[page] 151
On the Three remarkable Sexual Forms of Catasetum tridentatum, an Orchid in the possession of the Linnean Society. By CHARLES DARWIN, M.A., F.R.S., F.L.S.1
[Read April 3, 1862.]
THE President and Officers of the Linnean Society having kindly permitted me to examine the remarkable specimen, preserved in spirits in their collection, of an Orchid bearing flowers of two supposed genera, and known sometimes to bear the flowers of a third genus, I have thought that the Society might like to hear a short account and explanation of this singular case. The following details will hereafter appear in a small work on the 'Fertilization of Orchids by Insect-agency,' which I am preparing for early publication.
Botanists were astonished when Sir R. Schomburgk*2 stated that he had seen three distinct forms, believed to constitute three distinct genera, namely Catasetum tridentatum, Monachanthus viridis, and Myanthus barbatus, all growing on the same plant. Lindley† remarked that "such cases shake to the foundation all our ideas of the stability of genera and species." Sir R. Schomburgk affirms that he has seen hundreds of plants of Catasetum tridentatum in Essequibo3 without ever finding one specimen with seeds‡, but that he was surprised at the gigantic seed-vessels of the Monachanthus; and he correctly remarks that here we have traces of sexual difference in Orchideous flowers.
The general appearance of the flower of Catasetum tridentatum, in its natural position, is given in the diagram, p. 152 (fig. 1); but the two lower sepals have been cut off. The column is figured separately in an upright position, showing the two curious prolongations of the rostellum, or, as I shall call them, the antennæ.
* 'Transactions of the Linnean Society,' vol. xvii. p. 522. Another account, by Dr. Lindley, has appeared in the 'Botanical Register,' vol. xxiii. fol. 1951, of a distinct species of Myanthus and Monachanthus appearing on the same scape: he alludes also to other cases. Some of the flowers were in an intermediate condition, which is not surprising, seeing that in diœcious plants we sometimes have a partial resumption of the characters of both sexes. Mr. Rogers,4 of River Hill, informs me that he imported from Demerara a Myanthus, but that when it flowered a second time it was metamorphosed into a Catasetum. Dr. Carpenter5 ('Comparative Physiology,' fourth edition, p. 633) alludes to an analogous case which occurred at Bristol.
† 'The Vegetable Kingdom,' 1853, p. 178.
‡ Brongniart6 states (Bull. de la Soc. Bot. de France, 1855, tom. ii. p. 20) that M. Neumann,7 a skilful fertilizer of Orchids, could never succeed in fertilizing Catasetum.
1 This paper was largely an extract of Orchids, pp. 236-8, a book which Darwin had been working on since mid-1861. Darwin explained that the three radically different flowers produced by this orchid, which had led them to be classed in different genera, were three sexual forms male, female, and hermaphrodite. See Correspondence vol. 10.
2 Robert Hermann Schomburgk (1804-1865), German-born explorer and naturalist, see Schomburgk 1837.
3 A territory in British Guiana.
4 John Rogers (1807-1867), barrister and orchid grower who resided at River Hill House, Sevenoaks Weald, Kent.
5 William Benjamin Carpenter (1813-1885) physiologist and naturalist.
6 Adolphe-Théodore Brongniart (1801-1876), French botanist.
7 Louis Neumann (d. 1893), gardener at the Muséum d'Histoire Naturelle c. 1849-1878.
[page] 152
A deep chamber, which from its homological relations must be called the stigmatic chamber, lies between the bases of the an-
Fig. 1.
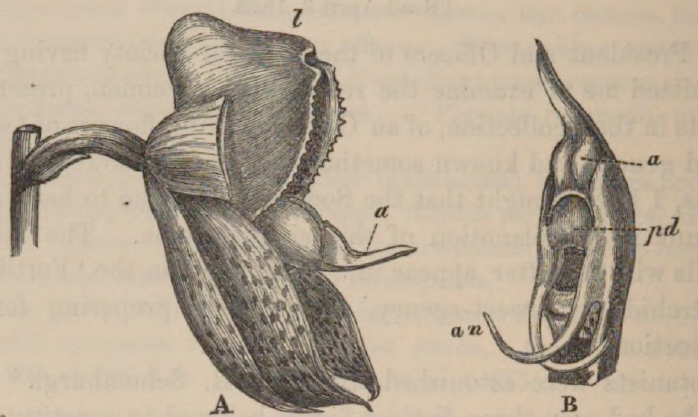
CATASETUM TRIDENTATUM.
| a. | anther. | an. | antennæ. |
| pd. | pedicel of pollinium. | l. | labellum. |
A. Side view of flower in its natural position with the properly lower sepals cut off.
B. Front view of column, placed upright.
tennæ; and the anther, with its concealed pollen-masses, is seated above. My object is not here to describe in detail the structure of the flower and its curious mechanism. But it must be observed that the ovarium is much shorter, thinner, less deeply furrowed, more solid in the centre, and the bract at its base smaller, than in the two succeeding sexual forms presently to be described. The ovarium is bent so that the bucket-like labellum stands uppermost, instead of forming the lower lip as in most Orchids.
From what I had myself observed previously to reading Sir R. Schomburgk's paper, I was led to examine carefully the female organs of this species, and, I may add, of C. callosum and C. saccatum. In no case was the stigmatic surface viscid, as it is in all other Orchids (excepting Cypripedium), and as is indispensable for securing the pollen-masses on the rupture of the caudicles. I carefully looked to this point in both young and old flowers of C. tridentatum. When the surface of the stigmatic chamber and of the stigmatic canal of the above-named three species is scraped off, after having been kept in spirits of wine, it is found to be composed of utriculi (with nuclei of the proper shape), but not nearly so numerous as with ordinary Orchids. The utriculi cohere more together, and are more transparent. I examined for comparison the utriculi of many kinds of Orchids, which had been kept in
[page] 153
spirits, and in all found they were much less transparent. Again, in all three species of Catasetum the ovule-bearing cords are short, and the ovules present a considerably different appearance, in being thinner, more transparent, and less pulpy than in the numerous other Orchids examined for comparison. They were, however, in not so completely an atrophied condition as in the genus Acropera. Although they correspond so closely in general appearance and position with true ovules, perhaps I have no strict right so to designate them, as I was unable in any case to make out the opening of the testa and the included nucleus; nor were the ovules ever inverted. From these several facts—namely, the shortness, thinness, and smoothness of the ovarium, the shortness of the ovule-bearing cords, the state of the ovules themselves, the stigmatic surface not being viscid, the empty condition of the utriculi—and from Sir R. Schomburgk never having seen C. tridentatum producing seed in its native home, we may confidently look at this species of Catasetum, as well as the other two species, as male plants.
Fig. 2.
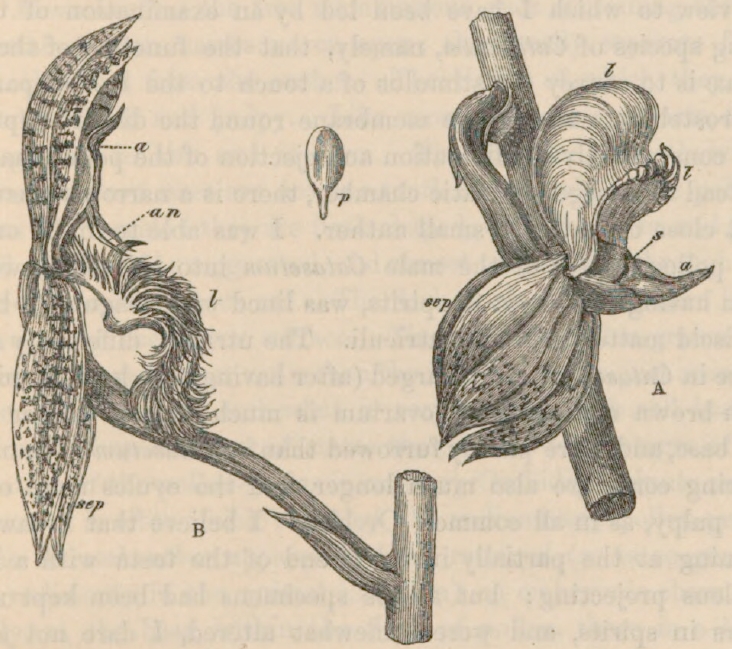
MYANTHUS BARBATUS. MONACHANTHUS VIRIDIS.
| a. | anther. | p. | pollen-mass, rudimentary. |
| an. | antennæ. | s. | stigmatic cleft. |
| l. | labellum. | sep. | two lower sepals. |
A. Side view of Monachanthus viridis in its natural position.
(The shading in both drawings has been added from M. Reiss's drawing in the 'Linnean Transactions.')1
B. Side view of Myanthus barbatus in its natural position.
1 C. F. Reiss of Demerara made a drawing published in Schomburgk 1837.
[page] 154
With respect to Monachanthus viridis and Myanthus barbatus, these two forms are seen, in the specimen sent home by Sir R. Schomburgk, and preserved in spirits in the Society's collection, to be borne on the same spike. They are represented in the diagrams, page 153. The flower of the Monachanthus, like that of the Catasetum, grows lower side uppermost. The labellum is not nearly so deep, especially on the sides, and its edges are crenated. The other petals and sepals are all reflexed, and are not so much spotted as in the Catasetum. The bract at the base of the ovarium is much larger. The whole column, especially the filament at its summit and the spike-like anther, is much shorter; and the front of the rostellum is much less protuberant. The antennæ or horn-like prolongations of the rostellum are entirely absent. The pollen-masses are rudimentary: I could find no trace of a viscid disk or of a pedicel; if they exist, they must be quite rudimentary, for there is hardly any space for the imbedment of the disk. The absence of the antennæ in this Orchid, which has no pollen-masses to eject, is an interesting fact, as it accords with the view to which I have been led by an examination of three living species of Catasetum, namely, that the function of the antennæ is to convey the stimulus of a touch to the medial part of the rostellum, causing the membrane round the disk to rupture, and consequently the liberation and ejection of the pollen-masses. Instead of a large stigmatic chamber, there is a narrow transverse cleft close beneath the small anther. I was able to insert one of the pollen-masses of the male Catasetum into this cleft, which, from having been kept in spirits, was lined with coagulated beads of viscid matter and with utriculi. The utriculi, differently from those in Catasetum, were charged (after having been kept in spirits) with brown matter. The ovarium is much longer, thicker near the base, and more plainly furrowed than in Catasetum; the ovule-bearing cords are also much longer, and the ovules more opake and pulpy, as in all common Orchids. I believe that I saw the opening at the partially inverted end of the testa with a large nucleus projecting; but as the specimens had been kept many years in spirits, and were somewhat altered, I dare not speak positively. From these several facts it is almost certain that Monachanthus is a female plant; and Sir R. Schomburgk saw it seeding abundantly. Altogether this flower differs in a most remarkable manner from that of the male Catasetum tridentatum, and it is no wonder that they were formerly ranked as distinct genera.
[page] 155
The pollen-masses offer so curious and good an illustration of a structure in a rudimentary condition, that they are worth description; but first I must briefly describe the perfect pollen-masses of the male Catasetum. These consist of a large sheet of cemented or waxy pollen-grains, folded over so as to form a sac with an open slit along the lower surface; into this slit cellular tissue enters whilst the pollen is in the course of development in the bud. Within the lower and produced end of each pollen-mass a layer of highly elastic tissue, forming the caudicle, is attached, the other end being attached to the strap-shaped pedicel of the pollinium. The exterior grains of pollen are more angular, have thicker walls, and are yellower than the interior grains. In the early bud the two pollen-masses are enveloped in two conjoined membranous sacs, which are soon penetrated by the two produced ends of the pollen-masses, and by their caudicles; and then the ends of the caudicles adhere to the pedicel. Before the flower expands, the membranous sacs including the pollen-masses open, and leave them resting naked on the back of the rostellum.
In Monachanthus the two membranous sacs containing the rudimentary pollen-masses never open; they easily separate from each other and from the anther. The tissue of which they are formed is thick and pulpy. Like most rudimentary parts, they vary greatly in size and in form. The included, and therefore useless, pollen-masses are not one-tenth of the bulk of the pollen-masses of the male: they are flask-shaped, with the lower and produced end greatly exaggerated, and almost penetrating through the exterior or membranous sac. The flask is closed, and there is no fissure along the lower surface. The exterior pollen-grains are square and have thicker walls than the interior grains, just as in the proper male pollen; and what is very curious, each cell has its nucleus. Now R. Brown* states that, in the early stages of the formation of the pollen-grains in ordinary Orchids, a minute areola or nucleus is often visible; so that the rudimentary pollen-grains of the Monachanthus apparently have retained (as is so general with rudiments in the animal kingdom) an embryonic character. Lastly, at the base, within the flask of pollen, there is a little sheet of brown elastic tissue—that is, a vestige of a caudicle—which runs far up the produced end of the flask, but does not (at least in some of the specimens) come to the surface, and could not have been attached to any part of the rostellum. These rudimentary caudicles are, therefore, utterly useless.
* Transactions of the Linnean Society, vol. xvi. p. 711.
[page] 156
We thus see that every single detail of structure of the male pollen-masses, with some parts exaggerated and some parts slightly modified, is represented by these mere rudiments in the female plant. Such cases are familiar to every observer, but can never be examined without renewed interest.
We now come to the third form, Myanthus barbatus, often borne on the same plant with the two preceding forms. Its flower, in external appearance, but not in essential structure, is the most different of all. It generally stands in a reversed position, compared with Catasetum and Monachanthus—that is, with the labellum downwards. The labellum is fringed, in an extraordinary manner, with long papillæ; it has a quite insignificant medial cavity, at the hinder margin of which a curious curved and flattened horn projects. The other petals and sepals are spotted and elongated, with the two lower sepals alone reflexed. The antennæ are not so long as in the male C. tridentatum, and they project symmetrically on each side of the horn-like projection at the base of the labellum, with their tips (which are not roughened with papillæ as in the male flower) almost entering the medial cavity. The stigmatic chamber is of nearly intermediate size between that of the male and female forms; it is lined with utriculi, charged with brown matter. The straight and well-furrowed ovarium is nearly twice as long as in Monachanthus, but is not so thick where it joins the flower; the ovules are not so numerous as in the female form, but are opake and pulpy after having been kept in spirits, and resemble them in all respects. I believe, but dare not speak positively as in the case of the Monachanthus, that I saw the nucleus projecting from the testa. The pollinia are about a quarter of the size of those of the male Catasetum, but have a perfectly well developed disk and pedicel. The pollen-masses were lost in the specimens examined by me; but fortunately M. Reiss has given, in the 'Linnean Transactions,' a drawing of them, showing that they are of due proportional size, and have the proper folded or cleft structure; so that there can hardly be a doubt that they are functionally perfect. As we thus see that both the male and female organs are apparently perfect, Myanthus barbatus may be considered as the hermaphrodite form of the same species, of which the Catasetum is the male, and the Monachanthus the female.
It is not a little singular that the hermaphrodite Myanthus should resemble in its whole structure much more closely the male forms of two distinct species (namely C. saccatum and, more especially, C. callosum) than either its own male or female forms.
[page] 157
Finally, the genus Catasetum is interesting in an unusual degree in several respects. The separation of the sexes is unknown in other Orchids, excepting probably in the allied genus Cycnoches and in one other member of the Vandeæ, namely, Acropera. In Catasetum we have three sexual forms, generally borne on separate plants, but sometimes mingled together; and these three forms are wonderfully different from each other—much more different than, for instance, a peacock is from a peahen. But the appearance of these three forms on the same plant now ceases to be an anomaly, and can no longer be viewed as an unparalleled instance of variability.
Still more interesting is this genus in its mechanism for fertilization. We see a flower patiently waiting, with its antennæ stretched forth in a well-adapted position, ready to give notice whenever an insect puts its head into the cavity of the labellum. The female Monachanthus, not having pollinia to eject, is destitute of antennæ. In the male and hermaphrodite forms, namely Catasetum and Myanthus, the pollinia lie doubled up like a spring, ready to be instantaneously shot forth when the antennæ are touched. The disk end is always projected foremost, and is coated with viscid matter, which quickly sets hard and firmly affixes the hinged pedicel to the insect's body. The insect flies from flower to flower, till at last it visits a female or hermaphrodite plant; it then inserts one of the balls of pollen into the stigmatic cavity. When the insect flies away, the elastic caudicle, made weak enough to yield to the viscidity of the stigmatic surface, breaks, and leaves behind the pollen-mass; then the pollen-tubes slowly protrude, penetrate the stigmatic canal, and the act of fertilization is completed. Who would have been bold enough to surmise that the propagation of a species should have depended on so complex, so apparently artificial, and yet so admirable an arrangement?