[page] 169
On the Sexual Relations of the Three Forms of Lythrum
salicaria.1
By CHARLES DARWIN, F.R.S., F.L.S., &c.
[Read 16 June 1864.]
SOME of the species of Lythrum offer in their manner of fertilization a more remarkable case than can, perhaps, be found in any other plant or animal. In Lythrum salicaria three plainly different forms occur: each of these is an hermaphrodite, each is distinct in its female organs from the other two forms, and each is furnished with two sets of stamens or males differing from each other in appearance and function. Altogether there are three females and three sets of males, all as distinct from each other as if they belonged to different species; and if smaller functional differences are considered, there are five distinct sets of males. Two of the three hermaphrodites must coexist, and the pollen be carried by insects reciprocally from one to the other, in order that either of the two should be fully fertile; but unless all three forms coexist, there will be waste of two sets of stamens, and the organization of the species, as a whole, will be imperfect. On the other hand, when all three hermaphrodites coexist, and the pollen is carried from one to the other, the scheme is perfect; there is no waste of pollen and no false co-adaptation. In short, nature has ordained a most complex marriage-arrangement, namely a triple union between three hermaphrodites,—each her-
1 Purple loosestrife. Edited portions of this article appeared in Forms of flowers, pp. 137-67. Darwin also discussed Lythrum salicaria in Origin 4th ed., p. 323 and Variation 2: 166, 183. See Correspondence vol. 12.
[page] 170
maphrodite being in its female organ quite distinct from the other two hermaphrodites and partially distinct in its male organs, and each furnished with two sets of males.
The three forms may be conveniently called, from the unequal lengths of their pistils, the long-styled, mid-styled, and short-styled. Their existence and differences were first observed by Vaucher*, and subsequently more carefully by Wirtgen;1 but, not being guided by any theory, neither author perceived some of the most curious points of difference. I will first briefly describe the three forms by the aid of the accompanying accurate diagram, which shows the flowers, six times magnified, in their natural position, with their petals and the near side of the calyx removed.
Long-styled form.—This can at once be recognized by the length of the pistil, which is (including the ovarium) fully one-third longer than that of the mid-styled, and more than thrice as long as that of the short-styled form. It is so disproportionately long, compared with the flower, that it projects in the bud through the unfolded petals. It stands out considerably beyond the longer stamens; its terminal portion depends a little, but the stigma itself is slightly upturned: the globular stigma is considerably larger than that of the other two forms. The six longer stamens project about two-thirds of the length of the pistil, and correspond in length with the pistil of the mid-styled form. The correspondence with the pistil in length in this and the two following cases is generally very close; the difference, where there is any, being usually in a slight excess of length in the stamens. The six shorter stamens (each of which alternates with a longer one) lie concealed within the calyx; their ends are upturned, and they are graduated in length, so as to form a triple row—both which characters are here much more marked than with the longer stamens, which vary in these respects. The anthers of the shorter stamens are smaller than those of the longer stamens. Knowing that the pollen differs greatly in the longer and shorter stamens of the two other forms, I carefully compared that of the two sets of stamens in this form: in both the pollen-grains are yellow, but they are a little larger in the longer than in the shorter stamens. The difference is slight, so that I convinced myself of its reality only by putting two small heaps close together under the compound mi-
* Hist. Phys. des Plantes d'Europe, tom. ii., 1841, p. 371.2 Wirtgen, "Ueber Lythrum salicaria und dessen Formen," Verhand. des naturhist. Vereins der preuss. Rheinl., 5. Jahrgang, 1848, S. 7.3
1 Philipp Wilhelm Wirtgen (1806-1870), German teacher and botanist who lived in Koblenz.
2 Vaucher 1841.
3 Wirtgen 1848.
[page] 171
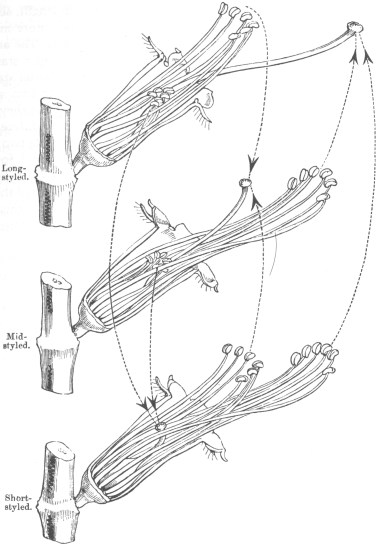
Diagrams of the flowers of the three forms of Lythrum salicaria, in their natural position, with the petals removed and with the near side of the calyx cut away: enlarged six times.
The dotted lines with the arrows show which pollen must be applied to each stigma to cause full fertility.
[page] 172
croscope, and I found I could always (with one exception) distinguish them: I then showed the specimens to two other persons, and they likewise distinguished the two kinds and pointed out which was the largest. The capsules of this form contain, on an average, 93 seeds: how this average was obtained will presently be explained. I repeatedly observed that the seed, when cleaned, seemed larger than that from the mid-styled or short-styled forms; consequently I placed 100 long-styled seeds in a good balance, and by the double method of weighing found that they equalled 121 seeds of the mid-styled and 142 of the short-styled; or, in short, that five long-styled seeds equalled six mid-styled and seven short-styled seeds. These slight differences in the weight of the seed, and, as we shall soon see, in the average number produced, are worth recording, as they characterize not mere varieties but coexisting forms of the same species.
Mid-styled form.—The pistil occupies the position represented in the diagram, with its extremity considerably, but in a variable degree, upturned; the stigma is seated between the anthers of the long and the short stamens. The six longer stamens correspond in length with the pistil of the long-styled form; their filaments are coloured bright pink; the anthers are dark-coloured, but from containing bright green pollen and from their early dehiscence they appear emerald-green. Hence the general appearance of these stamens is remarkably dissimilar from that of the longer stamens of the long-styled form. The six shorter stamens, enclosed within the calyx, resemble in all respects the shorter stamens of the long-styled form, and both correspond in length with the short pistil of the short-styled form. The green pollen-grains of the longer stamens are plainly larger than the yellow pollen-grains of the shorter anthers: this fact was conspicuous in several camera-lucida drawings made for me by my son, Mr. W. E. Darwin.1 There is some variability in size, but 12/7000 of an inch may be taken as about the average diameter of the green pollen-grains when distended with water, and 9/7000 as the diameter of the yellow grains of the shorter stamens; so that the difference in diameter is in about the proportion of four to three. The capsules contain, on an average, 132 seeds; but, perhaps, as we shall see, this is rather too high an average. The seeds themselves are smaller than those of the long-styled form.
Short-styled form.—The pistil is here very short, not one-third of the length of that of the long-styled form. It is enclosed within the calyx, which, differently from in the other two forms,
1 William Erasmus Darwin (1839-1914), Darwin's eldest child. The drawings survive in DAR27.2.
[page] 173
does not enclose any anthers. The end of the pistil is generally bent upwards at right angles. The six longer stamens, with their pink filaments and green pollen, resemble in size of the grains and in all respects the longer stamens of the mid-styled form, and both correspond in length with the long-styled pistil. The six shorter stamens, with their uncoloured filaments and yellow pollen, resemble in size of the grains and in all respects the longer stamens of the long-styled form, and both correspond in length with the mid-styled pistil. The capsules contain fewer seeds on an average than in either of the preceding forms, namely 83.5, and they are considerably smaller in size. In this latter respect, but not in number, there is a gradation parallel to that of the length of the pistil, the long-styled having the largest, the mid-styled the next in size, and the short-styled the smallest seed.
From this description we see that there are three distinct female organs, or rather females as they are borne on distinct individuals, differing in the length and curvature of the style, in the size of the stigma, and in the number and size of the seed. In the three forms, taken together, there are thirty-six stamens or males, and these can be divided into three sets of a dozen each, differing from each other in length, curvature, and colour of the filaments, in the size of the anthers, and especially in the colour and diameter of the pollen-grains. Each of the three forms bears half-a-dozen of one kind of stamens and half-a-dozen of another kind, but not all three kinds. The three kinds correspond in length with the three pistils: the correspondence is always between half the stamens borne by two forms with the pistil of a third form. These remarks apply to the structure, and not, as yet, to the functions, of the reproductive organs.
I ascertained the average number of seed by counting them in eight fine selected capsules taken from plants of the three forms growing wild, and the result was, as we have seen, for the long-styled (neglecting decimals) 93, mid-styled 132, and short-styled 83. I should not have trusted this result, but I had a number of plants in my garden which, from their youth, did not yield the full complement of seed, but they were of exactly the same age and grew under exactly the same conditions, and were freely visited by bees. I took six fine capsules from each, and found the average to be for the long-styled 80, for the mid-styled 97, and for the short-styled 61. Lastly, I made numerous artificial unions, and, as may be seen in the following Tables, these gave in the long-styled an average of 90 seeds, in the mid-styled 117,
[page] 174
and in the short-styled 71. So that we have good concurrent evidence of the different average production of seed by the three forms. To show that the artificial fertilizations, presently to be described, produced their full effect and may be trusted, I may state that one mid-styled capsule yielded 151 good seeds, which is the exact number of the finest wild capsule examined by me. Artificially fertilized short- and long-styled capsules actually produced a greater number of seeds than I have found in wild plants, but then I did not examine many of the latter. This Lythrum, I may add, offers a remarkable instance, how profoundly ignorant we are of the life-conditions of each species: naturally it grows "in wet ditches, watery places, and especially on the banks of streams," and though it produces so many minute seeds, it never spreads on the adjoining land; yet, planted in my garden, on clayey soil lying over the chalk, and which is so dry that a rush cannot be found, it thrives luxuriantly, grows to above six feet in height, produces self-sown seedlings, and (which is a severer test) is fully as fertile as in a state of nature. Nevertheless it would be almost a miracle to find this plant spontaneously growing on such land as my garden, though under its native climate.
According to Vaucher and Wirtgen, the three forms coexist in all parts of Europe. Some friends gathered for me in North Wales a number of twigs from separate plants growing near each other, and then classified them. My son did the same in Hampshire, and here is the result:—
| Long-styled | Mid-styled. | Short-styled. | Total. | |
| North Wales | 95 | 97 | 72 | 264 |
| Hampshire . | 53 | 38 | 38 | 129 |
| Total . . . . | 148 | 135 | 110 | 393 |
If twice or thrice the number had been collected, probably the three forms would have been found nearly equal; I infer this from considering the above figures, and from my son telling me that if he had collected in another spot, he felt sure that the mid-styled plants would have been in excess. I several times sowed small parcels of seed, and raised all three forms; but I neglected to record the parent form, except in one instance, in which I raised from short-styled seed twelve plants, of which only one turned out long-styled, four mid-styled, and seven short-styled.
[page] 175
Insects are necessary for the fertilization of this Lythrum. During two years I kept two plants of each form protected, and in the autumn they presented a remarkable contrast in appearance with the adjoining uncovered plants, which were densely covered with capsules. In 1863 a protected long-styled plant produced only five poor capsules; two mid-styled plants produced the same number; and two short-styled plants between them produced only one: these capsules contained very few seed; yet the plants were fully productive when artificially fertilized under the net. In a state of nature the flowers are incessantly visited for their nectar by hive- and humble-bees and various Diptera. The nectar is secreted all round the base of the ovarium; but a passage is formed along the upper and inner side of the calyx by the lateral deflection (not represented in the diagram) of the basal portions of the filaments; so that insects invariably alight on the upper side of the flowers, on the projecting stamens and pistil, and insert their probosces along the upper inner margin of the calyx. We can now see why the ends of the stamens with their anthers, and the ends of the pistils with their stigma, are a little upturned, in order that they may brush against the lower hairy surfaces of the insects' bodies. The short stamens which lie enclosed within the calyx of the long- and mid-styled forms can be touched only by the proboscis and the narrow chin of the sucking bee; hence they have their ends more upturned, and they are graduated in length, so as to fall into a narrow file, three deep, sure to be raked by the thin intruding proboscis. The anthers of the longer stamens stand laterally further apart and are more nearly of the same length, for they have to brush against the whole breadth of the insect's body. I may here incidentally remark, that in very many flowers the pistil, or the stamens, or both, are rectangularly bent to one side of the flower: this bending may be permanent, as with Lythrum and many others, or may be effected (as in Dictamnus fraxinella and many others) by a temporary movement which occurs in the stamens when the anthers dehisce, and in the pistil when the stigma is mature; but these two movements are by no means always contemporaneous in the same flower. Now I have found no exception to the rule, that when the stamens and pistil are bent, the bending is exactly to that side of the flower which secretes nectar (even though there be a rudimentary nectary of large size on the opposite side, as in some species of Corydalis); or, when nectar is secreted on all
[page] 176
sides, to that side where the structure of the flowers allows the easiest access to it, as in Lythrum, Papilionaceous flowers, and many others. The rule consequently is that when the pistil and stamens are bent, the stigma and anthers are brought into the pathway towards the nectary. There are a few cases which seem to be exceptions, but they are not so in truth: for instance, in the Gloriosa lily, the stigma of the grotesque and rectangularly bent pistil is brought, not into the pathway from the open air towards the nectar-secreting recesses of the flower, but into the circular route from one nectary to the other; in Scrophularia aquatica the pistil is bent downwards from the mouth of the flower, but it thus strikes the pollen-dusted breasts of the wasps which habitually visit these ill-scented blooms. In the above rule we see one more instance of the supreme dominating power of insects over all the minor structural details of flowers, especially of those which have irregular corollas. Flowers which are fertilized by the wind must of course be excepted, but I do not know of a single instance of an irregular flower which is fertilized or crossed by this means.
I have delayed too long on these points, but I must allude to one other. We have seen that the three pistils of different lengths have each two half-dozen sets of stamens of corresponding length. When bees suck the flowers, the longest stamens, bearing the green pollen, rub against the abdomen and the interior sides of the posterior legs, as does likewise the stigma of the long-styled form. The stamens of middle length and the stigma of the mid-styled form rub against the under side of the thorax and between the front pair of legs. The shortest stamens and the stigma of the short-styled form must rub against the proboscis and chin; for the bees in sucking insert only the front of their heads into the calyx. On catching bees, I observed much green pollen on the inner sides of the hind legs and on the abdomen, and much yellow pollen on the under side of the thorax. There was also pollen on the chin, and, it may be presumed, on the proboscis, but this was difficult to observe. I had, however, independent proof that pollen is carried on the proboscis; for in a protected short-styled plant (which produced only two capsules) one small branch was accidentally left during many days pressing against the fine net, and bees were seen inserting their probosces through the meshes, and in consequence numerous capsules were formed on this one small branch. From these several facts it follows that insects would chiefly carry to the
[page] 177
stigma of each form pollen from the stamens of corresponding length; and we shall presently see the importance of this adaptation. It must not, however, be supposed that the bees do not get more or less dusted all over with the several kinds of pollen; they certainly do, as could be seen with the green pollen from the longest stamens. Moreover, a case will presently be given of a long-styled plant which grew absolutely by itself, and produced an abundance of capsules, which must have been fertilized by its own two kinds of pollen; but these capsules contained a very poor average of seed. Hence insects, and chiefly bees, act both as general carriers of pollen, and as special carriers of the right kind*.
Variability.—Before passing on to more important topics, I must say a few words on this head. Wirtgen remarks† on the variability in the branching of the stem, in the length of the bracteæ, size of the petals, and in several other respects. The plants now growing in my garden have their leaves arranged oppositely, alternately, and in whorls of three, and differ greatly in shape. The stems of the plants bearing leaves in whorls are hexagonal; those of the other plants are quadrangular. But we are concerned only with the reproductive organs: the upward bending of the pistil is variable, and in a remarkable degree in the short-styled form, in which it is sometimes straight, sometimes slightly curved, but generally upturned at right angles. The stigma of the long-styled pistil frequently has longer papillæ or is rougher than that of the mid-styled, and this than that of the short-styled form; but this character, though fixed and uniform with the two forms of Primula, is here variable, and I have
* In my paper on the two forms of Primula (Journal Proc. Linn. Soc. 1862, p. 85) [Darwin 1862] I stated that I had only occasionally seen humble-bes sucking the flowers of the Cowslip (P. veris). Since then I have had some beds in my garden containing nearly 700 plants, and these were incessantly visited by Bombus hortorum and B. muscorum. I caught some of these bees, and I found (as I had anticipated in my paper, p. 86) that a vast majority of the pollen-grains which adhered to the base of the proboscis were large-sized and had come from the long stamens of the short-styled form, and were thus placed ready to fertilize the stigma of the long-styled form. On the other hand, on the middle, and near the tip of the proboscis, a very large proportion of the pollen-grains were of the small size, and had come from the short stamens of the long-styled form. My son caught, also, a moth (Cucullia verbasci) hovering over the bed, and I found on its proboscis a similar distribution of the two kinds of pollen-grains. I give these facts as a further illustration of the importance of the relative lengths of the stamens and pistil.
† Verhand. des naturhist. Vereins, 5. Jahrgang, 1848, S. 11, 13.
[page] 178
seen mid-styled stigmas rougher than those of the long-styled. The degree to which the longer and middle stamens are graduated in length and are upturned at their ends is variable; sometimes all are equal. The colour of the green pollen in the long stamens is variable*, and is sometimes pale greenish yellow; in one short-styled plant it was almost white. The grains vary a little in size: I examined one short-styled plant with the grains above the average size; and I have seen a long-styled plant with undistinguishable grains from the longer and shorter anthers. We have here considerable fluctuations of character; and if any of these slight structural differences were of direct service to the plant, or were correlated with useful functional differences, we can perceive that the species is just in that state in which natural selection might readily do much for its modification.
To return to our proper subject—we see that there are three kinds of females and three kinds of males, each kind of the latter being borne by half-dozens on two of the three forms. It remains to discover whether these several sexes or sexual organs differ from each other in function. Nothing brings more prominently forward the complexity of the reproductive system of this extraordinary plant, than the necessity, in order to ascertain the above fact, of artificially making eighteen distinct unions. Thus the long-styled form had to be fertilized with pollen from its own two distinct kinds of anthers, from the two in the mid-styled, and from the two in the short-styled form. The same process had to be repeated with both the mid- and short-styled forms. It might have been thought sufficient to have tried on each stigma the green pollen, for instance, from either the mid- or short-styled longer stamens, and not from both; but the result proves that this would have been insufficient, and that it was necessary to try all six kinds of pollen on each stigma. As in artificial fertilizations there will always be some failures, it would have been advisable to have
* Lagerstrœmia Indica, one of the Lythraceæ, is strangely variable in its stamens—I presume in part due to its growth in a hothouse. The most perfect flowers produced with me five very long stamens with thick flesh-coloured filaments and green pollen, and from nineteen to twenty-nine short stamens with yellow pollen; but many flowers produced only one, two, three, or four long stamens with green pollen, which in some of the anthers was wholly replaced by yellow pollen; one anther offered the singular case of half, or one cell being filled with bright green, and the other cell with bright yellow pollen. One petal had a furrow near its base, which contained pollen. According to analogy with Lythrum, this species would produce three forms; if so, the above plant was a mid-styled form: it was quite sterile with its own two kinds of pollen.
[page] 179
repeated each of the eighteen unions a score of times; but the labour would have been too great; as it was, I made 223 artificial unions; i. e., I fertilized, on an average, above a dozen flowers in the eighteen different methods. Each flower was castrated; the adjoining buds had to be removed, that the marking-thread, wool, &c. might be safely secured; and after each fertilization the stigma had to be examined with a lens to see that there was sufficient pollen. Plants of all three forms were protected during two years by large nets on a framework; two plants were used during one or both years, in order to avoid any individual peculiarity in any one plant. As soon as the flowers withered, the nets were removed; and in the autumn the capsules were daily inspected; when the seeds were ripe they were counted under the microscope. I have given these details that confidence may be placed in the following Tables, and as some excuse for two blunders which, I believe, I made. These blunders are referred to, with their probable causes, in two notes to the Tables; the erroneous numbers, however, are entered in the Tables, that it may not be supposed that I have in any one instance tampered with the results.
A few words explanatory of the three Tables must be given. Each is devoted to one form, and is divided into six compartments. The two upper ones in each table give the product of good seed from the application of pollen from the two sets of stamens which correspond in length with the pistil of that form. The two next lower compartments show the result of pollen from the other two sets of stamens, which do not correspond in length with the pistil, and which are borne by the same two forms. The two lowest compartments show the result of the application of each form's own two kinds of pollen. The term "own pollen," used here and in the Tables, does not mean pollen from the flower to be fertilized—for this was never used—but from another flower on the same plant, or more commonly from a distinct plant of the same form. In the result given, "0" generally means that no capsule was produced, or that the capsule contained no good seed. In some part of each row of figures in each compartment, a short horizontal line may be seen; the unions above this line were made in 1862, and below it in 1863. It is of importance to observe this, as it shows that the same general result ensued in two successive years; but more especially because 1863 was a very hot and dry season, and the plants had occasionally to be watered. This did not prevent the full complement of seed being produced from
[page] 180
the more fertile unions; but it rendered the less fertile unions even more sterile than they otherwise would have been. I have seen striking instances of this same fact in making homomorphic and heteromorphic unions in Primula*; and it is well known that the conditions of life must be highly favourable to give any chance of producing hybrids from species which cross with difficulty.
Table I.—LONG-STYLED FORM.
| I. | II. | ||
| 13 flowers fertilized by the longer stamens of the mid-styled. These stamens equal in length the pistil of the long-styled. | 13 flowers fertilized by the longer stamens of the short-styled. These stamens equal in length the pistil of the long-styled. | ||
| Product of good seed in each capsule. | Product of good seed in each capsule. | ||
| 36 | 53 | 159 | 104 |
| 81 | 0 | 43 | 119 |
| 0 | 0 | 96 poor seed. | 96 |
| 0 | 0 | 103 | 99 |
| 0 | 0 | 0 | 131 |
| — | 0 | 0 | 116 |
| 45 | — | ||
| 41 | 114 | ||
| 38 per cent. of these flowers yielded capsules. Each capsule contained, on an average, 51.2 seed. | 84 per cent. of these flowers yielded capsules. Each capsule contained, on an average, 107.3 seed. | ||
* In the spring of 1862 I crossed forty Cowslip flowers (P. veris) heteromorphically and homomorphically. The plants were accidentally exposed in the greenhouse to too hot a sun, and a number of umbels perished. Some, however, remained in moderately good health, and on these there were twelve flowers which had been fertilized heteromorphically and eleven which had been fertilized homomorphically. The twelve heteromorphic unions yielded seven fine capsules, containing on an average 57.3 good seed. Now mark the difference: the eleven homomorphic unions yielded only two capsules, of which one contained 39 seeds, but so poor, that I do not suppose one would have germinated, and the other only 17 fairly good seed. It would be superfluous to give any more details on this experiment, or on some which I made at the same time on P. Sinensis, after the appearance of Mr. John Scott's admirable paper1 on the various dimorphic species of Primula, in which he confirms my former results, and adds many original and valuable observations. Dr. Hildebrand2 has also (Botanische Zeitung, 1864, Jan. 1, S. 3) confirmed my general results with respect to P. Sinensis, and has corrected an error into which in some unaccountable manner I fell, namely, that the pollen-grains from the long- and short-styled forms were of the same size. Dr. Hildebrand has added a series of new and important experiments, for he fertilized homomorphically a number of flowers with pollen from the same form, and likewise from the same individual flower. These latter he found were thus rendered rather more sterile. This experiment, I believe, has never been systematically tried before.
1 Scott 1864.
2 Friedrich Hermann Gustav Hildebrand (1835-1915), German professor of botany, University of Freiburg, 1868-1907. Hildebrand 1864.
[page] 181
Table I.—LONG-STYLED FORM (continued).
| III. | IV. | ||
| 14 flowers fertilized by the short stamens of the mid-styled. | 12 flowers fertilized by the shorter stamens of the short-styled. | ||
| 3 | 0 | 20 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | — | 0 |
| — | 0 | 0 | 0 |
| 0 | 0 | 0 | |
| 0 | Too sterile for any
average. |
||
Too
sterile for any average. |
|||
| V. | VI. | ||
| 15 flowers fertilized by own longer stamens. | 15 flowers fertilized by own shorter stamens. | ||
| 2 | — | 4 | — |
| 10 | 0 | 8 | 0 |
| 23 | 0 | 4 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
Too
sterile for any average. |
Too sterile for any
average. |
||
I fertilized a considerable number of flowers with pollen, taken by a camel's-hair brush, from both the long and short stamens of their own (long-styled) form; but I did not examine with a lens (as I did in the cases in the Tables) whether sufficient pollen had been placed on the stigma: only 5 capsules were produced, and these yielded on an average 14.5 seed. In 1863 I tried a much better experiment: a long-styled plant was grown by itself, miles away from any other plant, so that its stigmas could have received only the two kinds of pollen proper to this form. The flowers were incessantly visited by bees, so that the stigmas must have received on the most favourable days, and at the most favourable hours, successive applications of pollen: all who have crossed plants know that this highly favours fertilization. This plant produced an abundant crop of capsules; I took by chance 20, and these (excluding one poor one) contained seed as below:—
| 20 | 20 | 35 | 21 | 19 |
| 26 | 24 | 12 | 23 | 10 |
| 7 | 30 | 27 | 29 | 13 |
| 20 | 12 | 29 | 19 | 35 |
This gives an average of 21.5 seed per capsule; and as we know that this form, when standing near plants of the other two forms and fertilized by insects, produces an average of 93.1 seed per capsule, we see that the long-styled form fertilized by its own two pollens yields only between one-fourth and one-fifth of the full number of seed. I have spoken as if this plant had received both its own kinds of pollen, and this is, of course, possible; but, from the enclosed position of the shorter stamens, it is much more probable that the stigma received almost exclusively the pollen from its own longer stamens.
[page] 182
Table II.—MID-STYLED FORM.
| I. | II. | ||
| 12 flowers fertilized by the longer stamens of the long-styled. These stamens equal in length the pistil of the mid-styled. | 12 flowers fertilized by the shorter stamens of the short-styled. These stamens equal in length the pistil of the mid-styled. | ||
| Product of good seed in each capsule. | Product of good seed in each capsule. | ||
| 138 | 122 | 112 | 109 |
| 149 | 50 | 130 | 143 |
| 147 | 151 | 143 | 124 |
| 109 | 119 | 100 | 145 |
| 133 | 138 | 33 | 12 |
| 144 | 0 | — | 141 |
| — | 104 | ||
| 92 per cent. of the flowers (probably 100 per cent.) yielded capsules. Each capsule contained, on an average, 127.3 seed. | 100 per cent. of the flowers yielded capsules. Each capsule contained, on an average, 108.0 seed; or, excluding capsules with less than 20 seed, the average is 116.7 seed. | ||
| III. | IV. | ||
| 13 flowers fertilized by the short stamens of the long-styled. | 15 flowers fertilized by the longer stamens of the short-styled. | ||
| 83 | 12 | 130 | 86 |
| 0 | 19 | 115 |
113 |
| 0 | 85 {seed small and poor. | 14 | 29 |
| — | 0 | 6 |
17 |
| 44 | 0 | 2 | 113 |
| 44 | 0 | 9 | 79 |
| 45 | 0 | — | 128 |
| |
132 | 0 | |
| 54 per cent. of the flowers yielded capsules. Each capsule contained, on an average, 47.4 seed; or, excluding capsules with less than 20 seed, the average is 60.2 seed. | 93 per cent. of the flowers yielded capsules. Each capsule contained, on an average, 69.5 seed; or, excluding capsules with less than 20 seed, the average is 102.8. | ||
| V. | VI. | ||
| 12 flowers fertilized by own longer stamens. | 12 flowers fertilized by own shorter stamens. | ||
| 92 | 0 | 0 | 0 |
| 9 | 0 | 0 | 0 |
| 63 | 0 | 0 | 0 |
| — | 0 | — | 0 |
|
136?* |
0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | ||
| Excluding the capsule with 136 seed, 25 per cent. of the flowers yielded capsules, and each capsule contained, on an average, 54.6 seed; or, excluding capsules with less than 20 seed, the average is 77.5. | 0 per cent. of the flowers yielded capsules. | ||
* I have hardly any doubt that this result of 136 seed is due to a gross error. The flowers to be fertilized by their own longer stamens were first marked by
[page] 183
I fertilized a considerable number of flowers with pollen, taken by a camel's-hair brush, from both the long and short stamens of their own (short-styled) form; but I did not examine with a lens (as I did in the cases in the Tables) whether sufficient pollen had been placed on the stigma: only 5 capsules were produced, and these yielded, on an average, 11.0 seed.
Table III.—SHORT-STYLED FORM.
| I. | II. | ||
| 12 flowers fertilized by the shorter stamens of the long-styled. These stamens equal in length the pistil of the short-styled. | 13 flowers fertilized by the shorter stamens of the mid-styled. These stamens equal in length the pistil of the short-styled. | ||
| 69 | 56 | 93 | 69 |
| 61 | 88 | 77 | 69 |
| 88 | 112 | 48 | 53 |
| 66 | 111 | 43 | 9 |
| 0 | 62 | 0 | 0 |
| 0 | 100 | 0 | 0 |
| — | — | 0 | |
| 83 per cent. of the flowers yielded capsules. Each capsule contained, on an average, 81.3 seed. | 61 per cent. of the flowers yielded capsules. Each capsule contained, on an average, 64.6 seed. | ||
| III. | IV. | ||
| 10 flowers fertilized by the longer stamens of the long-styled. | 10 flowers fertilized by the longer stamens of the mid-styled. | ||
| 0 | 14 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| — | 0 | — | 0 |
| 23 |
|
0 | |
| Too sterile for any average. | Too sterile for any average. | ||
"white thread," and those by the longer stamens of the long-styled form by "white silk;" a flower fertilized in the latter manner would have yielded about 136 seed, and it may be observed that one such pod is missing, viz., at the bottom of compartment 1. Therefore I have hardly any doubt that I fertilized a flower marked with "white thread," as if it had been marked with "white silk." With respect to the capsule which yielded 92 seed, in the same column with that which yielded 136, I do not know what to think. I endeavoured to prevent pollen dropping from an upper to any lower flower, and I tried to remember to wipe the pincers carefully after each fertilization; but in making eighteen different crosses, sometimes on windy days, and pestered by bees and flies buzzing about, some few errors could hardly be avoided. One day I had to keep a third man by me all the time to prevent the bees visiting the uncovered plants, for in a few seconds' time they might have done irreparable mischief. It was also extremely difficult to exclude minute Diptera from the net. In 1862 I made the great mistake of placing a mid-styled and long-styled under the same huge net: in 1863 I avoided this error.
[page] 184
Table III.—SHORT-STYLED FORM (continued).
| V. | VI. | ||
| 10 flowers fertilized by own longer stamens. | 10 flowers fertilized by own shorter stamens. | ||
| 0 | 0 | 64?* | 0 |
| 0 | 0 | 0 | 0 |
| 0 | 0 | 0 | 0 |
| — | 0 | — | 0 |
| 0 | 0 | 21 | 0 |
| 0 | |
9 | |
| Too sterile for any average. | Too sterile for any average. | ||
I fertilized a number of flowers without particular care with their own two pollens, but they did not produce a single capsule; the position of the stigma within the calyx renders the fertilization without some care difficult.
Summary of the three preceding Tables.
Long-styled form.—Twenty-six flowers fertilized by the stamens of corresponding length, borne by the mid- and short-styled forms, yielded 61.5 per cent. of capsules, which contained, on an average, 89.7 seed.
Twenty-six flowers fertilized by the other and shorter stamens of the mid- and short-styled forms yielded only two very poor capsules.
Thirty flowers fertilized by this form's own two sets of stamens yielded only eight very poor capsules; but flowers well fertilized by bees by one or both of their own kinds of pollen produced numerous capsules containing, on an average, 21.5 seed.
Short-styled form.—Twenty-five flowers fertilized by the stamens of corresponding length, borne by the long- and mid-styled forms, yielded 72 per cent. of capsules, which (excluding one capsule with only nine seeds) contained, on an average, 70.8 seed.
Twenty flowers fertilized by the longer stamens of the long- and mid-styled forms yielded only two very poor capsules.
Twenty flowers fertilized by both their own two sets of stamens yielded only two poor (or perhaps three) capsules.
Mid-styled form.—Twenty-four flowers fertilized by the stamens of corresponding length, borne by the long- and short-styled forms, yielded 96 (probably 100) per cent. of capsules, which con-
* I suspect that, by mistake, I fertilized this flower with the pollen of the shorter stamens of the long-styled form, and it would then have yielded about 64 seed. Flowers to be thus fertilized were marked with black silk; those with the pollen of the shorter stamens of the short-styled with black thread; and thus, I suspect, the mistake arose.
[page] 185
tained (excluding one capsule with 12 seed), on an average, 117.2 seed.
Fifteen flowers fertilized by the longer stamens of the short-styled form yielded 93 per cent. of capsules, which (excluding four capsules with less than 20 seed) contained, on an average, 102.8 seed.
Thirteen flowers fertilized by the shorter stamens of the long-styled form yielded 54 per cent. of capsules, which capsules (excluding one with 19 seed) contained, on an average, 60.2 seed.
Twelve flowers fertilized by own longer stamens yielded 25 per cent. of capsules, which (excluding one with 9 seed) contained, on an average, 77.5 seed.
Twelve flowers fertilized by own shorter stamens yielded not a single capsule.
Considering the three Tables and this summary, we may safely draw the following conclusions. First, that, as in structure so in function, there are three females or female organs: this is manifest; for when all three receive the very same pollen, they are acted on most differently. So conversely with the thirty-six stamens, we know that they consist of three separate sets of a dozen each, differing in various respects; and in function the pollen of these three sets when applied to one and the same stigma acts most differently, as a glance at the Tables proves. But we shall presently see that the action of the pollen of the whole dozen longest and of the whole dozen shortest stamens is not identical.
Secondly, we see that only the longest stamens fully fertilize the longest pistil, the middle stamens the middle pistil, and the shortest stamens the shortest pistil. And now we can comprehend the meaning of the almost exact correspondence in length between the pistil of each form and the two half-dozen sets of stamens borne by the two other forms; for the stigma of each form is thus rubbed against the same spot of the insect's body, which becomes most charged with the proper pollen. In all three forms, the female organ is but feebly, or not at all, acted on by its own two kinds of pollen. In my papers on the dimorphism of Primula and Linum, I used the terms "heteromorphic" for the fully fertile unions between the female element of the one form and the male element of the other, and "homomorphic" for the less fertile or quite sterile unions between the female and male elements of the same form. The principle involved in these terms holds good with Lythrum, but is insufficient; for though in each of the three forms the fertile unions are all heteromorphic, the ap-
[page] 186
propriate pollen coming from the stamens of corresponding length borne by the other two forms, and though the homomorphic unions of the females with their own two sets of males are always more or less sterile, there remain in each case two other sterile unions, not included in these two terms. Hence it will be found convenient to designate the two unions of each female with the two sets of stamens of corresponding length, which are fully fertile, as legitimate unions, and the four other, more or less sterile, unions of each female with the four other sets of stamens as illegitimate unions. Consequently, of the eighteen possible unions between the three forms, six are legitimate and twelve are illegitimate.
Another and curious conclusion cannot be considered as proved, but is rendered highly probable, by the Tables. The unions of the pistils and stamens of equal length are alone fully fertile. Now with the several illegitimate unions it will be found that the greater the inequality in length between the pistil and stamens, the greater the sterility of the result. There is no exception to this rule. Thus, with the long-styled form, its own shorter stamens are far less equal in length to the pistil than its own longer stamens; and the capsules fertilized by the pollen of the shorter stamens yielded fewer seeds: the same comparative result follows from the use of the pollen of the shorter stamens of the mid-styled form, which are much shorter than the shorter stamens of the short-styled (see diagram), and therefore less equal in length to the long-styled pistil. We shall see exactly the same result if we look to the four illegitimate unions under the mid- and short-styled forms. Certainly the difference in sterility in these several cases is very slight, but the sterility always increases with the increasing inequality of length between the pistil and the stamens which are used. Therefore I believe in the above rule; but a vast number of artificial unions would be requisite to prove it. If the rule be true, we must look at it as an incidental and useless result of the gradational changes through which this species has passed in arriving at its present condition. On the other hand, the correspondence in length between the pistil of each form and those stamens which alone give full fertility is clearly of service to the species, and is probably the result of direct adaptation.
Some of the illegitimate unions yielded, as may be seen in the Tables, during neither year a single seed; but, judging from the case of the long-styled plant, it is probable, if such unions could be effected repeatedly under the most favourable conditions, some few seeds would be produced. Anyhow, I can state that in all
[page] 187
the eighteen possible unions the pollen-tubes penetrated, after eighteen hours, the stigma. I have reason to believe that the offspring from the illegitimate unions present some singular characteristics; but until my observations on this head are repeated, I must be silent. At first I thought that perhaps two kinds of pollen placed together on the same stigma would give more fertility than any one kind; but we have seen that this is not the case with each form's own two kinds of pollen; nor is it probable in any case, as I occasionally got, by the use of single kinds of pollen, fully as many seed as I have seen in a capsule naturally fertilized. Moreover the proper pollen from a single anther is more than sufficient to fully fertilize each stigma; hence, in this as in so many other cases, at least twelve times as much of each kind of pollen is produced as is necessary to ensure full fertilization. From the dusted condition of the whole body of those bees which I caught on these flowers, it is probable that some pollen of all kinds is deposited on each stigma; but there can hardly be a doubt that the pollen of the stamens of corresponding length will be prepotent and will wholly obliterate any effect from the other kinds of pollen, even if previously deposited on the stigma. I infer this partly from the fact ascertained by Gärtner that each species' own pollen is so prepotent over that of any other species, that if put on the stigma many hours subsequently, it will entirely obliterate the action of the foreign pollen. But I draw the above inference especially from the following experiment: I fertilized homomorphically or illegitimately some long-styled Cowslip flowers (Primula veris) with their own pollen, and exactly twenty-four hours subsequently I fertilized these same stigmas heteromorphically or legitimately with pollen from a short-styled dark-red Polyanthus. I must premise that I have raised many seedlings from crossed Cowslips and Polyanthus, and know their peculiar appearance; and I further know, by the test of the fertility of the mongrels inter se, and with both parent forms, that the Polyanthus is a variety of the Cowslip, and not of the Primrose (P. vulgaris) as some authors have supposed. Now from the long-styled Cowslip twice fertilized in the manner explained, I raised twenty-nine seedlings, and every one of them had flowers coloured more or less red; so that the heteromorphic Polyanthus-pollen wholly obliterated the influence of the homomorphic pure Cowslip-pollen, which had been placed on the stigmas twenty-four hours previously, and not a single pure Cowslip was produced.
[page] 188
The last conclusion which may be deduced from the Tables, even from a glance at them, is that the mid-styled form differs from both the others in its much higher capacity for fertilization. Not only did the twenty-four flowers fertilized by the stamens of corresponding lengths, all, or all but one, yield capsules rich in seed; but of the other four illegitimate unions, that by the longer stamens of the short-styled form was highly fertile, though less than in the two legitimate unions, and that by the short stamens of the long-styled form was fertile to a considerable degree; the two unions with this form's own pollen were sterile, but in different degrees. So that the mid-styled form, when fertilized by the six kinds of pollen, evinces five different grades of fertility. By comparing compartments 3 and 6 in Table II. we learn a remarkable fact, namely, that though the pollen from the short stamens of the long-styled and from this form's own (mid-styled) short stamens, used in these two unions, is identical in all respects, yet that its action is widely different; in the one case above half the fertilized flowers yielded capsules containing a fair number of seed; in the other case not one single capsule was produced. So, again, the green, large-grained pollen from the long stamens of the short-styled and from this form's own (mid-styled) long stamens is identical in all respects, but its action, as may be seen in compartments 4 and 5, is widely different. In both these cases the difference in action is so plain that it cannot be mistaken, but it can be corroborated. If we look to Table III., to the legitimate action of the short stamens of the long- and mid-styled forms on the pistil of the short-styled form, we again see a similar but slighter difference, the pollen of the short stamens of the mid-styled form yielding a smaller average of seed during the two years of 1862 and 1863 than that from the short stamens of the long-styled form. Again, if we look to Table I., to the legitimate action of the green pollen of the two sets of long stamens, we shall find exactly the same result, viz. that the pollen of the long stamens of the mid-styled form yielded during both years fewer seeds than that from the long stamens from the short-styled form. Hence it is certain that the two kinds of pollen produced by the mid-styled form are less potent than the similar pollens produced by the corresponding stamens of the two other forms.
When we see that the capsules of the mid-styled form yield a considerably larger average number of seed than those of the other two forms,—when we see how surely the flowers are fertilized in the legitimate unions, and how much more productive
[page] 189
the illegitimate unions are than those of the other two forms, we are led to consider the mid-styled form as eminently feminine in its nature. And although it is impossible to consider as rudimentary or aborted the two perfectly developed sets of stamens of the mid-styled form which produce an abundance of perfectly well-developed pollen, yet we can hardly avoid connecting, as balanced, the higher efficiency of the female organ with the lesser potency of the two mid-styled pollens.
Finally, it is proved by the Tables that Lythrum salicaria habitually produces or consists of three females different in structure and widely different in function; that it produces or consists of three sets of males widely different in structure and function; and that two of the three sets of males are subdivided into subgroups of half a dozen each, differing in a marked manner in potency, so that regularly five kinds of pollen are elaborated by this one species of Lythrum.
Lythrum Græfferi.—I must now say a few words about some of the other species of the genus. I have examined numerous dried flowers of L. Græfferi, each from a separate plant, kindly sent me from Kew. This species, like L. salicaria, is trimorphic, and the three forms apparently occur in about equal numbers. In the long-styled form the pistil projects about one-third of the length of the calyx beyond its mouth, and is therefore shorter than in L. salicaria; the globose and hirsute stigma is larger than that of the other two forms; the longer stamens, which are graduated in length, have their anthers standing just above and just beneath the mouth of the calyx; the half-dozen shorter stamens rise rather above the middle of the calyx. In the mid-styled form the stigma projects just above the mouth of the calyx, and stands almost on a level with the longer stamens of the previous form; its own longer stamens project well above the mouth of the calyx and stand a little above the level of the stigma of the long-styled form; the shorter stamens correspond in all respects with the shorter ones in the previous form. In the short-styled form the stigma of the pistil is nearly on a level with the anthers of the shorter stamens in the two preceding forms; and the longer stamens correspond with the longer stamens of the mid-styled form, and the shorter stamens with the longer stamens of the long-styled form. In short, there is a close general correspondence in structure between this species and L. salicaria, but with some differences in the proportional lengths of the parts. Nevertheless the fact of each of the three pistils having two sets
[page] 190
of stamens, borne by the two other forms, of corresponding lengths, comes out conspicuously. In the mid-styled form the distended pollen-grains from the longer stamens had nearly double the diameter of those from the shorter stamens; so that there is a greater difference in this respect than in L. salicaria. In the long-styled form, also, the difference in diameter between the pollen-grains of the longer and shorter stamens was plainer than in L. salicaria. These comparisons, however, must be received with caution, as they were made on specimens long kept in a dried condition.
Lythrum thymifolia.—This form, according to Vaucher*, is dimorphic like Primula, and therefore presents only two forms. I received two dried flowers from Kew, which presented two forms: in the one form the stigma projected far beyond the calyx, in the other it was included within the calyx; in this latter form the style was only one-fourth of the length of the style of the other form. There are only six stamens; these are somewhat graduated in length, and in the short-styled form the anthers stand a little above the stigma, but yet the stamens by no means equal in length the pistil of the long-styled form; in the long-styled form the stamens are rather shorter than in the other form. These six stamens alternate with the petals, and correspond homologically with the longer stamens of L. salicaria and L. Græfferi. As there are only six stamens, it is scarcely possible that this species can be trimorphic.
Lythrum hyssopifolia.—This species is said by Vaucher, but I believe erroneously, to be dimorphic. I have examined dried flowers from twenty-two separate plants from various localities, kindly sent to me by Mr. Hewett C. Watson,1 Prof. Babington, and others. These were all essentially alike. Hence the species cannot be dimorphic. The pistil varies somewhat in length, but when unusually long the stamens are likewise generally long; in the bud the stamens are short: perhaps these circumstances deceived Vaucher. There are from six to nine stamens, graduated in length; the stamens which are variable in being present or absent correspond with the six shorter stamens of L. salicaria and with the six which are absent in L. thymifolia. The stigma is included within the calyx, and stands in the midst of the anthers, and would generally be fertilized by them; but as the stigma and anthers are upturned, and as, according to Vaucher, there is a passage left in the upper side of the flower to the nectary, there can hardly be a doubt that
* Hist. Phys. des Plantes d'Europe, tom. ii. (1841) pp. 369, 371.
1 Hewett Cottrell Watson (1804-1881), botanist and phrenologist who specialized in the distribution of British plants.
[page] 191
the flowers are visited by insects, which would occasionally bring pollen from other flowers of the same or of any adjoining plant, as surely as occurs with the short-styled L. salicaria, of which the pistil and corresponding stamens closely resemble those of L. hyssopifolia. According to Vaucher and Lecoq*, this species, which is an annual, generally grows almost solitarily, whereas the three preceding species are social; and this alone would almost have convinced me that L. hyssopifolia cannot be dimorphic, as such plants cannot habitually live by themselves any better than one sex of a diœcious species.
Nesæa verticillata.—I raised a number of plants from seed sent me by Professor Asa Gray, and they presented three forms. These differed from each other in the proportional lengths of their organs of fructification and in all respects in very nearly the same way as the three forms of Lythrum Græfferi. The green pollen-grains from the longest stamens, measured along their greater axis and not distended with water, were 13/7000 of an inch in length; those from the stamens of middle length 9-10/7000, and those from the shortest stamens 8-9/7000 of an inch.
We have seen that the genus Lythrum affords trimorphic, dimorphic, and monomorphic species.
The inquiry naturally arises, why do these species differ so remarkably in their sexual relations? of what service can reciprocal dimorphism or trimorphism be to certain species, whilst other species of the same genus present, like the great majority of plants, only one form? I have elsewhere given too briefly† the
* Géograph. Bot. de l'Europe, tom. vi. (1857) p. 157.
† 'Origin of Species,' 3rd edit., p. 101. Hugo von Mohl1 has recently (Bot. Zeitung, 1863, S. 309, 321), in a most interesting paper, advanced the case of the minute, imperfectly developed, closed and self-fertile flowers borne by Viola, Oxalis, Impatiens, Campanula, &c., as an argument against my doctrine that no species is self-fertilized for perpetuity. I may state that in the spring of 1862 I examined some of these flowers, and saw, though less thoroughly, all that H. von Mohl has so well described. I can add only one remark, which I believe is correct, that in V. canina there is an open channel for the pollen-tubes from the extremity of the stigma to the ovarium; for I gently pressed a minute bubble of air repeatedly backwards and forwards from end to end. Though the imperfectly developed and the perfect flowers are so different in structure, it is a rather curious case of correlation, that in the double purple Violet (V. odorata) the minute imperfect flowers are double to the very core, so that a section appears like the head of a cabbage when cut through. There can be, as von Mohl asserts, no doubt that these flowers are always self-fertilized; they are moreover specially adapted for this end, as may be seen in the remarkable difference in the shape of the pistil in V. canina (and in a less degree in V. hirta and the single V. odorata) as compared with that of the perfect flower;
1 Hugo von Mohl (1805-1872), German botanist and physiologist who co-founded the Botanische Zeitung, 1843. Mohl 1863.
[page] 192
general grounds of my belief that with all organic beings distinct individuals at least occasionally cross together, and reciprocal dimorphism is plainly one most efficient means for ensuring this result.
____________________________________________________________
and in the pollen-tubes which proceed from the grains within the anthers in V. canina, and from within the lower anthers of Oxalis acetosella, having the wonderful power of directing their course to the stigma. If these plants had produced the minute closed flowers alone, the proof would have been perfect that they could never have crossed with other individuals. I am aware that in some of these cases it has been stated that the perfect flowers never produce any seed; as far as Amphicarpæa is concerned, I hear from Professor Asa Gray that the Petaliferous flowers certainly sometimes yield seed. The completely enclosed flowers of that curious grass, the Leersia oryzoides, as described by M. Duval-Jouve1 (Bull. Soc. Bot. de France, tom. x. 1863, p. 194), apparently offer the best case of perpetual self-fertilization; for when perfect flowers are protruded from the culms, they are, as far as is yet known, always sterile. In a number of plants kept by me in pots in water, not one single perfect flower has protruded, but the enclosed flowers produced plenty of seed. Without wishing to throw any doubt on M. Duval-Jouve's excellent observations, I may add that with the enclosed flowers borne by my plants, the act of fertilization, that is, the penetration of the stigma by the pollen-tubes, took place in the air and not in fluid within the glumes. With the exception of the Leersia, as the case now stands, I cannot see how the production of the small, imperfect flowers invalidates my doctrine that no species is perpetually self-fertilized, more than the multiplication of many plants by bulbs, stolons, &c. As I observe that the production of seed by the perfect flowers of Viola is spoken of as something capricious and accidental, I may state that, although it varies much in different years, it depends exclusively on the visits of bees; I ascertained this by marking many flowers thus visited, and finding that they produced capsules, and by covering up many flowers which (excepting a few that I artificially fertilized) did not, when thus protected, produce a single capsule. After bees have visited these flowers, the pollen may be seen scattered on the papillæ and on the stigma itself, and they can hardly fail thus to cross distinct individuals. These remarks apply to V. canina, hirta, and odorata; with V. tricolor the case is somewhat different; but I must not enlarge any more on this subject. The production by so many plants of perfect and expanded, as well as of imperfect and closed flowers, seems to me to throw much light on many points; it shows how extraordinarily little pollen is necessary for full fertilization, for I ascertained with V. canina that the perfect and imperfect flowers (the latter producing so few pollen-grains) yielded the same average number of seeds; it shows us that fertilization can be perfected in closed flowers; it shows us that large, highly coloured petals, perfume, and the secretion of nectar are by no means indispensable for this act, even in those species which properly possess these characters. It seems to me that the necessity of an occasional cross with a distinct individual of the same species explains the universal presence of at least some expanded flowers, at the expense of injury from rain and the loss of much pollen by innumerable pollen-robbing insects; it explains the enormous superfluity of pollen from its liability to loss from these causes and during conveyance from flower to flower; it explains the use of a gaily coloured corolla, perfume, and nectar, namely, to attract insects, except in those comparatively few cases in which wind is the agent, and in these the last-named attributes are deficient.
1 Joseph Duval-Jouve (1810-1883), French botanist.
[page] 193
This result would appear to be one of high importance, for with dimorphic plants it is ensured at the risk of occasional sterility; not only is the pollen of each plant useless or nearly useless to that individual, but so is the pollen of all the plants of the same form, that is, of half the total number of individual plants. In that extensive class of plants called by C. K. Sprengel dichogams, in which the pollen of each flower is shed before its own stigma is ready, or in which the stigma (though this case occurs more rarely) is mature before the flower's own pollen is ready sterility can hardly fail to be the occasional result; and it would be the inevitable result with both dichogamous and reciprocally dimorphic flowers unless pollen were carried by insects (and in some few species by the wind) from one flower or plant to the other. As with reciprocal dimorphism so with dichogamy, within the same genus some of the species are and some are not thus characterized. Again, in the same genus, as in that of Trifolium, some species absolutely require insect-aid to produce seed, others are fertile without any such aid; now when insects are requisite for fertilization, pollen will generally be carried from one flower to the other. We thus see, by means of reciprocal dimorphism, of dichogamy, and of insect-aid, that some species require, or at least receive, incessant crosses with other individuals of the same species; whereas other species of the same genera can be, and probably are often fertilized during long periods by the pollen of their own flowers. Why this wide difference in the frequency of crosses should occur we are profoundly ignorant. I will only further remark on this head, that it would be a great mistake to suppose that many flowers, which are neither reciprocally dimorphic nor dichogamous, nor require insect-aid for their fertilization, nor show any particular adaptation in their structure for the visits of insects, are not habitually crossed with the pollen of other individuals; this occurs, for instance, habitually with cabbages, radishes, and onions, which nevertheless are perfectly fertile (as I know by trial) with their own pollen without aid of any kind.
But it may be further asked, granting that reciprocal dimorphism is of service by ensuring at each generation a cross (but I am far from pretending that it may not have some additional unknown signification), why did not dimorphism suffice for L. salicaria and Græfferi? why were they rendered reciprocally trimorphic, entailing such complicated sexual relations? We cannot answer, except perhaps so far:—if we suppose two plants of
[page] 194
the L. salicaria to grow by themselves, then if the species were dimorphic it would only be an equal chance in favour of the two turning out different forms and consequently both being fertile; but as the species is trimorphic and each form can fertilize the two other forms, it is two to one in favour of the two turning out different forms and being consequently both fertile. We thus see how reciprocal trimorphism must be an advantage; and probably it would be more advantageous to this Lythrum, which commonly grows in almost a single row along the banks of streams, than it would be to Primroses or Cowslips which have neighbours on all sides. But even if trimorphism effected no good beyond that gained by dimorphism, we ought not to feel much surprised at its occurrence, for we continually see throughout nature the same end gained by the most complicated as well as by the most simple means: to give one instance:—in many diœcious plants pollen is carried from the male to the female by the wind, which is perhaps the simplest method conceivable, or by the adherence of the grains to the hairy bodies of insects, which is a method only a little less simple; but in Catasetum the conveyance is effected by the most complex machinery; for in this orchid we have sensitive horns which when touched cause a membrane to rupture, and this sets free certain springs by which the pollen-masses are shot forth like an arrow, and they adhere to the insect's body by a peculiar viscid matter, and then by the breaking of an elastic thread of the right strength the pollen is left sticking to the stigma of the female plant. The complexity of the means used in this and in many other cases, in fact depends on all the previous stages through which the species has passed, and on the successive adaptations of each part during each stage to changed conditions of life.
As some authors consider reciprocal dimorphism to be the first step toward diœciousness, the difficulty of understanding how a trimorphic plant like Lythrum salicaria could become diœcious should be noticed; and as dimorphism and trimorphism are so closely allied, it is not probable that either state is necessarily in any way related to a separation of the sexes—though it may occasionally lead to this end. As far as Lythrum salicaria is concerned, the one tendency which we can discover is towards the abortion of the two sets of stamens in the mid-styled form. This tendency is evinced by its pollen, though abundant and apparently good, yielding a smaller percentage of seed than does the pollen of the corresponding stamens in the other two forms; and this
[page] 195
fact is in itself curious, and shows by what insensibly graduated steps nature moves. If this tendency were carried out the mid-styled form would become a female, depending for its fertilization on two sets of stamens in the long- and short-styled forms; and these two forms would reciprocally fertilize each other like the two forms of Primula or Linum; but there would be no approach to a diœcious condition.
As the case of the trimorphic species of Lythrum is so complicated, and as it is easier to perceive the relations of the sexes in the animal than in the vegetable kingdom, it may be worth while to give, before concluding, a somewhat elaborate simile. We may take the case of a species of Ant, and suppose all the individuals invariably to live in three kinds of communities; in the first, a large-sized female (not to specify other differences) living with six middle-sized and six small-sized males; in the second, a middle-sized female with six large- and six small-sized males; and in the third community, a small-sized female with six large- and six middle-sized males. Each one of these three females, though enabled to unite with any male, would be nearly sterile with her own two sets of males, and likewise with two other sets of males living in the other two communities; for she would be fully fertile only when paired with a male of her own size. Hence the thirty-six males, distributed by half-dozens in the three communities, would be divided into three sets of a dozen each; and these sets, as well as the three females, would differ from each other sexually in exactly the same manner as distinct species of the same genus. Moreover the two sets of males living in the community of the extraordinarily fertile middle-sized female would be less potent sexually than the males of corresponding size in the two other communities. Lastly, we should find that from the eggs laid by each of the three females, all three sorts of females and all three sorts of males were habitually reared—proving to demonstration that all belonged to one and the same species.
To appreciate fully this remarkable case of the reciprocally trimorphic species of Lythrum, we may take a glance at the two great kingdoms of nature and search for anything analogous. With animals we have the most astonishing diversity of structure in the so-called cases of alternate generation, but as such animals have not arrived at maturity, they are not properly comparable with the forms of Lythrum. With mature animals we have extreme differences in structure in the two sexes; we have in
[page] 196
some of the lower animals males, females, and hermaphrodites of the same species; we have the somewhat more curious case of certain Cirripedes which are hermaphrodites, but are sexually aided by whole clusters of what I have called complemental males;1 we have, as Mr. Wallace has lately shown, the females of certain Lepidoptera existing under three distinct forms; but in none of these cases is there any reason to suspect that there is more than one female or one male sexual element. With certain insects, as with Ants, in which there exist, besides males and females, two or three castes of workers, we have a slightly nearer approach to our case, for the workers are so far sexually affected as to have been rendered sterile. With plants, at least with phanerogamic plants, we have not that wonderful series of successive developmental forms so common with animals; nor could this be expected, as plants are fixed to one spot from their birth, and must be adapted throughout life to the same conditions. With plants we have sexual differences in structure, but apparently less strongly marked than with animals, from causes which are in part intelligible, such as there being no sexual selection; again, we have that class of dimorphic flowers so ably discussed recently by Hugo von Mohl, in which some of the flowers are minute, imperfectly developed, and necessarily self-fertile, whilst others are perfect and capable of crossing with other flowers of the same species; but in these several cases we have no reason to suspect that there is more than one female or one male sexual element. When we come to the class of reciprocally dimorphic plants, such as Primula, Linum, &c., we first meet with two masculine and two feminine sexes. But these cases, which seemed only a short time since so strange, now sink almost into insignificance before that of the trimorphic species of Lythrum.
Naturalists are so much accustomed to behold great diversities of structure associated with the two sexes, that they feel no surprise at the fact; but differences in sexual nature have been thought to be the very touchstone of specific distinction. We now see that such sexual differences—the greater or less power of fertilizing and being fertilized—may characterize and keep separate the coexisting individuals of the same species, in the same manner as they characterize and have kept separate those groups of individuals, produced from common parents during the lapse of ages or in different regions, which we rank and denominate as distinct species.
1 Living Cirripedia (1851), p. 55.