the dentine to the cæmentum, and reciprocally. Retzius observes with respect to the human tooth, that "the fine tubes of the cæmentum enter into immediate communications with the cells and tubes of the dentine (zahnknochen), so that this part can obtain from without the requi-
site humours after the central pulp has almost ceased to exist." In the Megatherium, however, those anastomoses have not to perform a vicarious office, since the pulp maintains its full size and functional activity during the whole period of the animal's existence. It relates to the higher organized condition, and greater degree of vitality of the entire grinder in that extinct species.
The conical cavities (d. Pl. XXXI.) attest the size and form of the persistent pulp; the diameter of its base is equal to the part of the crown of the tooth which is formed by the coarse and fine dentine. From the gradual thinning off, and final disappearance of these substances as they reach the base of the tooth, I conclude that they were both formed at the expense of the pulp. The fine tubes and cells must have been excavated in its peripheral layer for the reception of the hardening salts of the dense dentine, and the rest converted into the parallel series of medullary canals with their respective systems of calcigerous tubes, in a manner closely analogous to the development of the entire tooth of the Orycteropus. The coarser dentine of the tooth of the Megatherium differs, in fact, from the entire tooth of the Orycteropus, only in that the parallel medullary canals and their radiating calcigerous tubes are not separated from the contiguous canals by a distinct layer of cæmentum, and that the medullary canals anastomose at their peripheral extremities. The wide spaces, (e. Pl. XXXI.) indicate the thickness of the dental capsule by the ossification of which the exterior stratum of cement was formed. It was not until I knew the true structure of the tooth of the Megatherium, that I could comprehend the mode of its formation. The parallel layers of enamel in the Elephant's grinder are formed, as is well known, by membranous plates passing from the coronal end of the closed capsule towards the base of the tooth; but a certain extent of enamel can only thus be formed, and when the crown of the grinder has once protruded, and come into use, the enamel cannot be added to. The modification of the structure of the tooth of the Megatherium readily permits the uninterrupted and continuous formation of the dense substance which is analogous to the enamel of the Elephant's grinder.
With respect to the question of the respective affinities of the Megatherium to the Bradypodoid or Dasypodoid families, the result of this examination of the teeth speaks strongly for its closer relationship with the former group: the Megalonyx, Mylodon, and Scelidotherium, in like manner correspond in the structure of their teeth with the Sloth, and differ from the Armadillo.
If from a similarity of dental structure we may predicate a similarity of food, it may reasonably be conjectured that the leaves and soft succulent sprouts of trees may have been the staple diet of the Megatherioid quadrupeds, as of the existing Sloths. Their enormous claws, I conclude, from the fossorial character of the powerful mechanism by which they were worked, to have been employed,
P
[page] 106 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
not, as in the Sloths, to carry the animal to the food, but to bring the food within the reach of the animal, by uprooting the trees on which it grew.
In the remains of the Megatherium we have evidence of the frame-work of a quadruped equal to the task of undermining and hawling down the largest members of a tropical forest. In the latter operation it is obvious that the immediate application of the anterior extremities to the trunk of the tree would demand a corresponding fulcrum, to be effectual, and it is the necessity for an adequate basis of support and resistance to such an application of the fore-extremities which gives the explanation to the anomalous development of the pelvis, tail, and hinder extremities in the Megatherioid quadrupeds. No wonder, therefore, that their type of structure is so peculiar; for where shall we now find quadrupeds equal, like them, to the habitual task of uprooting trees for food ?
DESCRIPTION OF FRAGMENTS OF BONES, AND OF OSSEOUS
TESSELATED DERMAL
COVERING OF LARGE EDENTATA.
It is now determined that there once existed in South America, besides the Megatherium, the Megalonyx, and the allied genera described in the preceding pages of the present work, gigantic species of the order Bruta belonging to the Armadillo family, and defended, like the small existing representatives of that family, by a tesselated bony dermal covering. The largest known species of these extinct Dasypodidœ is the Glyptodon clavipes, of which the armour and parts of the skeleton have been described by MM. Weiss and D'Alton in the Berlin Transactions for 1827 and 1834: and the generic and specific characters and name, with an account of the dental system, and bones of the extremities, were recorded in the Geological Proceedings for March 1839. It would seem that parts of the same, or a nearly allied gigantic species were described in the same year by M. Lund; under the name of Hoplophorus. Of the valuable and interesting discoveries of this able Naturalist I regret that I was not aware until the appearance of a notice of them in the Comptes Rendus for April, 1839.* Amongst the fragments of bony tesselated armour in Mr. Darwin's collection are a few pieces which were found by him, associated with remains of Toxodon and Glossotherium near the Rio Negro in Banda Oriental.† These fragments, if we may judge from their thickness, must have belonged to an animal at least as
* An excellent translation of the description of the Brazilian fossils found by M. Lund, is published in the Annals of Natural History, July and August, 1839.
† At the distance of a few leagues from the locality here mentioned, other fragments were found by Mr. Darwin; also near Santa Fé, in Entre Rios; also on the shores of the Laguna, near the Guardia del Monte, South of Buenos Ayres; also, according to the Jesuit Falkner, on the banks of the Tercero.
large as the Glyptodon clavipes; but the pattern differs in the greater equality of size of the component tesseræ. The thickness of the largest fragment is one inch and a half, the tesseræ vary in diameter from one inch to half an inch, and are separated by grooves about two lines in depth, and two in diameter. The pattern formed by the anastomosis of these grooves is an irregular net-work; the contour of the tesseræ is either unevenly subcircular, hexagonal, pentagonal, or even four-sided; with the sides more or less unequal. In those portions of this armour, where one of the tesseræ exceeds the contiguous ones in size, the imagination may readily conceive it to be the centre of a rosette, around which the smaller ones arrange themselves, but there is no regular system of rosettes, as in the portions of the dermal armour of the Glyptodon figured by Weiss, and those brought to England by Sir Woodbine Parish, in which the central piece is double the size of the marginal ones.
The portions of the tesselated bony dermal covering of a Dasypodoid quadruped, figured in Pl. XXXII. figs. 5 and 4, of the natural size, were discovered folded round the middle and ungueal phalanges, figs. 2 and 3, at Punta Alta, in Bahia Blanca, in an earthy bed interstratified with the conglomerate containing the remains of the fossil Edentals.
In one of these fragments, measuring six inches long by five broad, the tesseræ are arranged in rosettes, and so closely correspond in size and pattern with the bony armour described by M. Lund, as characterizing his species, Hoplophorus euphractus, that I feel no hesitation in referring them to that animal. One of the pattern rosettes is figured at fig. 4, together with the thickness of the armour at this part, and the coarse tubulo-cellular structure of the bone. Another portion of dermal armour from the same locality, gives the pattern shown in fig. 5, formed by square or pentagonal tesseræ, arranged in transverse rows; it is certain that this portion of armour belonged to the same animal as the preceding piece; and probably that it constituted part of the transverse dorsal bands of the Hoplophorus.
The middle and ungueal phalanx, as well as the portions of armour, are given of the natural size, in Pl. XXXII. The upper and outer surface of the phalanx, is shown in fig. 2. It is smooth and flat; joins the inner surface by a sharp edge, which runs along the upper and inner side of the bone; and passes by a gradual convexity to the under surface; the ridge corresponding with the base of the claw, is feebly developed at the under and lateral parts of the base of the claw. Below the double trochlear joint for the middle phalanx, there are two articular surfaces for two large sesamoid bones.
The middle phalanx corresponds in its small antero-posterior diameter and wedge-shape, with that of the great Glyptodon: but the terminal phalanx is longer and deeper, in proportion to its breadth.
P 2
[page] 108 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
Among the collection of fossils from Punta Alta, in Bahia Blanca, there is an interesting fragment of the head of a gigantic animal of the Edentate order, including the glenoid cavity, and part of the zygomatic process of the left side. The articular surface for the lower jaw, exhibits, in its flatness, extent, and the absence of a posterior ridge, the well-marked characteristics of this part of the Edental structure. It measures two inches four lines in the transverse, and two inches two lines in the antero-posterior diameter. The commencement of the zygomatic process presents a vertical diameter of two inches, and a transverse diameter of eight lines at the thickest part. It is slightly concave at its lower border, and convex above. The small portion of the cranial parietes, which is preserved, exhibits the cellular structure consequent upon the great extension and development of the nasal air-sinuses: this condition of the cranial parietes, has already been noticed in the description of the more perfect skulls of the large extinct Edentata.
————————————
NOTICE OF FRAGMENTS OF MOLAR TEETH OF A
MASTODON.
Of the remains of this gigantic extinct Pachyderm, observed by Mr. Darwin at Santa Fé, in Entre Rios, and on the banks of the Tercero, the fragments of the teeth and portions of the skeleton which reached England, are not sufficient to lead to a determination of the species; but sufficiently prove it to have been nearly allied, if not identical, with the Mastodon angustidens of Cuvier, and unquestionably distinct from the Mastodon giganteum of the United States.
————————————
NOTICE OF THE REMAINS OF A SPECIES OF
EQUUS,
Found associated with the extinct Edentals and Toxodon at Punta Alta, in Bahia Blanca, and with the Mastodon and Toxodon at Santa Fé, in Entre Rios.
The first of these remains is a superior molar tooth of the right side; it was embedded in the quartz shingle, formed of pebbles strongly cemented together with calcareous matter, which adhered as closely to the tooth in question, as the corresponding matrix did to the associated fossil remains. The tooth was as completely fossilized as the remains of the Mylodon, Megatherium, and Scelidothere; and was so far decomposed, that in the attempt to detach the adherent matrix, it
became partially resolved into its component curved lamellæ. Every point of comparison that could be established proved it to differ from the tooth of the common Equus Caballus only in a slight inferiority of size.
The second evidence of the co-existence of the horse with the extinct Mammals of the tertiary epoch of South America reposes on a more perfect tooth, likewise of the upper jaw, from the red argillaceous earth of the Pampas at Bajada de Santa Fé, in the Province of Entre Rios.*
This tooth is figured at Pl. XXXII. fig. 13 and 14, from which the anatomist can judge of its close correspondence with a middle molar of the left side of the upper jaw.
This tooth agreed so closely in colour and condition with the remains of the Mastodon and Toxodon, from the same locality, that I have no doubt respecting the contemporaneous existence of the individual horse, of which it once formed part.
This evidence of the former existence of a genus, which, as regards South America, had become extinct, and has a second time been introduced into that Continent, is not one of the least interesting fruits of Mr. Darwin's palæontological discoveries.
DESCRIPTION OF REMAINS OF RODENTIA, INCLUDING THE
JAWS AND TEETH OF
AN EXTINCT SPECIES OF
CTENOMYS.
The fragment of the upper jaw, figured in Pl. XXXII. fig. 6, exhibits the first and second molar in situ, and the socket of the third and fourth molar, of a Rodent, which by the form and number of the upper maxillary teeth is referable to the genus Ctenomys. The molars are a little larger, the longitudinal groove on their external surface is somewhat deeper, and the last molar is relatively wider than in the existing subterraneous species,—the Tucutucu (Ctenomys Brasiliensis, Bl.), of whose habits so interesting an account is given in the description of the Mammalia of the present Collection (No. IV. p. 79). The form of the grinding surface of the first and second upper molar is shown below the fig. 6, and three views of the second grinder are given at figs. 7, 8, and 9. The fragment of the lower jaw of the same fossil Rodent is figured at fig. 10 and 11. The long anterior incisor is relatively narrower than in the Ctenomys Brasiliensis. I have not had the means of comparing this fossil with the Ctenomys Magellanicus; but since it is probable that the Ct. Magellanicus may not be specifically different from the Ct.
* Mr. Darwin has more particularly described the circumstances of the embedment of this tooth in his Journal of Researches, p. 149, during the Voyage of the Beagle.
[page] 110 ZOOLOGY OF THE VOYAGE OF THE BEAGLE.
Brasiliensis, it may be concluded that the present fossil is equally distinct from both.
The portion of the right hind-foot of the Rodent figured at fig. 12, includes the calcaneum, astragalus, cuboides, external and middle cuneiform bones, and the metatarsals and proximal phalanges of the toes corresponding with the three middle toes of five-toed quadrupeds. The metatarsals are chiefly remarkable for the well-developed double-trochlear articular surface, and intermediate ridge. These remains, as well as the jaws and teeth of the Ctenomys, were discovered at Monte Hermoso in Bahia Blanca.
In the same reddish earthy stratum of that locality, Mr. Darwin discovered the decomposed molar of a Rodent, equalling in size, and closely resembling in the disposition of its oblique component laminæ, the hinder molar of the Capybara (Hydrochærus.) The fossil differs, however, in the greater relative breadth of the component laminæ.
I have, lastly, to notice the head of a femur, and some fragments of pelvic bones from the same formation which bear the same proportion to the tooth above alluded to, as subsists between the teeth and bones of the Capybara, and which are sufficient to prove that there once has existed in South America a species of the family Caviidæ, as large as the present Capybara, but now apparently extinct.
This fact, together with the greater part of those which have been recorded in the foregoing pages of the present work, establishes the correspondence, in regard to the characteristic type, which exists between the present and extinct animals of the South American Continent: we have abundant evidence likewise of the greater number of generic and specific modifications of these fundamental types which the animals of a former epoch exhibited, and also of the vastly superior size which some of the species attained.
At the same time it has been shewn that some of the present laws of the geographical distribution of animals would not have been applicable to South America, at the period when the Megatherioids, Toxodon, and Macrauchenia existed: since the Horse, and according to M. Lund, the Antelope and the Hyæna, were then associated with those more strictly South American forms. The Horse, which, as regards the American continent, had once become extinct, has again been introduced, and now ranges in countless troops over the pampas and savannahs of the new world. If the small Opossums of South America had been in like manner imported into Europe, and were now established like the Squirrels and Dormice in the forests of France, an analogous case would exist to that of the Horse in South America, as the fossil Didelphys of Montmartre proves.
————————————
With respect to the geological contemporaneity of the fossils collected by him, Mr. Darwin subjoins the following observations:—
"The remains of the following animals were embedded together at Punta Alta in Bahia Blanca:—The Megatherium Cuvierii, Megalonyx Jeffersonii, Mylodon Darwinii, Scelidotherium leptocephalum, Toxodon Platensis (?) a Horse and a small Dasypodoid quadruped, mentioned p. 107; at St. Fé in Entre Rios, a Horse, a Mastodon, Toxodon Platensis, and some large animal with a tesselated osseous dermal covering; on the banks of the Tercero the Mastodon, Toxodon, and, according to the Jesuit Falkner, some animal with the same kind of covering; near the Rio Negro in Banda Oriental, the Toxodon Platensis, Glossotherium, and some animal with the same kind of covering. To these two latter animals the Glyptodon clavipes, described by Mr. Owen in the Geological Transactions, may, from the locality where it was discovered, and from the similarity of the deposit which covers the greater part of Banda Oriental, almost certainly be added, as having been contemporaneous. From nearly the same reasons, it is probable that the Rodents found at Monte Hermoso in Bahia Blanca, co-existed with the several gigantic mammifers from Punta Alta. I have, also, shown in the Introduction, that the Macrauchenia Patachonica, must have been coeval, or nearly so, with the last mentioned animals. Although we have no evidence of the geological age of the deposits in some of the localities just specified, yet from the presence of the same fossil mammifers in others, of the age of which we have fair means of judging, (in relation to the usual standard of comparison, of the amount of change in the specific forms of the invertebrate inhabitants of the sea,) we may safely infer that most of the animals described in this volume, and likewise the Glyptodon, were strictly contemporaneous, and that all lived at about the same very recent period in the earth's history. Moreover, as some of the fossil animals, discovered in such extraordinary numbers by M. Lund in the caves of Brazil, are identical or closely related with some of those, which lately lived together in La Plata and Patagonia, a certain degree of light is thus thrown on the antiquity of the ancient Fauna of Brazil, which otherwise would have been left involved in complete darkness."
Pl: XXVIII.

Lithog. from Nat. by G. Scharf. Printed by C.
Hallmandel.
Left Astragalus.
Fig. 1. Megatherium. 1/3
Nat. Size. Fig. 2. Scelidotherium. 2/3
Nat. Size. Fig. 3-6. Mylodon. ? 2/3
Nat. Size.
Pl. XXIX.

Lithog. from Nat. by G. Scharf. Printed by C. Hallmandel.
Lower Jaw of Megalonyx.
Fig: 1. 2/3 Fig: 2. Nat: Size.
Pl. XXX.

Lithog. from Nat. by G. Scharf. Printed by C. Hallmandel.
Megatherium. 1/2. Nat: Size.
Pl. XXXI.
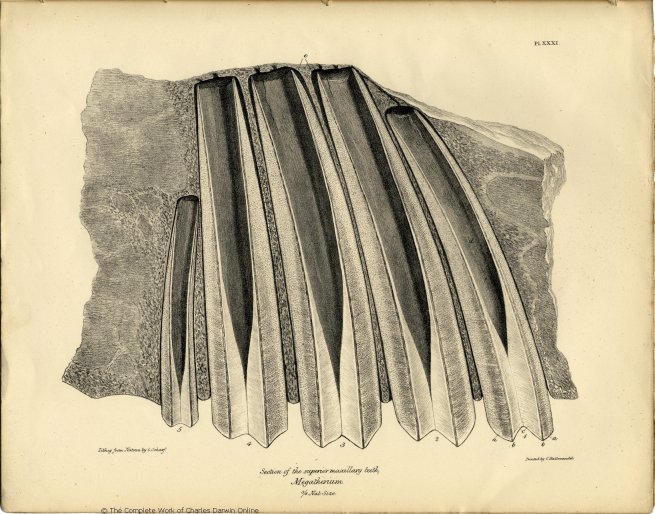
G. Scharf del et lithog. Printed by C. Hallmandel.
Section of the superior maxillary teeth,@@@
Pl. XXXII.
Lithog. from Nat: by G. Scharf. Printed by C. Hallmandel.
1. Megatherium. 2-5 Hoplophorus. 6-12. Ctenomys. 13-14. Equus.
THE
ZOOLOGY
OF
THE VOYAGE OF H.M.S. BEAGLE,
UNDER THE COMMAND OF CAPTAIN FITZROY, R.N.,
DURING THE YEARS
1832 TO 1836.
PUBLISHED WITH THE APPROVAL OF
THE LORDS
COMMISSIONERS OF HER MAJESTY'S TREASURY.
Edited and Superintended by
CHARLES DARWIN, ESQ. M.A. F.R.S. SEC. G.S.
NATURALIST TO THE EXPEDITION.
~~~~~~~
PART I.
FOSSIL MAMMALIA:
BY
RICHARD OWEN, ESQ. F.R.S.
PROFESSOR OF ANATOMY AND PHYSIOLOGY TO THE ROYAL COLLEGE OF SURGEONS IN LONDON; CORRESPONDING MEMBER OF THE INSTITUTE OF FRANCE, ETC. ETC.
~~~~~~~
LONDON:
PUBLISHED BY SMITH, ELDER AND CO. 65, CORNHILL.
MDCCCXL.
CONTENTS.
————————————
| Page | ||
| GEOLOGICAL INTRODUCTION . . . . . . | 3 |
|
| TOXODON PLATENSIS, Description | of Cranium . . . . . | 16 |
| of Lower Jaw and Teeth . . . . . | 29 |
|
| MACRAUCHENIA PATACHONICA, | Cervical Vertebræ . . . . . | 35 |
| Lumbar Vertebræ . . . . . | 40 |
|
| Scapula . . . . . | 43 |
|
| Antibrachium and Fore-foot . . . . . | 44 |
|
| Femur . . . . . | 48 |
|
| Tibia, Astragalus, and Metatarsal Bone . . . . . | 50 |
|
| GLOSSOTHERIUM, Fragment of Cranium . . . . . . | 57 |
|
| MYLODON DARWINII, Lower Jaw and Teeth . . . . . . | 63 |
|
| SCELIDOTHERIUM LEPTOCEPHALUM, | Cranium and Teeth . . . . . | 73 |
| Vertebral Column . . . . . | 84 |
|
| Bones of the Extremities . . . . . | 88 |
|
| MEGALONYX JEFFERSONII, Lower Jaw . . . . . . | 99 |
|
| MEGATHERIUM CUVIERI, Cranium and Teeth . . . . . . | 100 |
|
| TESSELATED ARMOUR AND BONES, large Edentata . . . . . . | 106 |
|
| MASTODON ANGUSTIDENS, Cuv. (?) . . . . . . | 108 |
|
| EQUUS, Molar Teeth . . . . . . | 108 |
|
| CTENOMYS PRISCUS, and a large Rodent . . . . . . | 109 |
|
| Geological contemporaneity of preceding extinct Mammalia . . . . . . | 110 |
|
a
LIST OF PLATES.
———————
| Plate
I. |
Cranium of Toxodon Platensis, nat. size, basal view, with the sixth molar, right side upper jaw, and grinding surface of the seventh molar left side. | |
| II. | Cranium of Toxodon Platensis, side view. | |
| III. | Ditto, upper view. | |
| IV. | Ditto, back view, with two views
of the sixth molar, right side, upper jaw. |
|
| V. | Fragment of lower jaw, and teeth of Toxodon Platensis. | |
| VI. | Cervical vertebræ of the Macrauchenia Patachonica. | |
| VII. | Ditto, ditto, and of Auchenia Llama. | |
| VIII. | Lumbar vertebræ of the Macrauchenia Patachonica. | |
| a. | Medium convex articular surface joining the body of the sacrum. | |
| b. b. | Lateral concavities joining the transverse processes of the sacrum. | |
| c. | Ossified ligament of an anchylosed lumbar vertebra. | |
| IX. | Fig. 1 and 2. Fragments of the Scapula. | |
| Fig. 3. Distal end of the femur of Macrauchenia Patachonica. | ||
| X. | Proximal ends of the anchylosed radius and ulna of Macrauchenia Patachonica. | |
| XI. | Bones of the right fore-foot, Macrauchenia Patachonica. | |
| XII. | Femur, Macrauchenia Patachonica. | |
| XIII. | Anchylosed tibia and fibula, Macrauchenia Patachonica. | |
| XIV. | Right astragalus, Macrauchenia Patachonica. | |
| XV. | Fig. 1. External metatarsal bone of the right hind-foot. | |
| Fig. 2. Proximal end of internal metacarpal bone, right fore-foot. | ||
| XV. | (continued.) | |
| Fig. 3 and 4. Proximal end of middle metacarpal bone. | ||
| Fig. 5 Proximal end of external metacarpal bone, right fore-foot, Macrauchenia Patachonica. | ||
| XVI. | Fragment of cranium of the Glossotherium. | |
| Fig. 1. Side view. | ||
| Fig. 2. Base view. | ||
| Fig. 3. Inside view, all half natural size. | ||
| Fig. 4. Os tympanicum, natural size. | ||
| XVII. |
Fig. 1. Megalonyx Jeffersonii. | |
| Fig. 2. Megalonyx laqueatus. | ||
| Fig. 3 and 4. Mylodon Harlani. | ||
| Fig. 5. Mylodon Darwinii. | ||
| XVIII. |
Lower jaw of the Mylodon Darwinii. | |
| XIX. |
Fig. 1. Side view of the lower jaw. | |
| Fig. 2 and 3. Last molar tooth. | ||
| Fig. 4. Front view of the symphysis of the lower jaw, Mylodon Darwinii. | ||
| XX. |
Portion of the skeleton of the Scelidotherium leptocephalum. | |
| XXI. |
Fig. 1. Side view of the cranium. | |
| Fig. 2. Back view of the cranium. | ||
| Fig. 3, 4, and 5, anterior molar, upper jaw Scelidotherium. | ||
| XXII. | Upper view of the cranium of the Scelidotherium. | |
| XXIII. |
Fig. 1. Inside view of cranium. | |
| Fig. 2. Articular surface for the lower jaw. | ||
| Fig. 3. Section of upper jaw and molar teeth. | ||
| Fig. 4. Section of lower jaw and molar teeth, Scelidotherium. | ||
| XXIV. | Cervical and anterior dorsal vertebræ of, | |
| Fig. 1. Scelidotherium. |
| XXIV. | (continued). | |
| Fig. 2. Orycteropus. | ||
| Fig. 3. Dasypus. | ||
| Fig. 4. Myrmecophaga jubata; all one-third nat. size. | ||
| XXV. | Fig. 1 and 2. Humerus, radius and ulna of Scelidotherium. | |
| Fig. 3. Head of humerus. | ||
| Fig. 4. Head of radius. | ||
| Fig. 5, 6, and 7. Femur of Scelidotherium. | ||
| XXVI. | Fig. 1, 3, and 5. Astragalus of Megatherium. | |
| Fig. 2, 4, and 6, Astragalus of Scelidotherium. | ||
| XXVII. | Fig. 1 and 2. Patella and head of tibia of Scelidotherium. | |
| Fig. 3, 4, and 5. Ungueal phalanx of Scelidotherium. | ||
| XXVIII. | Fig. 1. Astragalus of Megatherium. | |
| Fig. 2. Astragalus of Scelidotherium. | ||
| Fig. 3-6. Astragalus of Mylodon or Megalonyx. | ||
| XXIX. | Lower jaw of Megalonyx Jeffersonii. | |
| XXX. | Fragment of the Cranium of Megatherium Cuvieri. | |
| XXXI. | Section of the upper jaw and teeth of the Megatherium Cuvieri. | |
| XXXII. | Fig. 1. Magnified view of structure of molar of Megatherium. | |
| Fig. 2-5. Ungueal phalanx and portions of tesselated armour of Hoplophorus euphractus. | ||
| Fig. 6-11. Jaws and teeth of Ctenomys priscus. | ||
| Fig. 12. Bones of right hind-foot of a Rodent. | ||
| Fig. 13 and 14. Fossil tooth of a Horse. |
THE NUMBERS ALREADY PUBLISHED OF THIS WORK ARE AS FOLLOWS:
Nos. I. VII. VIII. AND XIII.
FOSSIL MAMMALIA,
BY RICHARD OWEN, Esq. F.R.S.
Professor of Anatomy and Physiology to the Royal
College of Surgeons, London.
This Division of the Work is now complete, Price £1 10s.
Nos. II. IV. V. and X. MAMMALIA,
BY GEORGE R. WATERHOUSE, Esq. Curator of The Zoological Society of London, &c.
This Division of the Work is now complete, Price £1 18s.
NOS. III. VI. IX. AND XI.
BIRDS.
BY JOHN GOULD, Esq. F.L.S.
WITH A NOTICE OF THEIR HABITS AND RANGES,
BY CHARLES DARWIN, Esq. M.A., Sec. G.S.
*** The whole of the Plates are engraved in the highest style of Art, from Drawings taken expressly for this Work, and beautifully coloured after Nature.
NOW PUBLISHING IN PARTS,
UNDER THE AUTHORITY OF THE LORDS COMMISSIONERS OF HER MAJESTY'S TREASURY,
ILLUSTRATIONS OF
THE ZOOLOGY OF SOUTH AFRICA:
Consisting chiefly of Figures and Descriptions of the Objects of Natural History collected during an Expedition into the Interior of South Africa, in the years 1834, 1835, and 1836, fitted out by
"The Cape of Good Hope Association for Exploring Central Africa."
TOGETHER WITH
A SUMMARY OF AFRICAN ZOOLOGY,
AND AN INQUIRY INTO THE GEOGRAPHICAL RANGES OF SPECIES IN THAT QUARTER OF THE GLOBE.
BY ANDREW SMITH, M.D.
SURGEON TO THE FORCES, AND DIRECTOR OF THE EXPEDITION.
The Drawings for this Work were made on the spot, and have since been engraved in the very first style, and beautifully coloured after Nature.
The First Ten Parts are now ready, Nine containing Ten Engravings each with Descriptive Letter-press, Price 10s.; and One of Insects, Price 16s.
*** In order to secure to science the full advantage of Discoveries in Natural History, the Lords Commissioners of Her Majesty's Treasury have been pleased to make a liberal grant of money towards defraying part of the expenses of these two important publications. They have, in consequence, been undertaken on a scale worthy of the high patronage thus received, and are offered to the public at a much lower price than would otherwise have been possible.
PUBLISHED BY SMITH, ELDER AND CO., 65, CORNHILL.