




Darwin's Study of the Cirripedia1
An introduction by Marsha Richmond
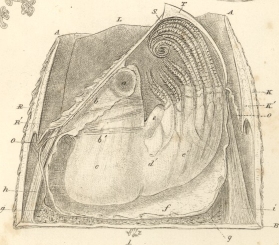 Darwin's work on barnacles (Cirripedia), conducted between 1846 and 1854, has long posed problems for historians. Coming between his transmutation notebooks and the Origin of species, it has frequently been interpreted as a digression from Darwin's species work. Yet when this study is viewed in the context of Darwin's earlier interests, in particular his studies of marine invertebrates carried out during his student days in Edinburgh and later on board the Beagle, the monograph on the Cirripedia seems less anomalous. Moreover, Darwin's study of cirripedes, far from being merely a dry, taxonomic exercise, was a highly theoretical work that addressed several problems at the forefront of contemporary natural history. Treating a group of organisms of considerable interest to mid-nineteenth century naturalists and approaching their classification using the most recent methods available, Darwin was able to provide a thorough taxonomic study that has remained a standard work in cirripede morphology and systematics. For Darwin personally, the barnacle work perfected his understanding of scientific nomenclature, comprising both theoretical principles and technical facility with the methods of comparative anatomy. It also provided him with an empirical means of testing his views on the species question (Crisp 1983).
Darwin's work on barnacles (Cirripedia), conducted between 1846 and 1854, has long posed problems for historians. Coming between his transmutation notebooks and the Origin of species, it has frequently been interpreted as a digression from Darwin's species work. Yet when this study is viewed in the context of Darwin's earlier interests, in particular his studies of marine invertebrates carried out during his student days in Edinburgh and later on board the Beagle, the monograph on the Cirripedia seems less anomalous. Moreover, Darwin's study of cirripedes, far from being merely a dry, taxonomic exercise, was a highly theoretical work that addressed several problems at the forefront of contemporary natural history. Treating a group of organisms of considerable interest to mid-nineteenth century naturalists and approaching their classification using the most recent methods available, Darwin was able to provide a thorough taxonomic study that has remained a standard work in cirripede morphology and systematics. For Darwin personally, the barnacle work perfected his understanding of scientific nomenclature, comprising both theoretical principles and technical facility with the methods of comparative anatomy. It also provided him with an empirical means of testing his views on the species question (Crisp 1983).
Darwin's interest in invertebrate zoology stemmed from his years in Edinburgh and, in particular, his contact with Robert Edmond Grant (Autobiography, pp. 49-50; Hodge 1985; Sloan 1985). This interest in marine organisms was exercised during the Beagle voyage. In 1835, in the Chonos Archipelago off the coast of Chile, Darwin found 'most curious' minute cirripedes buried within the shell of a gastropod mollusc. In his zoological notes, Darwin recorded: 'The thick shell of some of the individuals of the Concholepas Peruviana is completely drilled by the cavities formed by this animal.—' (DAR 31: 305). He gave a detailed description and tentatively identified this burrowing barnacle as a member of the Balanidae, or sessile acorn barnacles, most frequently found attached to rocks. Yet from the absence of a shell and its unusual parasitic nature, Darwin recognised that it differed greatly from common barnacles.
It was perhaps Darwin's further discovery of developing eggs within the base of the barnacle that most captured his interest. He recorded seeing four different stages in the larval development of this 'Balanus' and remarked on the resemblance of one stage to that observed in the metamorphosis of Crustacea (DAR 31: 307). This observation was notable, for in 1835 the presence of larval stages of cirripedes was still a matter of dispute among naturalists.
Prior to the publication in 1830 of John Vaughan Thompson's account of the developmental history of cirripedes, which pointed out the similarity of barnacle larvae to those of Crustacea, most naturalists had followed Linnaeus and Cuvier in classifying the cirripedes as molluscs because of their external shelly covering and because their mantle cavity contained sea-water (Winsor 1969). Thompson's sequential observations of the metamorphosis of nauplius and cypris larvae into adult barnacles, reinforced (and reinterpreted) a few years later by Hermann Burmeister (Burmeister 1834), first revealed the developmental stages of these organisms. The consequent sudden shift of the Cirripedia from one branch of the animal kingdom to another—from the Mollusca to the Articulata—indicated to mid-nineteenth century naturalists that a revaluation of the group, based on a systematic and anatomical comparison intra se and with other Crustacea, was needed.
Such a revaluation had not been undertaken when, in 1846, Darwin began to examine several invertebrates that remained undescribed from his Beagle collections. After writing two papers, one on Sagitta and another on the Planariae, he began to work on his curious South American cirripede, which he initially called Arthrobalanus and later renamed Cryptophialus minutus. 'I had originally intended,' he explained in the preface to Living Cirripedia (1851): vii, 'to have described only a single abnormal Cirripede, from the shores of South America, and was led, for the sake of comparison, to examine the internal parts of as many genera as I could procure.' For fourteen months Darwin pursued an anatomical study of pedunculated and sessile cirripedes, during which time he realised the profound state of disarray in the taxonomy of the group. Late in 1847, John Edward Gray, keeper of the zoological collections at the British Museum and himself a cirripede expert, suggested to Darwin that he prepare a monograph of the entire sub-class. Gray provided him with his own collection, arranged access to the museum's specimens, and advised him on procuring other collections. At the time Darwin could not have foreseen that this project would occupy his attention for the next seven years. To appreciate why Darwin would have undertaken such a study, it is helpful to review the science of systematics and some of the concerns of natural history in the nineteenth century.
Classification was a primary concern of eighteenth and nineteenth-century naturalists (Knight 1981). Many of Darwin's contemporaries undertook taxonomic study of one major group or another. But it was perhaps the invertebrates, selectively treated by Linnaeus and Cuvier, that presented the greatest challenge to systematists. As historians have recently shown, an individual's conception of the order of nature shaped the particular classificatory scheme they developed (Desmond 1982; Richards 1987; Winsor 1969).
Darwin's views on classification were tempered by his attempts to classify specimens during the Beagle voyage, subsequent discussions with experts describing his collections, and his consideration of the subject in the light of his views on the transmutation of species. Darwin's discussions with Owen no doubt helped sharpen his thinking about classifying. Influenced by the philosophical anatomy of Ettienne Geoffroy Saint-Hilaire, Owen was formulating new guidelines for taxonomy, including a precise definition of homology (a term he introduced to replace the vaguer notion of 'affinity'), a particular application of the data of comparative anatomy, and an archetype to represent the common design perceived among organisms. Within Darwin's maturing evolutionary perspective, the principles of natural classification began to assume a new meaning (Ospovat 1981). As Ghiselin has noted, they 'ceased to be merely descriptive and became explanatory.' (Ghiselin 1969, p. 83).
By the early 1840s, Darwin's ideas on classification were well developed (see Correspondence vol. 2, letter to George Robert Waterhouse, [26 July 1843]). Within his theory of classification, homological relations became more than simply tools for description. For Darwin homology revealed actual phylogenetic relationship rather than just similarity in the basic plan of organisation. In Darwin's classification of the cirripedes, homology provided the key to making out the evolutionary relationships linking members of a group and the possible line of descent of one species from another previously existing form.
Darwin's evolutionary interpretation of the meaning of classification explains why he readily adopted embryology as a methodological tool for revealing homologies. Through the work of naturalists such as Robert Brown, Martin Barry, and Owen in England and Henri Milne-Edwards in France, the substance of Karl Ernst von Baer's work on embryological development began to enter discussions of classification in the late 1830s and early 1840s (Ospovat 1976; Richards 1987; Appel 1987). Having been introduced to these ideas through Grant (Desmond 1984; Sloan 1985), Darwin's understanding of the importance of embryology deepened through his reading of Milne-Edwards's influential essay on classification (Milne-Edwards 1844). Like von Baer, Milne-Edwards recognised that comparative embryogenesis could be used to yield information about systematic relationships. Within members of the same branch of the animal kingdom, the progress of development appeared to illustrate an increasing divergence from an early resemblance shared by all members of a class to those later features special to the particular order, family, genus, and species.
Milne-Edwards drew from this generalisation several principles that proved important for classification in the mid-nineteenth century: (1) the most general structures of a class appear earliest in development, and these establish higher taxonomic affinities; (2) characters shared by organisms reflect the degree of zoological parentage; (3) some organisms, in contrast to the general phenomenon of 'progressive' development, exhibit arrested or retrograde development; (4) increasing specialisation in embryogenesis illustrates the tendency in higher organisms toward a 'division of physiological labour' and this principle could be used to determine 'lowness' and 'highness' in particular groups; and (5) embryology, by revealing homological relationships in development, was the best means for classification.2
Darwin was particularly struck by Milne-Edwards's formulation; he prefaced his lengthy abstract of this essay with the statement, '—This is the most profound paper I have ever seen on Affinities' (DAR 72: 117). Many of the points discussed by Milne-Edwards re-emerged in his monographs within his treatment of the natural history and systematics of the cirripedes. Darwin was also influenced by another important paper by Gaspard Auguste Brullé (Brullé 1844), who argued that the most complex and characteristic organs in a group were the first to develop, thereby adding the dimension of time and functional importance (DAR 72: 123 and v.; Rachootin 1984). Darwin's understanding of embryological development, as outlined in his essay of 1844 (Foundations, pp. 57-255), accorded well with these views.3 In Living Cirripedia, Darwin appears to have used Brullé's law to establish some of the embryological homologies.4
Darwin clearly believed that a classification based on homologies established through embryology as well as anatomy would best reveal genetic relationships. Accordingly, homology became a key element in taxonomic evaluation of the cirripedes. It justified, for example, his decision to rank the Cirripedia as a separate sub-class of the Crustacea rather than subsuming the group within another sub-class (Milne-Edwards 1852) or elevating it to a separate class altogether (Owen 1855). Darwin, however, with his particular embryological criterion of homology, believed that the resemblances in the metamorphosis of Crustacea and the Cirripedia indicated their community of descent. Since cirripedes exhibited characters common to two different crustacean sub-classes, he decided to rank the Cirripedia as a separate sub-class of Crustacea.5
An understanding of the theoretical principles upon which Darwin drew thus provides the context for assessing the monograph on the Cirripedia. Yet it is Darwin's particular application of these classificatory concepts that is the key to interpreting the barnacle study, and this can perhaps best be illustrated by citing a few of the leading problems treated in this work (see also Autobiography, pp. 117- 18).
In both volumes of Living Cirripedia (1851 and 1854), Darwin devoted an introductory section to a description of the metamorphosis of cirripedes, explaining that this was necessary 'on account of the great importance of arriving at a correct homological interpretation of the different parts of the mature animal.' (Living Cirripedia (1851): 25). As a basis for his homologies, Darwin relied upon Milne-Edwards's model of an archetypal crustacean consisting of twenty-one segments, variously divided in different organisms between cephalic, thoracic, and abdominal somites (Milne-Edwards 1834-40; Appel 1987, pp. 218-19). Darwin identified seventeen of these twenty-one segments in the archetypal cirripede and assumed that the four terminal crustacean segments were missing in barnacles.6 This archetypal cirripede was essential to Darwin's treatment of the group; it formed the basis for making out the anatomical organisation of specimens and guided his assessment of taxonomic rank. Furthermore, studying the developmental history of cirripedes and thereby analysing the homologies of the adult, Darwin found some of the basic tenets of his species theory well demonstrated: the homology of parts in related organisms, the loss of useless organs (e.g., the abdominal segments and the swimmerets), and the transformation in function of homologous organs (e.g., thoracic limbs for walking into cirri feeding).
The use of homology as a means to assess relationship was particularly central to Darwin's discussions of difficult or anomalous forms. For delineating the higher taxa, Darwin was guided by a comparison of the metamorphosis and the segmentation of particular forms with the segments of the archetypal cirripede. One example of the importance of embryological homologies comes from his description of the organs that served to transform a previously mobile crustacean into a state of permanent attachment. Such a system was of particular interest to him from a theoretical standpoint. In cirripede larvae in the last stage of development, Darwin observed 'two long, rather thick, gut-formed masses, into the anterior ends of which the cement-ducts running from the prehensile antennæ could be traced', and he came to believe that these were the incipient ovaria and the cement glands of the organism (Living Cirripedia (1851): 20). This association suggested to him that the cementing apparatus was homologically equivalent to the ovarian tube. The case appeared to be a striking instance of how an organ had been transformed to perform a new function (Living Cirripedia (1851): 37-8; Living Cirripedia (1854): 151-2).
Crisp (1983) has pointed to Darwin's interpretation of the cement glands as an example in which evolutionary views influenced, in this case erroneously, his understanding of the phenomena. Certainly Darwin's view did accord well with Milne-Edwards's principle of the division of physiological labour as a means for the development of structural specialisation and with Darwin's understanding of how pre-existing organs could become modified to fulfill a new function in an organism.7 It explains why Darwin found it difficult to abandon this homology when it was challenged in 1859 by August Krohn. As he admitted in a letter to Charles Lyell, 28 September 1860 (Correspondence vol. 8), 'It is chiefly the interpretation which I put on parts that is so wrong; & not the part which I describe.' But this 'blunder' (as Darwin called it in his Autobiography), was more than a matter of interpretation. It challenged his best case for descent with modification and ultimately his picture of how the archetypal cirripede had evolved from the ancestral crustacean.
Perhaps the clearest example of how Darwin's transformist views may have influenced his taxonomic decisions comes from his discussion of the sexual relations of cirripedes. The hermaphroditism of cirripedes is one of the major characters distinguishing the majority of the group from other crustaceans. Soon after commencing the monograph, however, Darwin discovered in the genus Ibla a species in which small, rudimentary males were found parasitic on the female. Subsequently Darwin encountered a further sexual peculiarity which he regarded as even more significant. In both Ibla and Scalpellum, he also found minute or, as he called them, 'complemental' males attached, not to a female, but to a hermaphrodite. Apart from the 'marvelous' fact that these complemental males were so 'utterly different in appearance and structure' from the hermaphrodite—the two representing such 'diverse beings, with scarcely anything in common, and yet all belonging to the same species!' (Living Cirripedia (1851): 293)—this discovery was unique in the animal kingdom, and it touched upon, Darwin realised, the question of sexuality and its evolution.8
Darwin viewed his discovery of males and complemental males in these two genera as a significant confirmation of his transformist views. Here seemed to be a prominent illustration of a gradual change in nature within a few members of a group from one condition to another—in this case, from hermaphroditism to separate sexes (Living Cirripedia (1854): 29).9 Darwin's elation over this discovery was frequently expressed in his correspondence. Immediately after finding little males attached to hermaphrodite cirripedes, for example, on 10 May 1848 Darwin informed Hooker how this discovery related to his species theory, writing: 'these parasites, I now can show, are supplemental males, the male organs in the hermaphrodite being unusually small, though perfect & containing zoosperms: so we have almost a polygamous animal, simple females alone being wanting. I never shd. have made this out, had not my species theory convinced me, that an hermaphrodite species must pass into a bisexual species by insensibly small stages, & here we have it, for the male organs in the hermaphrodite are beginning to fail, & independent males ready formed.' (Correspondence vol. 4). It was certainly his species theory that prompted him to look for similar relations in closely allied genera.
Another notable aspect of Darwin's study of the Cirripedia was the amount of variation shown by members of the group. Interesting though this was from a theoretical point of view, it caused considerable difficulty in deciding whether such variations indicated different species or merely varieties (Southward, 1983). In Living Cirripedia (1854), Darwin clearly stated the consequences variation had for taxonomic evaluation, concluding with a generalisation that appeared again in chapter five of Origin (see Living Cirripedia (1854): 155). One of the first systematists to break from the type-concept of species, Darwin realised that within a given species variation must necessarily occur. But he had difficulty deciding, as do modern systematists, where species variations stopped and distinct species began. It was, perhaps, because Darwin had an overall concept of evolutionary change that he 'confounded some taxa and unjustifiably separated others' (Henry and McLaughlin 1975, p. 8).
Darwin specifically addressed how his views on species influenced taxonomic decisions in his correspondence with Hooker (see Correspondence vol. 4, letter to J. D. Hooker, 13 June [1850], and Correspondence vol. 5, letter to Hooker on 25 September [1853]). It is clear that Darwin's views on the impermanence of species led him to lump what we today recognise as many separate species together as varieties of one species.
Thus, long before Darwin commenced his study of barnacles, he was intrigued by the processes of generation and embryonic development of marine invertebrates. Moreover, he was well informed about the current views of leading systematists concerning the theory of classification. By the time he took up the study of barnacles, he was familiar with the embryological criterion of homology offered by respected naturalists, studying their works with a high degree of interest and attention and drawing out of their writings statements that directly bore on the Cirripedia. From these sources Darwin developed a methodology that could yield a classification based on phylogenetic relationships. He combined theory with considerable practical skill in anatomical dissection, an important prerequisite for the preparation of a taxonomic monograph. His notable accomplishments were recognized in 1853, even before completing the second parts of Living Cirripedia and Fossil Cirripedia, when he was awarded the Royal Medal of the Royal Society of London (see Proceedings of the Royal Society 6 (1853): 355-6).
Marsha Richmond
January 2007
See also Dates of publication of two of Darwin's volumes on the Cirripedia (barnacles) by W. A. Newman.
1851 [=1852]. Living Cirripedia, A monograph on the sub-class Cirripedia, with figures of all the species. The Lepadidæ; or, pedunculated cirripedes. Vol. 1 Text Image PDF F339.1
1854. Living Cirripedia, The Balanidæ, (or sessile cirripedes); the Verrucidæ. Vol. 2 Text Image PDF F339.2
1851. Fossil Cirripedia of Great Britain: A monograph on the fossil Lepadidae, or pedunculated cirripedes of Great Britain. Vol. 1 Text Image PDF F342.1
1854 [=1855]. A monograph on the fossil Balanidæ and Verrucidæ of Great Britain. Vol. 2 Text Image PDF F342.2
1 This essay is an abridged version of Appendix II, published in Frederick Burkhardt, Sydney Smith, Janet Browne, Stephen Pocock, Marsha L. Richmond, and Anne Secord eds. 1988. The Correspondence of Charles Darwin. Cambridge: Cambridge University Press, pp. 388-409. Written by Marsha Richmond, the essay was expertly revised by Janet Browne, Stephen Pocock and Anne Secord. I thank the Editors of the Darwin Correspondence and Cambridge University Press for granting permission to reproduce the essay in an abridged form.
2 In his essay of 1842, Darwin wrote: “The natural system being on theory genealogical, we can at once see, why fœtus, retaining traces of the ancestral form, is of the highest value in classification.” (Foundations, p. 45). See the fuller discussion of this topic in the 1844 essay (Foundations, p. 229).
3 In his species essay of 1844, for example, Darwin stated: “The cause of the greater value of characters, drawn from the early stages of life, can . . . be in a considerable degree explained, on the theory of descent, although inexplicable on the views of the creationist.” (Foundations, p. 201).
4 Darwin later, somewhat reluctantly, abandoned Brullé's law, having been assured by Thomas Henry Huxley that it was empirically invalid (Correspondence vol. 6, letter to T. H. Huxley, 5 July [1857], and letter from T. H. Huxley, 7 July 1857).
5 Darwin had arrived at such a view of cirripede systematics by 1844, judging by statements in the essay on species drafted in that year. He expressed his belief that a “natural classification follows according to the degrees of resemblance in the parts of most physiological importance”, illustrating the point by reference to the cirripedes: “The affinity of the common rock-barnacle with the Crustaceans hardly be perceived in more than a single character in its mature state, but whilst young, locomotive, and furnished with eyes, its affinity cannot be mistaken.” (Foundations, p. 201).
6 Darwin discussed his conception of archetype in a letter to Huxley, 23 April [1853] (Correspondence vol. 5): “The discovery of the type or 'idea' (in your sense, for I detest the word as used by Owen, Agassiz & Co) of each great class, I cannot doubt is one of the very highest ends of natural history: & certainly most interesting to the worker out ... I shd. have thought that the archetype in imagination was always in some degree embryonic, & therefore capable & generally undergoing further development.—'
7 See Foundations, pp. 40-1, 238.
8 This phenomenon had long been recognised by botanists; Darwin, in fact, referred to the analogy with plants in Living Cirripedia (1851): 214: “Although the existence of Hermaphrodites and Males within the limits of the same species, is a new fact amongst animals, it is far from rare in the Vegetable Kingdom: the male flowers, moreover, are sometimes in a rudimentary condition compared to the hermaphrodite flowers, exactly in the same manner as are the male Iblas.” Darwin's earlier interest in the question of hermaphroditism can be seen in his Notebook D (Notebooks, 1987), where he developed the theory of the origin of separate sexes from an ancestral hermaphroditic organism.
9 Stauffer, Natural selection, pp. 362-3. Darwin often used 'bisexual' in two different senses, sometimes as a synonym for “hermaphrodite” and at other times to denote the separation of sexes.
RN5